

 PDF
PDF Citation
Citation Print
Print


Introduction
Human pluripotent stem cell (hPSC)-derived intestinal organoids (HIOs) recapitulate developmental processes, and can differentiate into the multiple cell types found within the human intestine in vivo (1, 2). HIOs are cultured three-dimensionally inside extracellular matrix (ECM) domes such as a Matrigel dome, and showing the morphology of a miniaturized gut with a crypt-villus structure. Owing to the high structural and functional similarities between the in vivo human intestine and in vitro HIOs, HIOs can be used in various application studies such as disease modeling, drug screening, and development of regenerative therapies (3, 4). Although HIOs have high potential and applicability, there is a growing need for intestinal stem cell (ISC) model systems to study ISC biology. Most early findings related to human ISCs have involved expensive, labor-intensive, and time-consuming mouse models and colorectal cancer cell lines (5). Therefore, it is necessary to develop a robust and reliable culture method capable of exponential proliferation and long-term culture of ISCs.
To address these issues, we recently developed a culture method for 3D expanding intestinal spheroids (InSexp) derived from 3D HIOs differentiated from hPSCs. InSexp comprise mainly intestinal stem cells (ISCs) and progeni-tors. In this study, we investigated the exponential proliferation and long-term culture of ISCs by inducing rapid expansion of InSexp through WNT pathway activation.
Materials and Methods
Human pluripotent stem cells
The H9 human embryonic stem cells (hESCs) were purchased from WiCell Research Institute (Madison, WI, USA). Human induced pluripotent stem cells (hiPSCs) were generated as described previously (6).
Differentiation of hPSCs into immature and mature 3D HIOs
hPSC-derived 3D HIOs were generated as reported previously (1, 2). The 3D HIOs were mechanically dissociated and re-embedded every 10∼14 days. For in vitro maturation of HIOs, 1 ng/ml of IL-2 (R&D systems) was added to the culture medium when the spheroids were embedded. IL-2 was freshly added after at least two passages.
Isolation and culture of 3D InSexp derived from HIOs
The 3D InSexp derived from HIOs were generated as previously reported (7). The growth medium was replaced every other day. To optimize the organoid growth, 1 μM of CHIR99021 (Tocris) was included in the InSexp growth medium.
Passaging of 3D InSexp
For passaging, InSexp were dissociated using trypsin-EDTA and a P1000 pipette was used to break up the Matrigel dome. The total mixture was transferred to a conical tube, incubated at 37℃ in a water bath for 5 min, and mechanically dissociated using 10 rounds of pipetting. The dissociated cells were resuspended and centrifuged at 1,500 rpm, 4℃, for 5 min. After resuspending the cell pellet with Matrigel, the HIO-Matrigel mixture was plated onto a four-well culture plate and incubated at 37℃ in a CO2 incubator for more than 30 min to solidify the Matrigel, followed by the addition of InSexp growth medium supplemented with 10 μl of Y-27632 (Tocris), and 1 μM Jagged-1 (AnaSpec, Fremont, CA, USA) for the first 2 days. Subsequently, this medium was replaced with the InSexp culture medium, which was then replaced every other day.
Quantitative reverse transcription-polymerase chain reaction
Total RNA was extracted from cells using the RNeasy kit (Qiagen), and cDNA was prepared using a Superscript IV cDNA synthesis kit (Thermo Fisher Scientific Inc.). Quantitative polymerase chain reaction (qPCR) was performed using a 7500 Fast Real-Time PCR system (Applied Biosystems, Foster city, CA, USA). Relative expression was calculated using the ΔΔCt method. The primers used in this study are listed in Table S1.
Immunofluorescence analysis
Immunofluorescence staining was performed on 10% formalin-fixed InSexp as described previously (8). Cryosections were permeated with 0.1% Triton X-100, and then blocked for 1 hour with 4% bovine serum albumin. After blocking, the sections were incubated with primary antibodies at 4℃ overnight and the secondary antibody at room temperature. Finally, 4’,6-diamidino-2-phenylindole (DAPI; 1 mg/ml, Thermo Fisher Scientific Inc.) was used to counterstain the nuclei. Fluorescence images were acquired using a fluorescence microscope (IX51, Olympus, Tokyo, Japan) and LSM800 conforcal microscope (Carl Zeiss, Gottingen, Germany). The list of primary antibodies used is provided in Table S2.
Cell counting and cell size analysis
The InSexp were dissociated to single cells using 0.25% trypsin-EDTA for 10 min at 37℃. The dissociated cells were then diluted with advanced DMEM/F12 basal media and centrifuged for 5 min at 1,500 rpm. The single cells were resuspended in InSexp growth medium and stained with trypan blue. The cell counts and sizes were determined using the Countess 3Automated Cell Counter (Thermo Fisher Scientific Inc.).
Results
Establishment of the 3D InSexp culture system and characterization of marker expression
To develop a fast expandable 3D ISC culture method, we dissociated hPSC-derived HIOs into small fragments and embedded them in a Matrigel dome (Fig. 1A). Embedded cells formed morphologically distinct structures compared to HIOs and showed a hollow spherical shape (Fig. 1B). This new type of 3D intestinal organoids, designated as InSexp, is highly proliferative and easily passaged by single cell dissociation for more than 6 months without spontaneous differentiation and cell death (Fig. 1C). The InSexp is mostly composed of ISCs and progenitors, and thus exhibits higher expression of stem cell/progenitor marker genes and proteins (Fig. 1D and 1E). Furthermore, the InSexp can be cryopreserved without any loss of morphology and function upon thawing (Fig. 1B). These data suggest that the newly developed InSexp culture method enables rapid expansion and cryopreservation through the enrichment of ISCs and progenitors. Interestingly, ISC and progenitor-enriched InSexp further differentiated into 3D HIOs by re-embedding in a new Matrigel without cell dissociation, and the generation of multiple budding structures in the re-embedded InSexp is similar to the structural characteristics of the original 3D HIOs (Fig. 1F).
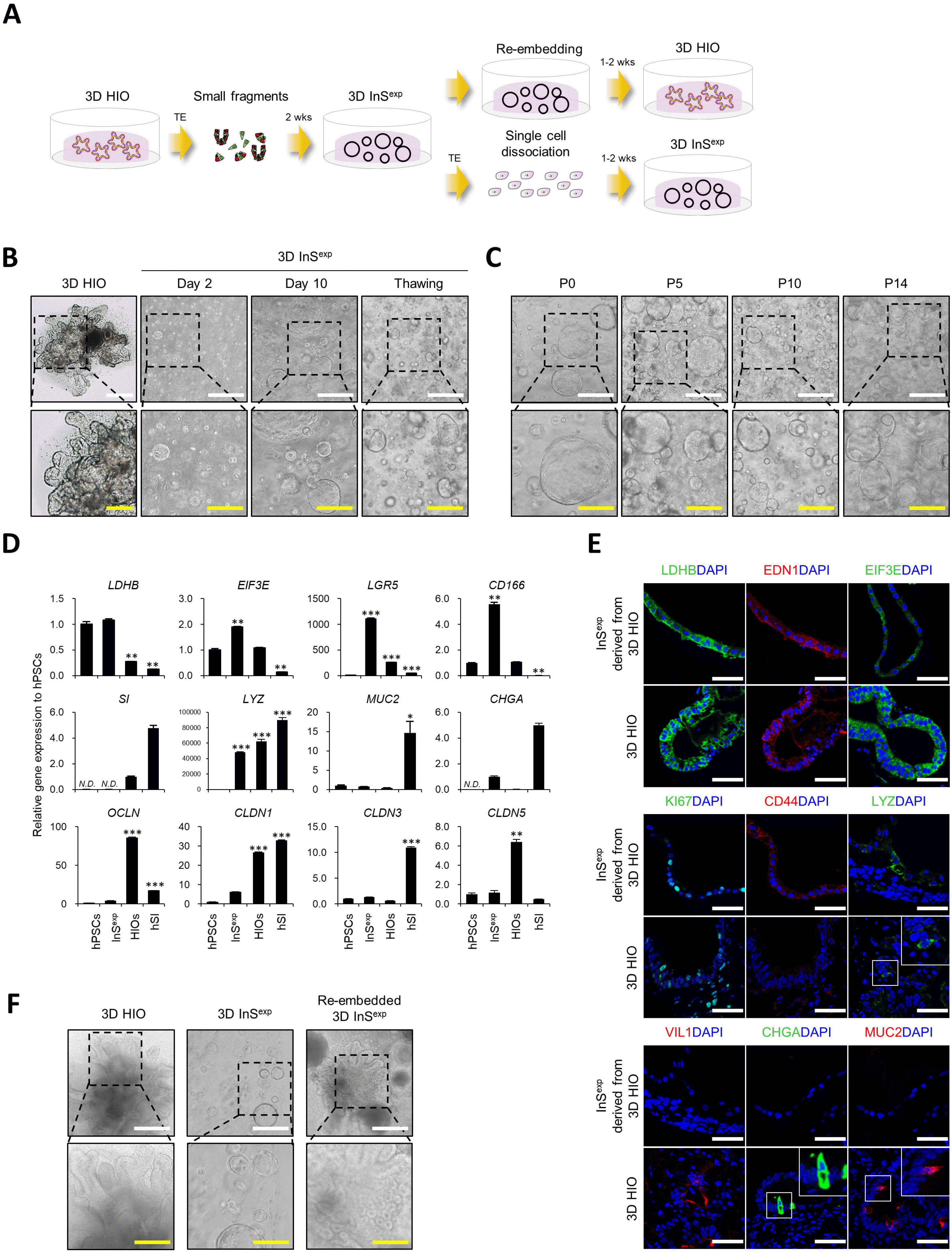
Fig. 1
Culture and characterization of InSexp derived from 3D HIOs. (A) Schematic diagram of the InSexp culture method. (B, C) Bright field images of InSexp by data, after thawing (B), and by passaging (C). White scale bars, 500 μm. Yellow scale bars, 250 μm. (D) Relative expression of intestinal epithelial cell marker genes in hPSCs, InSexp, and HIOs. (E) Immunofluorescence analysis of intestinal epithelial cell markers in InSexp. White scale bars, 50 μm. (F) Bright field images of 3D HIOs, InSexp, and re-embedded InSexp. White scale bars, 500 μm. Yellow scale bars, 250 μm. *p<0.05, **p<0.01, ***p<0.001 using two-tailed t-test.
![]()
Optimization of 3D InSexp culture conditions through WNT pathway activation
As treatment with CHIR99021 has been reported to enhance the growth of mouse and human adult stem cell-derived intestinal organoids (9, 10), we supplemented a glycogen synthase kinase 3 (GSK-3) inhibitor CHIR99021 in the InSexp culture medium to optimize the culture medium. First, we performed a titration experiment with increasing concentrations of CHIR99021 to assess the optimal concentration range of WNT activator. The growth of InSexp was maximized by treatment with 1 μM CHIR99021; however, InSexp growth was suppressed when treated with CHIR99021 at a concentration higher than 1 μM (Fig. 2A and 2B). Interestingly, treatment with CHIR99021 at more than 6 μM completely inhibited the growth of InSexp (Fig. 2A and B). As the growth of InSexp was maximized when treated with 1 μM of CHIR99021, this was the concentration used in all subsequent experiments. We next examined the effect of WNT ligand depletion on the growth of InSexp. As expected, the growth rate of InSexp was significantly decreased by depletion of WNT ligands, WNT3A and R-spondin 1, from the growth medium, and was mostly recovered by supplementation with CHIR99021 (Fig. 2C and 2D). In line with this, the spheroid formation and growth of InSexp were completely diminished by treatment with WNT pathway inhibitors such as WNT C-59 and XAV939 (Fig. 2E). Therefore, these data indicate that the WNT pathway plays an essential role in controlling the growth of InSexp, and that the growth of InSexp can be enhanced by moderate activation of the WNT pathway using 1 μM of CHIR99021. The WNT pathway was found to be essential for InSexp proliferation, as KI67+ proliferating cells disappeared under WNT-depleted or WNT inhibitor-treated conditions; however, these did not induce cell death (Fig. 2F). Taken together, we suggest that moderate activation of the WNT pathway augments the growth rate of InSexp by increasing cell proliferation, and that the growth medium was optimized by adding a low concentration of CHIR99021.
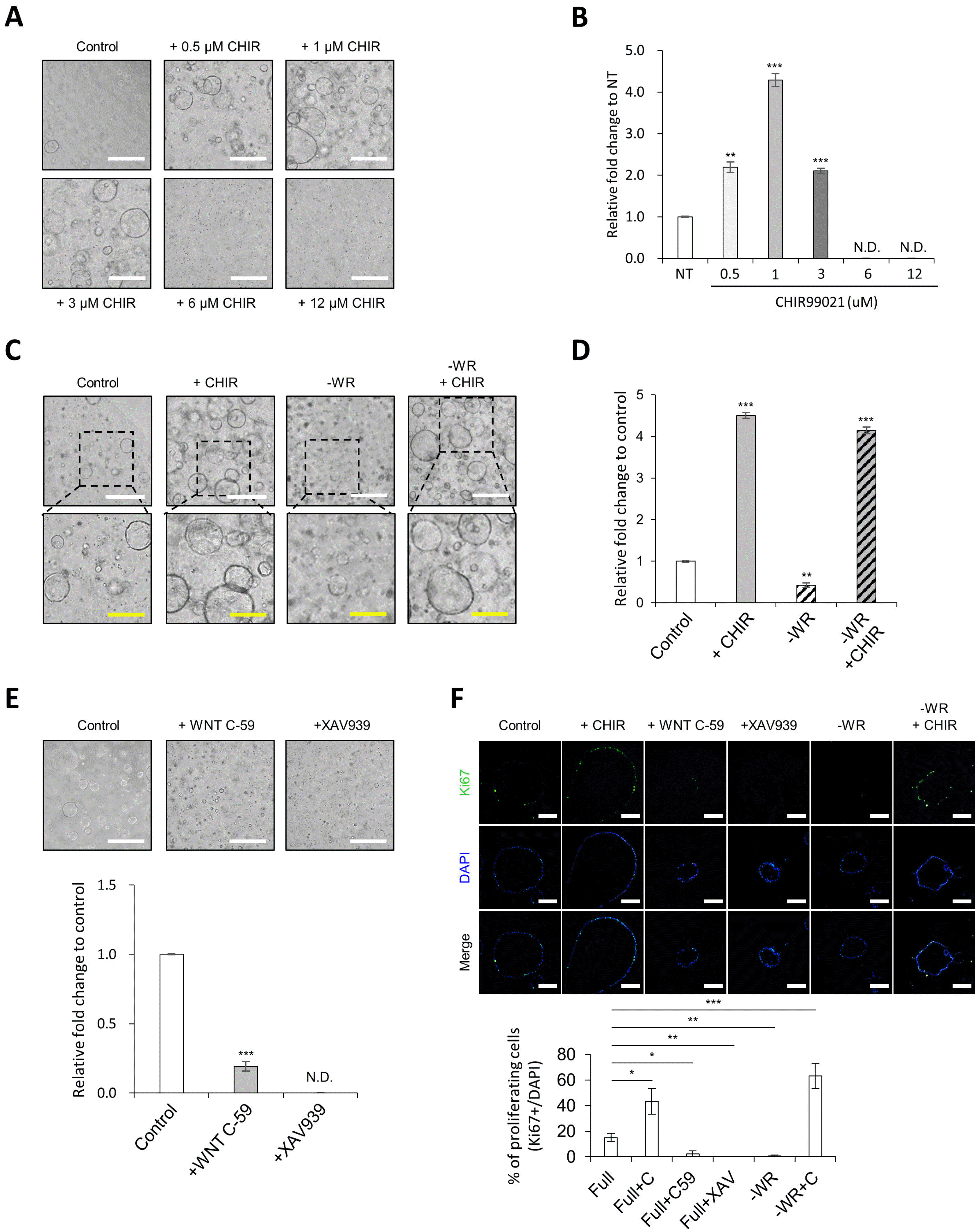
Fig. 2
CHIR99021 treatment promotes growth rate of InSexp. (A, B) Effect of CHIR99021 on the InSexp growth rate at multiple concentrations. Bright field images of the InSexp (A) and Relative fold change in the InSexp cell number (B). White scale bars, 500 μm. Data represent means±SEM (n=4). (C, D) Recovery of InSexp-forming competency by addition of CHIR99021 in WNT3A- and R-spondin 1-depleted medium. Bright field images of InSexp (C) and Relative fold change in the InSexp cell number (D). White scale bars, 500 μm. Yellow scale bars, 250 μm. Data represent means±SEM (n=4). (E) Inhibition of InSexp growth by treatment with WNT inhibitors. White scale bars, 500 μm. Data represent means±SEM (n=4). (F) Immunofluorescence analysis and quantification of proliferating cell markers in InSexp. Scale bars, 200 μm. Data represent means±SEM (n=5). *p<0.05, **p<0.01, ***p<0.001 using two-tailed t-test.
![]()
Immature and mature HIO-derived 3D InSexp display distinct profiles of maturity gene expression
We previously demonstrated that hPSC-derived hIOs can be matured in vitro using interleukin-2 treatment, and these were more similar to the adult human small intestine (2). Therefore, we investigated whether 3D InSexp derived from immature or in vitro-matured HIOs exhibited distinct marker gene expression pattern and functional characteristics. First, when the same number of cells was embedded in a Matrigel dome, the spheroid formation efficiency of mature HIO-derived InSexp was significantly higher than that of immature HIO-derived InSexp (Fig. 3A). On day 14 of culture, the total cell number of mature HIO-derived InSexp was ∼2.6 times greater compared to that of immature HIO-derived InSexp (0.78±0.0265 (×105) in immature HIO-derived InSexp vs. 2.55±0.046 (×105) in mature HIO-derived InSexp) (Fig. 3B). Interestingly, CHIR 99021 treatment showed that mature HIO-derived InSexp responded more sensitively to WNT pathway activation compared to immature HIO-derived InSexp (Fig. 3A and 3B). Furthermore, the WNT target genes were confirmed to be highly expressed in mature HIO-derived InSexp compared to that in immature HIO-derived InSexp (Fig. 3C∼E). Activation of the canonical WNT pathway governs the expression of stemness- and proliferation-related genes (11, 12); the expression of these genes and proteins including CD44, β-catenin, ASCL2 and KI67 was also higher in mature HIO-derived InSexp than that in immature HIO-derived InSexp (Fig. 3C∼E). Our findings indicate that immature and mature HIO-derived InSexp retain maturity-specific functional characteristics and gene expression profiles.
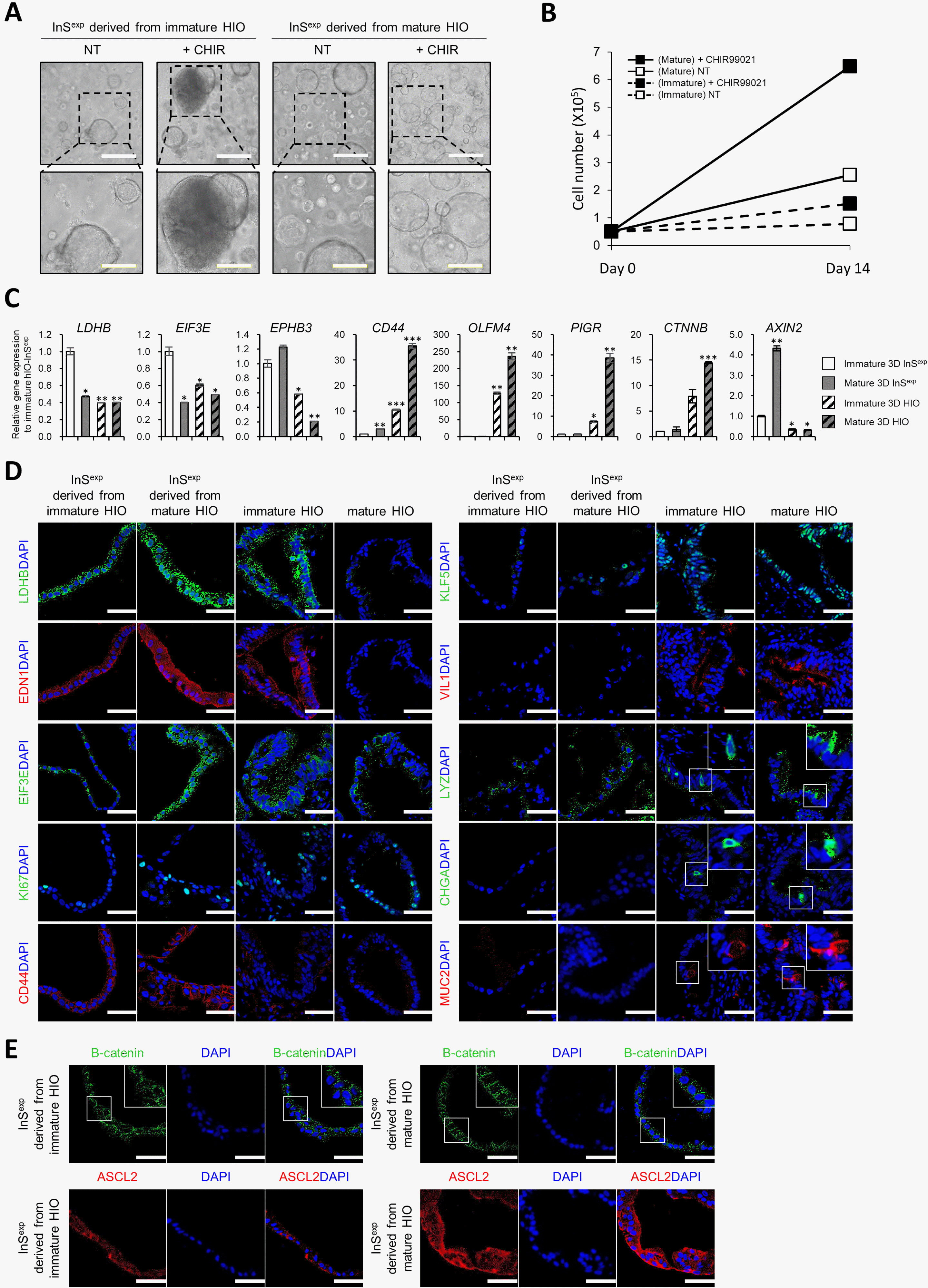
Fig. 3
Characterization of InSexp derived from immature and mature 3D HIOs. (A, B) Effect of CHIR99021 on InSexp derived from immature and mature HIOs. Bright field images of InSexp (A) and cell number increase of the InSexp after day 14 (B). White scale bars, 500 μm. Yellow scale bars, 250 μm. Data represent means±SEM (n=4). (C) Relative gene expression of intestinal epithelial cell markers and WNT target genes in InSexp derived from immature and mature HIOs. (D, E) Immunofluorescence analysis of the intestinal epithelial cell markers and WNT target genes in InSexp derived from immature and mature HIOs. White scale bars, 50 μm. *p<0.05, **p<0.01, ***p<0.001 using two-tailed t-test.
![]()
Discussion
In intestinal biology and its applications, there is an increasingly evident need for the development of various intestinal epithelial models such as 3D hIO and 2D monolayer culture systems (7, 13). Here we present a novel culture method for the scalable expansion of human ISCs enriched from hPSC-derived HIOs. Unlike 3D HIOs, the InSexp have a spherical shape without a budding and villus structure, and mostly comprise ISCs and progenitors residing in the crypt region in HIOs (Fig. 1B and 1D). Owing to the enrichment of ISCs and progenitors, InSexp can be cultured with rapid expansion kinetics and can be maintained long-term through multiple passaging and freeze-thaw cycles (Fig. 1B). The growth rate of InSexp was further increased by WNT pathway activation (Fig. 2A and 2B). Interestingly, the growth of InSexp was enhanced by treatment with a low concentration of CHIR99021, whereas it was inhibited by treatment with a high concentration of CHIR99021 (Fig. 2A and 2B). It is unclear how low and high concentrations of CHIR99021 have contrasting effects on InSexp growth; however, this phenomenon has been repeatedly observed in previous reports (10, 14). Although the reason for the distinct effects between low and high concentrations of CHIR99021 on InSexp growth remains unclear, we will elucidate the underlying molecular mechanism in future studies.
hPSC-derived HIOs are known to exhibit immature characteristics similar to the fetal intestine, and it can be matured similar to the adult intestine through in vivo transplantation or in vitro maturation (2, 15, 16). In this study, mature HIO-derived InSexp exhibited higher spheroid formation capability and growth rate compared to immature HIO-derived InSexp (Fig. 3A and 3B). The surface area of spheroids and total number of cells in the mature HIO-derived InSexp were significantly increased by treatment with CHIR99021, as they responded more sensitively to WNT pathway activation (Fig. 3A and 3B). These results suggest that the memory of the maturity of HIO is retained even in InSexp, as confirmed through the differences in the expression of the maturation marker genes between mature HIO-derived InSexp and immature HIO-derived InSexp (Fig. 3C and 3D). However, it is necessary to verify whether there is a difference in the epigenetic modifications including DNA methylation between mature HIO-derived InSexp and immature HIO-derived InSexp in further studies.
In conclusion, we established a culture method for InSexp, which can be used as a cell source for studying human ISC biology. Furthermore, as InSexp mostly comprise ISCs and progenitors and can be cryopreserved, they can be developed for use in regenerative medicine in the future, and provide an alternative platform for studying ISC biology.
Supplementary Materials
Supplementary data including two tables can be found with this article online at https://doi.org/10.15283/ijsc21209.
 XML Download
XML Download