

 PDF
PDF Citation
Citation Print
Print


Introduction
Chronic liver damage from various etiologies such as toxins, viral infections, autoimmune reactions and metabolic diseases usually follow a final common route, which is fibrosis. Chronic liver fibrosis can lead to cirrhosis, which is associated with fatal complications and sequelae. Liver fibrosis is the end result of increased synthesis and decreased degradation of extracellular matrix (ECM) proteins, the most common of which is type I collagen. ECM degradation is mediated by a family of zinc-dependent enzymes, matrix metalloproteinase (MMP) (1). Tissue inhibitor of metalloproteinase (TIMP) regulates the activity of MMP by binding to MMP and blocking their collagen degradation (2). MMP-TIMP interaction is a major regulation point for liver fibrosis and can be a therapeutic target for patients with ongoing liver fibrosis.
Stem cells for hepatocytes are categorized into extrahepatic and intrahepatic stem cells with respect to their origin. It has been reported that hematopoietic stem cells and liver-derived mesenchymal stem cells (MSCs) can differentiate into hepatocyte-like cells in vitro (3, 4). Prior animal studies have shown that MSCs may elicit protective or therapeutic effects against liver fibrosis in animal models (5). This concept has since translated to clinical trials that have applied cell therapy using autologous and allogeneic MSCs in chronic liver diseases, with varying results (6, 7).
Human liver stem cells (HLSCs) are elongated spindle cells isolated from human liver tissue that have mesenchymal morphology and express MSC markers. HLSCs possess intrahepatic stem cell properties and can differentiate into mesenchymal lineages such as osteoblasts, chondrocytes, and hepatocyte-like cells in vitro (2, 6). Here, we introduce a rat model of CCl4-induced liver fibrosis and trans-portal transplantation of HLSC. We demonstrate this procedure to effectively alleviate liver fibrosis in rats and elaborate the possible mechanism by which HLSCs elicit their anti-fibrotic effects in vivo.
Materials and Methods
Isolation of HLSCs
HLSCs were isolated from either unused deceased donor liver grafts that were unsuitable for liver transplant, or the non-tumor portion of livers from patients undergone surgical resection for benign liver tumors. Written informed consent were obtained from all participants that provided tissue samples. This study was approved by the Institutional Review Board at Samsung Medical Center (No. 2003-10-021, No. 2017-04-042). We analyzed the characteristics of HLSCs from 14 donors for this study and ultimately used HLSCs from 3 donors (HLSC7, HLSC15, HLSC26) for transplantation. Hepatocyte preparation was conducted using a minor modification of the two-step collagenase perfusion technique (8, 9). In brief, the liver was perfused via the portal vein with a warm oxygenized perfusion buffer (NaCl 8 g/l, KCl 0.4 g/l, NaH2PO4∙2H2O 0.06 g/l, Na2HPO4∙12H2O 0.06 g/l, glucose 2.7 g/l, NaHCO3 1.05 g/l, HEPES 2.38 g/l, and EDTA 0.19 g/l) and liver digestion was accomplished by recirculating a perfusion buffer supplemented with collagenase (0.3 g/l) and calcium chloride (0.735 g/l) at 37℃ to break down the ECM. The cell suspension was centrifuged three times at 50×g for 3 minutes at 4℃ to remove hepatocytes. Then the supernatant was collected from the liver cell suspensions and the cells were collected by centrifugation at 200×g for 5 minutes at 4℃. The pellet was resuspended in phosphate-buffered saline (PBS), layered on a Histopaque cushion, and centrifuged at 800×g for 20 minutes at 4℃. Cells at the interface were collected and washed. HLSCs were cultured on plastic dishes at a density of 5×104 cells per cm2 in Dulbecco’s modified eagle medium with 10% fetal bovine serum. In the experiment, we used the sixth passage HLSCs.
Generation of liver fibrosis rat model and HLSC transplantation
Sprague Dawley (SD) rats were obtained from OrientBio (Seongnam-si, Korea). All rats were bred under conditions in accordance with the ethical guidelines for the care of mice at the Laboratory Animal Research Center at Samsung Medical Center (Seoul, Korea).
The experimental scheme is outlined in Fig. 1. Cirrhosis was induced by intraperitoneal injection of Carbon tetrachloride (CCl4, Aldrich chemical Co., Milwaukee, USA) dissolved in paraffin oil (1:1). Liver fibrosis was induced in rats using 2 protocols with varying durations of CCl4 treatment (Supplementary Table S1). Rats received 0.65 ml/kg of CCl4 twice weekly for 8 weeks and underwent HLSC transplantation (1×107 cells) with simultaneous left hepatic lobectomy in the moderate toxicity protocol. CCl4 treatment was continued for 6 subsequent weeks at 0.5 ml/kg twice weekly. In the intense toxicity protocol, rats received 0.75 ml/kg of CCl4 twice weekly for 10 weeks and underwent HLSC transplantation (1×107 cells) with simultaneous left hepatic lobectomy. CCl4 treatment was continued for 4 subsequent weeks at 0.5 ml/kg twice weekly. Animals in both groups were sacrificed following completion of the 14-weeks course of CCl4 treatment before and after HLSC transplantation. Liver specimens from the sacrificed rats (n=5 in each group) were analyzed (Fig. 1). Histological and immunohistochemistry analyses of the left hepatic lobe was done to provide baseline parameters of liver fibrosis at the time of HLSC transplantation. Rats in the control group underwent 14 weeks of CCl4 treatment without HLSC transplantation.
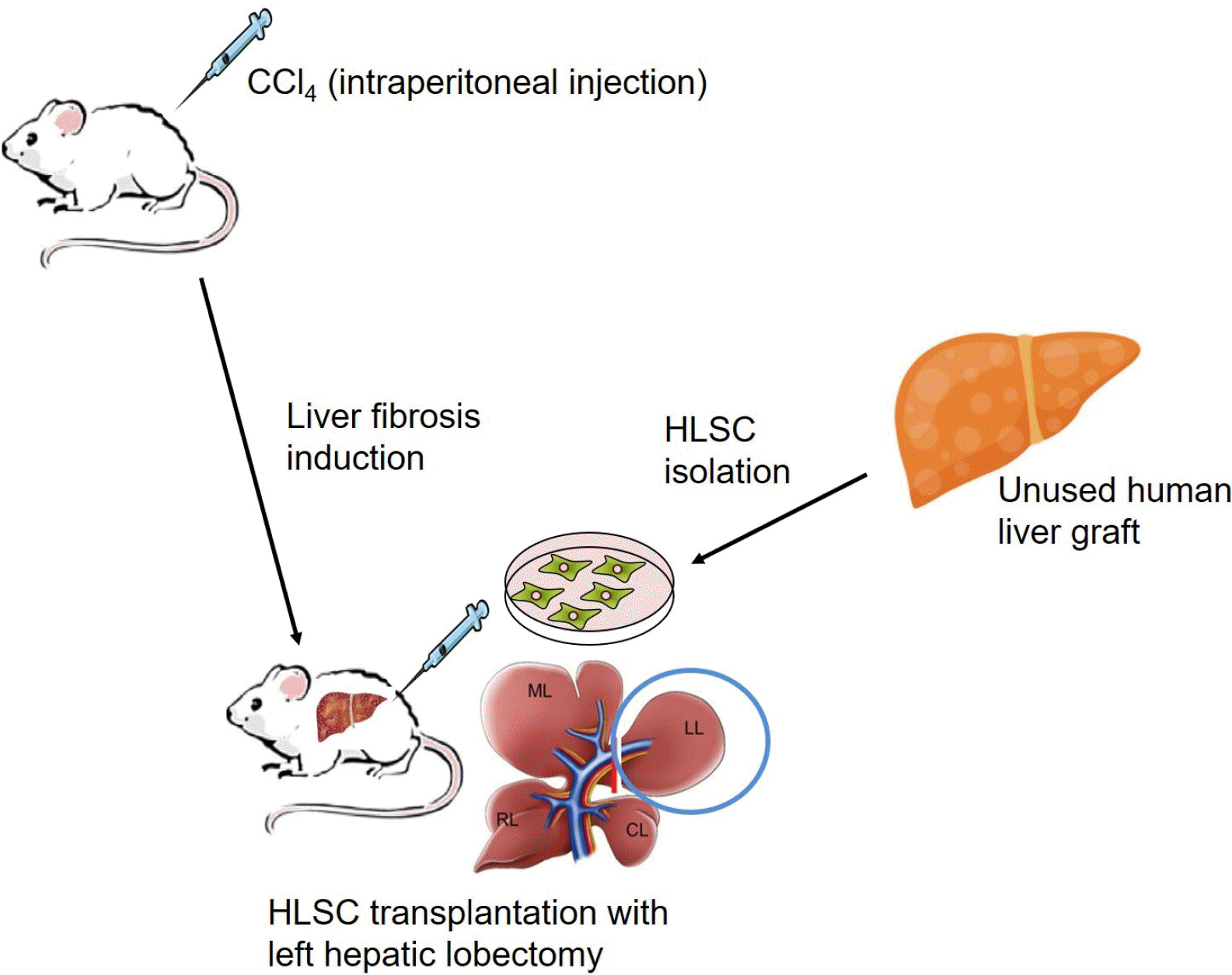
Fig. 1
Scheme for the generation of rat liver fibrosis model and HLSC transplantation. SD rats received biweekly intraperitoneal injections of CCl4 0.65∼1.0 ml/kg for up to 10 weeks. HLSCs were isolated from human liver grafts unsuitable for transplantation, as described in ‘Methods’. Rats underwent abdominal surgery trans-portal HLSC transplantation with simultaneous left hepatic lobectomy.
![]()
HLSCs were infused at a dose of 1×106 cells and saline was infused at the same volume into the portal vein. Three schemes for HLSC transplantation were applied. First was the standard transplant scheme, where 1×107 cells were transplanted once following 10 weeks of CCl4 treatment (HLSCx group). Second was a double-dose scheme, where 2×107 cells were transplanted following 10 weeks of CCl4 treatment (HLSC2x group). Third was a tandem transplant scheme, where two HLSC transplant sessions with 1×107 cells were done in 2-week intervals, following 10 weeks of CCl4 treatment (HLSCxx group). Rats continued to be treated with CCl4. After cell infusion, rats were also given cyclosporine at 5 mg/kg body weight daily for two weeks to suppress immunity. At 4 weeks after HLSC transplantation, rats were euthanized and the liver and blood sample were obtained.
Quantitative analysis of liver fibrosis
Rat liver tissue samples were immediately fixed in 10% neutral buffered formalin at 4℃ and routinely processed for paraffin embedding and cross-sectioned to obtain 4 μm-thick sections with circular layer and myenteric ganglia cut longitudinally. Before use, sections were deparaffini-zed, rehydrated and processed for routine haematoxylin/eosin and histochemical staining.
The slides were incubated with a 0.1% Direct Red 80 (Sigma-Aldrich, Missouri, USA) dissolved in aqueous saturated picric acid (Sigma-Aldrich, Missouri, USA) for 1 hour, washed in acidified water (0.5% acetic acid (glacial). Dehydrated and mounted with VECTOR Mounting (VECTOR, California, USA). Collagen and non-collagen components were red- and orange-stained, respectively.
Digital images were captured with an Aperio Scanscope System AT (Leica, Wetzlar, Germany) and the area of fibrosis was quantified in 20×-magnification fields five area per specimen. An RGB (Red, Green, Blue) threshold was used to identify the areas of fibrosis using Imagescope software (Leica, Wetzlar, Germany). Fibrosis was expressed as percentage (%) of total area.
Immunohistochemistry
For IHC, formalin-fixed rat liver tissue sections were deparaffinized and rehydrated, then heated at 100℃ for 30 min in a HIER Citrate Buffer (0.01 M), (Zytomed, Berlin, Germany) for antigen retrieval. The sliced tissues were later dealt with Hydrogen Peroxide Block (Thermo, Massachusetts, USA) to block the endogenous peroxidase activity. Next, the slides were blocked with Ultra V Block (Thermo, Massachusetts, USA). After rinsing in wash buffer, the sections were incubated with anti-alpha 1 antitrypsin antibody (Abcam, Cat. No. ab9373, Cambridge, UK), rabbit polyclonal to human serum albumin (Abcam, Cat. No. ab2406, Cambridge, UK), and monoclonal mouse anti-human cytokeratin 18 (Dako, Cat. No. M7010, Santa Clara, USA) for 1 hour at room temperature. The slides were then rinsed in wash buffer and incubated for 15 min with secondary antibody horseradish peroxidase-conjuga-ted (HRP Polymer, Thermo, Massachusetts, USA). Immune complexes were detected through the standard substrate detection of HRP. Last, the slides were stained with hematoxylin and dehydrated in graded alcohols and xylene. Digital images were captured with an Aperio Scanscope System AT (Leica, Wetzlar, Germany).
Real-time quantitative polymerase chain reaction (qPCR)
Total RNAs were extracted from fresh rat liver tissue with Trizol reagent (Invitrogen Life Technologies, Carlsberg, USA), following the manufacturer’s protocol (Invitrogen). To remove Genomic DNA, the RNA samples were treated with DNase I (Promega, Wisconstin, USA). Total RNA was extracted from HLSCs with RNeasy mini kit (Qiagen, Hilden, Germany). The A260/A280 ratio of the optical density of the RNA samples (measured with NanoDrop 1000; peQLab, Erlangen, Germany) was between 1.95 and 2.05 indicating adequate RNA quality. The reverse transcrip-tion was performed with the total RNAs (500 ng) and the Superscript III Reverse Transcriptase (Invitrogen Life Technologies, Carlsberg, USA).
Real-time qPCR was performed with the SensiFAST SYBR HI-ROX 2X Mix Kit (BioLIne, Toronto, Canada) by the 7900HT Fast Real-Time PCR system (Applied Biosystem, California, USA). The sequences of the specific primers used were as follows: Collagen 1A2 (Col1A2), 5’-TAAGGCTGTCATTCTGCAGGGC-3’ and 5’-GTCCAG AGGTGCAATGTCAAGG-3’; MMP2, 5’-CTGATAACCTG GATGCAGTCGTG-3’ and 5’-CAGCCAGTCCGATTTGA TGCTTC-3’; MMP1, 5’-AGCTGAAGGTAGAGCCAGGG TAC-3’ and 5’-AGAACGACAGCAGCTGCACTCAC-3’; TIMP1, 5’-TCCCTTGCAAACTGGAGAGTGAC-3’ and 5’-GGTATTGCCAGGTGCACAAATC-3’; TIMP2, 5’-ATCA CACGCTGCCCTATGATCC-3’ and 5’-CATCACTTCTC TTGATGCAGGC-3’; Transforming growth factor beta 1 (TGF-b1), 5’-TAGGAAGGACCTGGGTTGGAAG-3’ and 5’-CACCCGGGTTGTGTTGGTTGTAG-3’; hepatocyte growth factor (HGF), 5’-AGCTAGTCTATGGACCTGAAGGC-3’ and 5’-GTGTAGCCCCAGCCGTAAATACT-3’. Each sample contained 0.8 μl cDNA, 2X SensiFAST SYBR HI-ROX Mix, and 10 μM each primer in final volume of 20 μl. Amplification conditions were as follows: one cycle of denaturation at 95℃ for 2 minutes, 40 cycles denaturation at 95℃ for 5 seconds, annealing at 60℃ for 30 seconds, extension at 72℃ for 15 seconds, and melting curve at 55℃ with the temperature gradually increased for 0.5℃, up to 95℃. β-actin mRNA was used as an internal control for real-time qPCR. The results were analyzed with the 2−ΔΔCt method. To prove the correct lengths of the qPCR products, the samples were analyzed with agarose gel electrophoresis (Bioneer, Daejeon, Korea).
Results
Alleviation of liver fibrosis following HLSC transplantation
Gross appearance of the liver observed at the time of left hepatic lobectomy with or without HLSC transplantation (at 8 weeks following moderate toxicity protocol and at 10 weeks following intense toxicity protocol, “baseline”) showed cholestatic discoloration with surface nodularity and blunt edges. Sirius red staining of the liver specimens revealed deposition of collagen fibers in the extracellular (Fig. 2A). The liver specimens of rats were then analyzed at post-14 weeks of CCl4 treatment. In the HLSC transplantation group of both CCl4 treatment protocols, the cholestatic discoloration and surface nodularity appeared to have been alleviated and Sirius red staining showed deposition of collagen fibers to a lesser degree. Presence of hALB secreting cells were detected by immunohistochemistry in all post-14 weeks liver specimens of HLSC transplantation group. In the control group, both gross appearance of the liver and Sirius red staining showed evidence of persistent liver fibrosis (Fig. 2A).
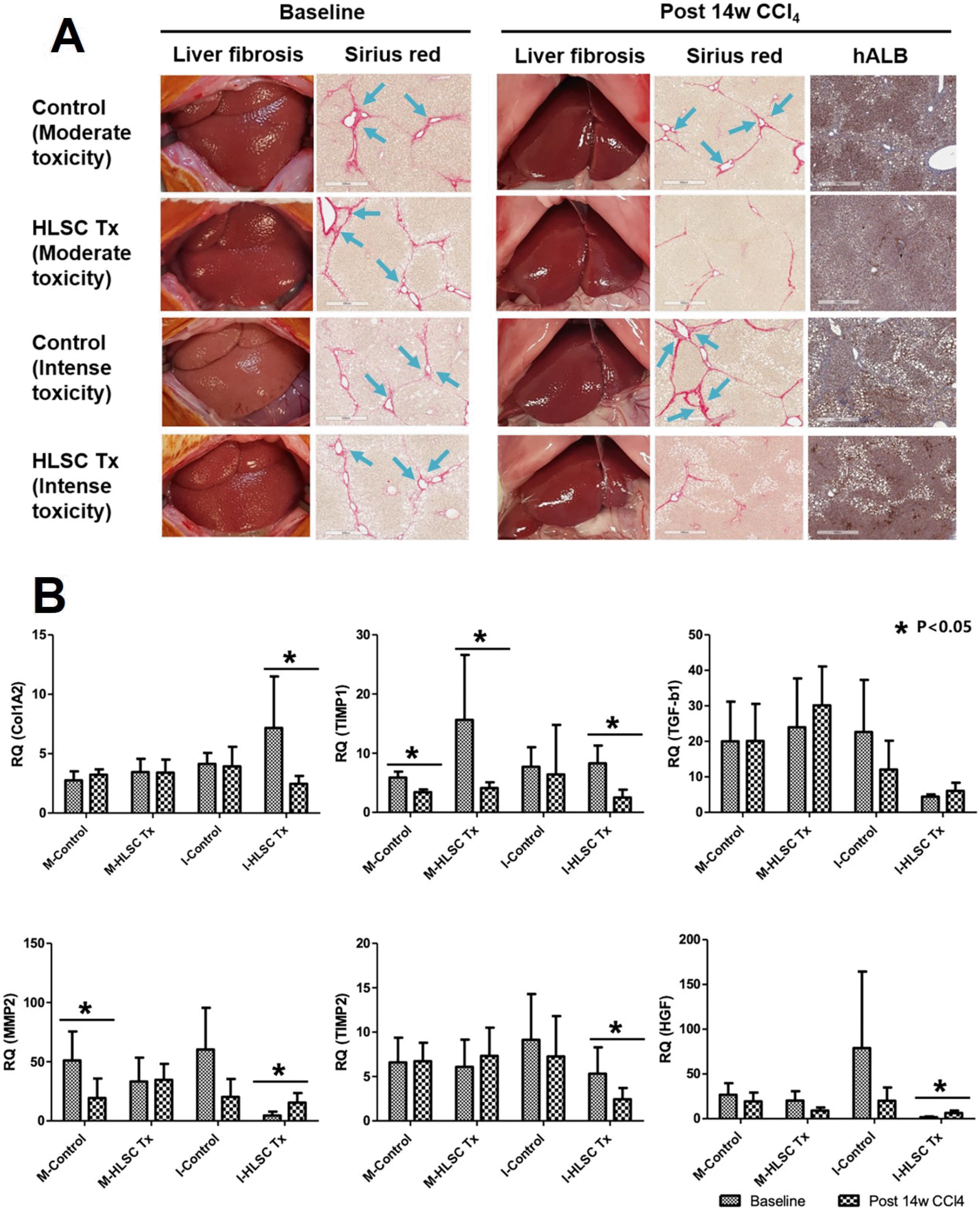
Fig. 2
(A) Therapeutic effects of HLSC transplantation in CCl4-induced liver fibrosis in rat model. Liver tissues were obtained from rats at 10 weeks (baseline) and at 14 weeks after initiation of CCl4 treatment (post 14w CCl4). Extracellular deposition of collagen fibers was stained with Sirius red. The presence of the transplanted HLSCs in the liver tissue was identified by immunohistoche-mistry using antibodies against human albumin (hALB). (B) Real-time qPCR analyses of ECM remodeling factors in rat liver specimens following two fibrosis induction protocols. Significant increase in MMP2, HGF expression and decrease in TIMP1, TIMP2 expression are seen following HLSC transplantation with intense toxicity protocol. These changes are not consistently observed in control groups.
![]()
Quantitative measurements of fibrosis area stained by Sirius red were compared between baseline and post-14 weeks liver specimens. Fibrosis area significantly decreased from baseline to post-14 weeks in the control group as well as the HLSC transplantation group in the moderate toxicity protocol (Fig. 3A). In the intense toxicity protocol, fibrosis area increased in the control group during the final 4 weeks of CCl4 treatment. However, fibrosis area significantly decreased from baseline to post-14 weeks in the HLSC transplantation group in the intense toxicity protocol (Fig. 3A).
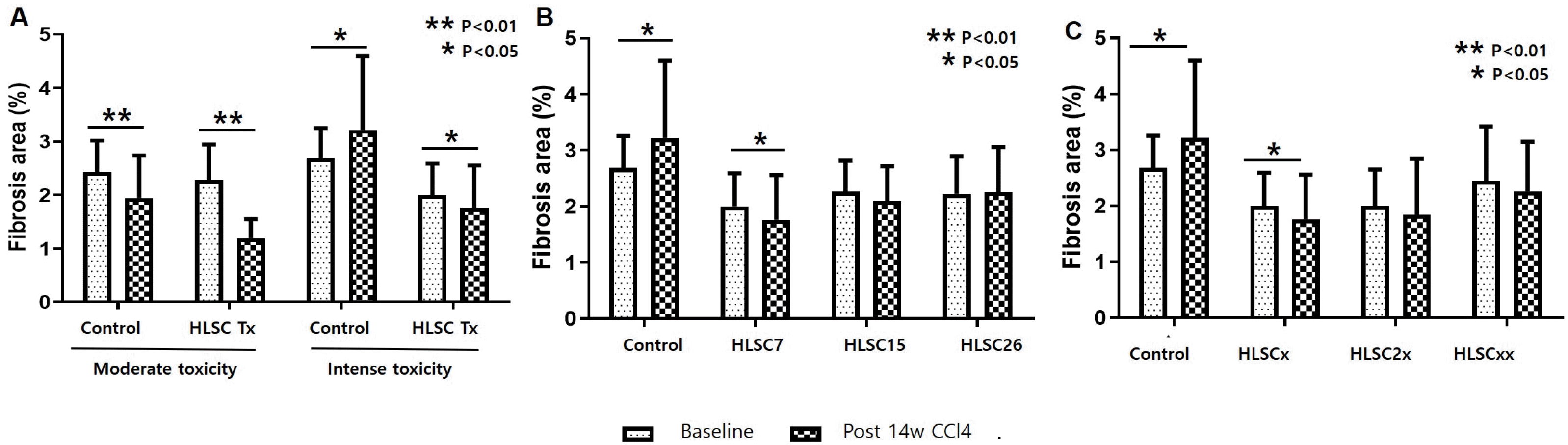
Fig. 3
(A) Changes in fibrosis area following two fibrosis induction protocols. Significant decrease in fibrosis area compared to baseline is seen at 14 weeks after liver fibrosis induction when HLSC transplantation is performed. In the control group, fibrosis area is increased from baseline to week 14 when CCl4 is given with the intense toxicity protocol. In the moderated toxicity protocol, spontaneous regression of liver fibrosis is observed at 14 weeks. (B) Changes in fibrosis area according to transplantation of three HLSC strains. Significant decrease in fibrosis area compared to baseline is seen at 14 weeks after liver fibrosis induction with HLSC7 transplantation. No significant change is seen in the other two HLSC strains. In the control group, fibrosis area is increased from week 10 to week 14. (C) Changes in fibrosis area following three transplantation schemes. Significant decrease in fibrosis area compared to baseline is seen in HLSCx (standard transplantation scheme) 14 weeks after liver fibrosis induction. No significant change is seen in the other two transplant schemes. In the control group, fibrosis area is increased from week 10 to week 14.
![]()
RNA expression in liver tissues at baseline and post-14 weeks were analyzed (Fig. 2B). TIMP1 and TIMP2 expression were significantly decreased, while hepatocyte growth factor (HGF) expression was increased following HLSC transplantation in liver tissues of rats in the intense toxicity protocol.
Effect of HLSC transplantation according to HLSC strains
Gene expression levels of factors contributing to ECM remodeling were analyzed among the HLSCs from multiple donors (Supplementary Fig. S1). Compared to MSCs, HLSCs expressed elevated levels of MMP1, TIMP2, and HGF, while MMP2, TIMP1, and TGF-b expression were decreased. Three HLSC strains (HLSC7, HLSC15, HLSC26) were selected for transplantation based upon elevated expression of MMP1 and decreased expression of TIMP1, MMP2, and TGF-b. An overall net effect towards ECM degradation by HLSCs may be expected with this pattern of mRNA expression. The comparative effects of transplantation of three HLSC strains were analyzed in rats undergone liver fibrosis induction using the intense toxicity protocol.
The liver specimens of rats (n=5 in each group) were analyzed at post-14 weeks of CCl4 treatment (Fig. 4A). In the three HLSC transplantation groups, the cholestatic discoloration and surface nodularity appeared to have been alleviated and Sirius red staining showed deposition of collagen fibers to lesser degrees. Presence of hALB secreting cells were detected by immunohistochemistry in post-14 weeks liver specimens of all three HLSC strains. Fibrosis area significantly decreased from baseline to post-14 weeks following HLSC transplantation in HLSC7 group (p<0.01). Fibrosis area significantly increased in the control group from baseline to post-14 weeks (Fig. 3B). AST and ALT levels significantly decreased after 4 weeks following HLSC transplantation of all three HLSC strains. APRI, which is a surrogate marker for fibrosis, significantly decreased after 4 weeks following HLSC transplantation of all three HLSC strains. These changes were not observed in the control group (Table 1). RNA expression in liver tissues at baseline and post-14 weeks were analyzed (Fig. 4B). TIMP1 and TIMP2 expression were significantly decreased, while HGF expression was increased following HLSC7 and HLSC26 transplantation.
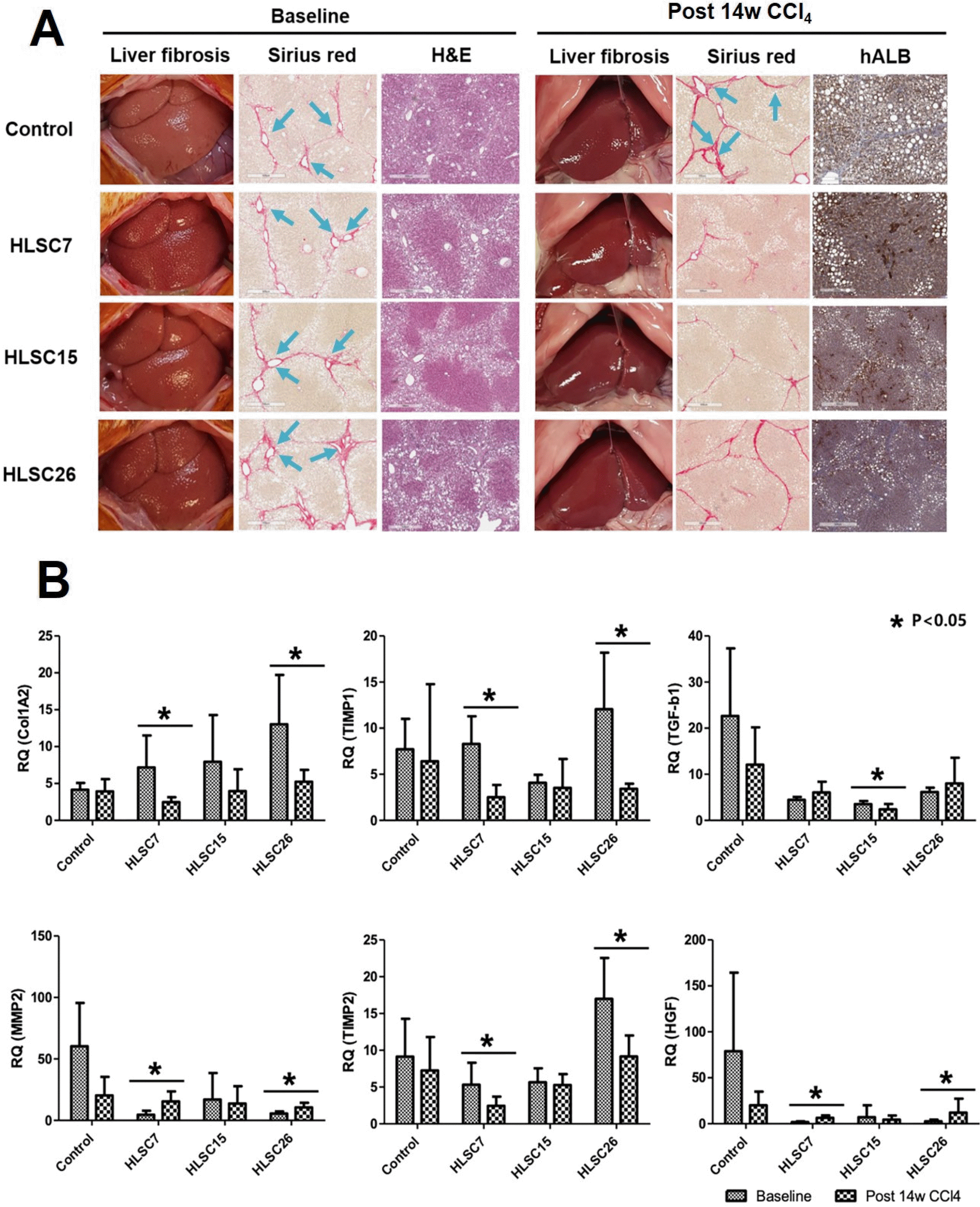
Fig. 4
(A) Therapeutic effects of HLSC transplantation with various HLSC strains. Each strain of HLSCs were transplanted following 10 weeks of CCl4 treatment. Liver tissues were obtained from rats at 10 weeks (baseline) and at 14 weeks after initiation of CCl4 treatment (post 14w CCl4). Extracellular deposition of collagen fibers was stained with Sirius red. Histopathological changes of liver sections were observed by staining with hematoxylin and eosin (H&E). The presence of the transplanted HLSCs in the liver tissue was identified by immunohistoche-mistry using antibodies against human albumin (hALB). (B) Real-time qPCR analyses of ECM remodeling factors in rat liver specimens according to transplantation of three HLSC strains. Significant increase in MMP2, HGF expression and decrease in TIMP1, TIMP2 expression is seen following transplantation with HLSC7. These changes are not consistently observed in HLSC15 or HLSC26 stains.
![]()
Table 1
Biochemical markers of liver fibrosis and inflammation according to HLSC strains and HLSC transplantation scheme
| AST | ALT | APRI | ||||||
|---|---|---|---|---|---|---|---|---|
| Baseline | Post 14w CCl4 | Baseline | Post 14w CCl4 | Baseline | Post 14w CCl4 | |||
| Control | 341.50 | 117.17 | 188.50 | 49.17* | 0.66 | 0.48 | ||
| HLSC7 | 573.50 | 93.00* | 742.33 | 65.67* | 1.06 | 0.24* | ||
| HLSC15 | 411 | 139* | 437 | 85* | 0.76 | 0.25* | ||
| HLSC26 | 538 | 105* | 629 | 56* | 0.98 | 0.25* | ||
| Control | 341.50 | 117.17 | 188.50 | 49.17* | 0.66 | 0.48 | ||
| HLSCx | 573.50 | 93.00* | 742.33 | 65.67* | 1.06 | 0.24* | ||
| HLSC2x | 436.00 | 92.40 | 465.80 | 49.60 | 0.93 | 0.28 | ||
| HLSCxx | 262.40 | 120.00 | 249.40 | 54.40* | 0.50 | 0.18 | ||
![]()
Effect of HLSC transplantation according to HLSC transplantation schemes
The liver specimens of rats (n=5 in each group) were then analyzed at post-14 weeks of CCl4 treatment (Fig. 5A). Fibrosis area decreased from baseline to post-14 weeks following HLSC transplantation with all three schemes (HLSCx, HLSC2x, HLSCxx) compared to a significant increase in fibrosis area in control group (p<0.05). However, only HLSCx group showed significant decrease in fibrosis area following HLSC transplantation (p<0.05, Fig. 3C). AST, ALT levels and APRI significantly decreased after 4 weeks following HLSC transplantation in HLSCx scheme. No significant decrease in APRI were observed in the control group (Table 1). TIMP1 and TIMP2 expression were significantly decreased, while HGF expression was increased following single transplant of HLSC (HLSCx). TGF-b expression were also significantly decreased in this group (Fig. 5B).
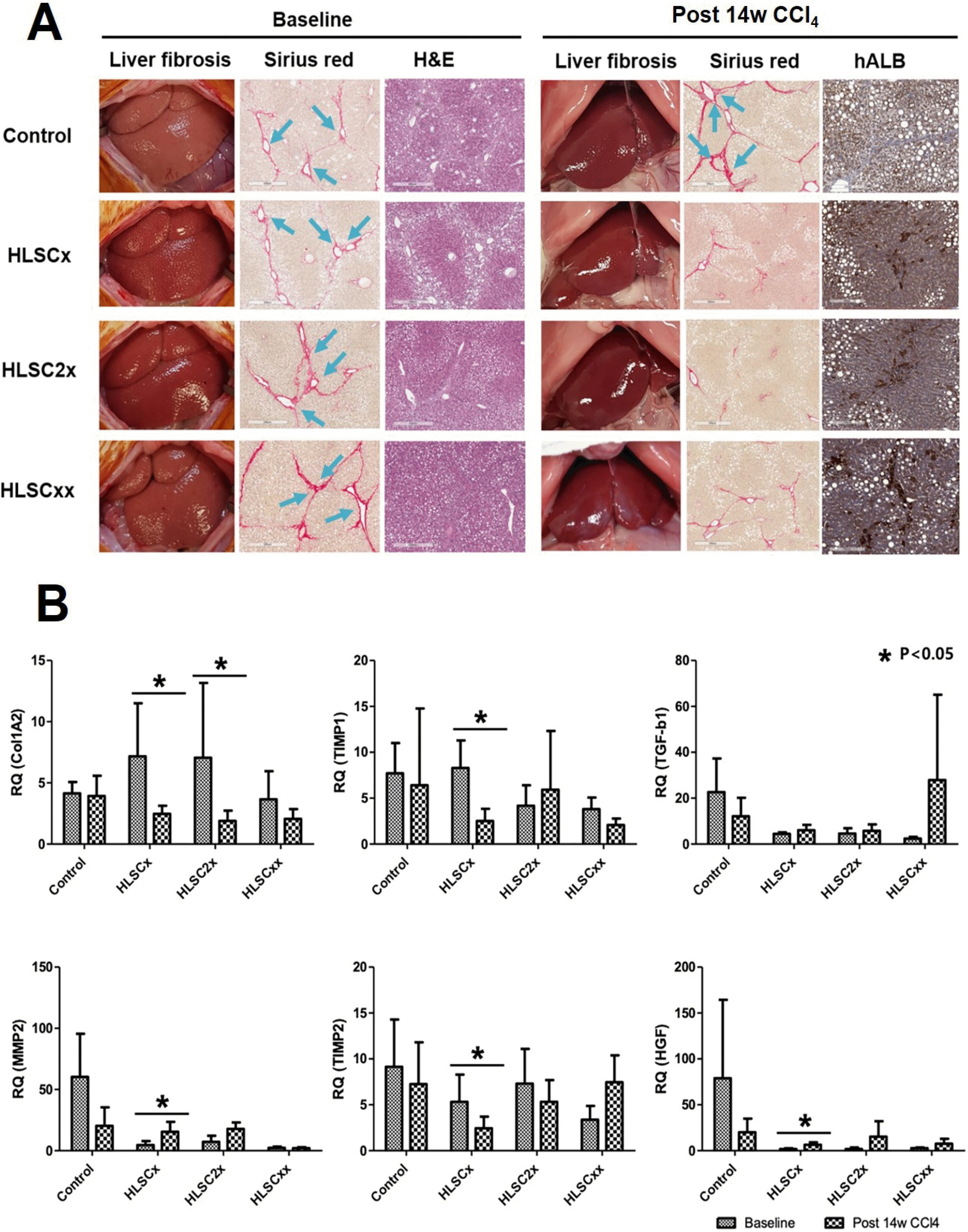
Fig. 5
(A) Therapeutic effects of HLSC transplantation according to transplant schemes. HLSCx group received a single transplant of 1×107 cells. HLSC2x group received a single transplant of 2×107 cells. HLSCxx group received 2 HLSC transplants with 1×107 cells, 2 weeks apart. Liver tissues were obtained from rats at 10 weeks (baseline) and at 14 weeks after initiation of CCl4 treatment (post 14w CCl4). Extracellular deposition of collagen fibers was stained with Sirius red. Histopathological changes of liver sections were observed by staining with hematoxylin and eosin (H&E). The presence of the transplanted HLSCs in the liver tissue was identified by immunohistoche-mistry using antibodies against human albumin (hALB). (B) Real-time qPCR analyses of ECM remodeling factors in rat liver specimens following three transplantation schemes. Significant increase in MMP2 and HGF expression is seen following HLSC transplantation. Significant decrease in TIMP1 and TIMP2 expression is seen following HLSC transplantation. These changes are not consistently observed in HLSC2x or HLSCxx schemes.
![]()
Discussion
Liver fibrosis is the end result of increased synthesis and decreased degradation of ECM proteins, the most common of which is type I collagen. Activation of hepatic stellate cells (HSC) acts as a major source of type I collagen in liver fibrosis. Following fibrous noxious stimulation, HSCs are activated and increase the synthesis of procollagen type I mRNA, resulting in excessive accumulation of ECM and scarring of tissues (10, 11). ECM degradation is mediated by a family of zinc-dependent enzymes, MMPs. MMP1 and MMP13 (interstitial collagenases) degrade type I collagen, whereas MMP2 and MMP9 (gelatinases) regulate the accumulation of type IV collagen (1, 12). TIMPs regulate the activity of MMP by binding to MMPs and blocking their collagen degradation (1, 2). TIMP mRNA and protein levels drastically increase during fibrosis, while MMP levels remain relatively stable (13, 14). The final effect is a rapid increase in ECM synthesis, along with increased TIMP activity and fibrous scar formation. In addition, TIMP1 expression was related to the inhibition of HSC apoptosis in vitro and the persistence of HSCs activated in vivo (15, 16). Therefore, MMP-TIMP interaction is a major regulation point for liver fibrosis and can be a therapeutic target for patients with ongoing liver fibrosis.
The outcomes of this study demonstrate that intraportal injection of human liver-derived cells with MSC-like features can effectively reduce the degree of fibrosis in a rat model of CCl4-induced liver fibrosis. As with MSCs from other tissues, liver-derived human MSCs possess pro-angiogenic, anti-inflammatory, and anti-apoptotic properties. In addition to these features that they share with other MSC types, liver-derived MSCs have the distinct potential to differentiate into hepatocyte-like cells that are able to produce cytochrome P450, albumin, and urea (1, 2, 17). Thus, liver-derived MSCs have recently drawn much interest as a potential therapeutic source for various liver conditions including acute liver injury, liver failure, and liver regeneration (18-21).
The role of MSC in liver fibrosis has not been fully understood. It is believed that MSC elicits its antifibrotic effects by suppression of HSC activation and secretion of cytokines and enzymes that promote ECM degradation (14, 22-24). The therapeutic potential of MSCs isolated from human bone marrow, adipose tissue, and umbilical cord in relieving liver fibrosis have been shown in rat and mice models (3, 25, 26). Compared to that from other sources of MSCs, the medium of liver-derived MSCs contain higher levels of HGF, TNF-a, and IFN-g which are mediators of HSC suppression and ECM degradation (23, 27-29). Thus, it may be postulated that liver-derived MSCs would be a more effective source of cell therapy against liver fibrosis. However, there are no published studies to date that have addressed the therapeutic potential of liver-derived MSCs in liver fibrosis.
The cells used in our experimental model are spindle-shaped cells of human liver origin with properties in close resemblance to MSCs (2, 7, 30). HLSCs isolated from various donor livers tended to express higher levels of MMP1, HGF and lower levels of MMP2, TIMP1, and TGF-b (Supplementary Fig. S1). An overall net effect towards ECM degradation of HLSCs could be hypothesized with this pattern of mRNA expression. In fact, the net anti-fibrotic effect was demonstrated as 1) improvement in the gross appearance of the liver and histologic findings, 2) in improvements in biomarkers of liver fibrosis and inflammation, and most importantly 3) as significant reduction in calculated fibrosis area following HLSC transplan-tation. The net therapeutic effect of HLSC transplantation in terms of quantitative reduction in fibrosis area were uniformly demonstrated in models of more severe liver damage (Fig. 3A) and with varying strains of HLSCs from different donor livers (Fig. 3B). However, real-time qPCR analyses did not show a consistent trend in the expression of factors related to ECM synthesis or degradation. This inconsistency may indicate that there are other important factors and players in the interaction of HLSCs and liver fibrosis. Further studies should aim to elaborate the relation between HLSC and liver fibrosis in greater detail.
In conclusion, we developed a rat model of CCl4-indu-ced liver fibrosis and HLSC transplantation via the portal vein. This procedure was effective in reducing the area of liver fibrosis and was accompanied by elevation of increased expression of factors promoting ECM degradation. These findings indicate the potential therapeutic role of HLSCs in various liver diseases presenting with liver fibrosis.
Supplementary Materials
Supplementary data including one table and one figure can be found with this article online at https://doi.org/10.15283/ijsc21031.
 XML Download
XML Download