

 PDF
PDF Citation
Citation Print
Print


Introduction
In acute myocardial infarction (MI), atherosclerotic vascular occlusions trigger pathological injury owing to myocardial ischemia and hypoxia. Angiogenesis and vascular remodeling play essential roles in affording protection against myocardial ischemia (1). Notably, angiogenesis primarily decelerates the development of heart failure (2). Percutaneous coronary intervention (PCI) and coronary artery bypass grafting have failed to rescue microvascular injury and reportedly result in PCI-related microvascular damage (3). Thus, new medical approaches have revealed a critical role for angiogenesis against progressive cardiac deterioration during cardiovascular diseases. In previous studies, as well as in our present study, human bone marrow mesenchymal stem cells (BMSCs) improved angiogenesis through a paracrine effect, primarily rescuing cardiac function in the MI mouse model (4).
Irisin, a new multifunctional myokine related to metabolic factors, is predominantly released from skeletal muscle and myocardium. Irisin is known to be involved in cell physiology and pathology, reportedly associated with the browning of white adipocytes and increased energy expenditure (5). Irisin is cleaved from fibronectin type-III domain-containing protein 5 (FNDC5) and released into the circulation, which is reportedly involved in energy metabolism in skeletal muscle, adipose tissue, and pancreatic islets (6). In a transverse aortic constriction (TAC)-induced cardiac hypertrophy mouse model, irisin reportedly alleviated cardiac fibrosis and hypertrophy, thus improving cardiac function through 5’-adenosine monophosphate-acti-vated protein kinase (AMPK)-mammalian target of rapamycin (mTOR) signaling (7). Similarly, irisin attenuates cardiac fibrosis in TAC-induced hypertrophy by increasing autophagy (8). Furthermore, irisin is involved in the angiogenesis of human umbilical vein endothelial cells (HUVECs), thus protecting cardiac function after MI (9, 10). Recent studies have reported that irisin regulates the crosstalk between adipose tissue and muscle via paracrine signaling (10). However, the role of irisin in the paracrine signaling of BMSCs needs to be elucidated. In the present study, we hypothesized that irisin could enhance the paracrine effect of BMSCs on angiogenesis to improve cardiac function post-MI injury.
Materials and Methods
Animal experiment
All animal procedures were conducted in accordance with the guidelines of the Animal Use Committee of the Guizhou Provincial People’s Hospital. To establish the MI mouse model, the left anterior descending coronary artery of 8∼12-week old male C57BL/6J mice (Guizhou Medical University) was ligated as previously described (11). BMSCs with/without irisin treatment (1.5×105 cells suspended in 20 μl Dulbecco’s Modified Eagle Medium [DMEM] per mouse) were immediately transplanted into the border zone at four different sites. The DMEM group received DMEM without BMSCs (mice received an equivalent volume of DMEM).
Human BMSCs isolation and culture
Based on a previously reported protocol (11), human BMSCs were isolated and cultured from patients who underwent hip replacement upon receipt of informed consent and approval by the Human Ethics Committee of Guizhou Provincial People’s Hospital.
BMSCs at 4-8 passage were treated with irisin (200 ng/ml; Phoenix Pharmaceuticals, USA) and LY294002 (50 μM; Selleck Chemicals, USA), as previously reported (10), for 24 h under normoxic conditions. Dimethyl sulfoxide (DMSO), the solvent employed for irisin, was used as a control.
Immunostaining
Twenty-eight days post-MI, mice were sacrificed, and their hearts were harvested and embedded in O.C.T. compound (Sakura Finetek, USA). Heart tissue sections (7-μm thick) were fixed using 4% paraformaldehyde. After permeabilization with 0.2% Triton, tissue slices were blocked with 3% bovine serum albumin (BSA) and incubated with primary antibody at 4℃ overnight. Sections were incubated with the secondary antibody at room temperature for 1 h and then observed using a fluorescence microscope (Leica, Germany). The vascular density at border areas was calculated as the positive number of vascular markers per area for one high-power field (34.68 μm2), with measurements performed in at least five random high-power fields for each section.
Sirius Red staining
Twenty-eight days post-MI, mice were euthanized, and hearts were harvested and embedded in paraffin. After sectioning (3-μm thick sections), the cardiac tissue was stained using Sirius Red (Solaribio, Beijing, China), according to the manufacturer’s instructions. Infarct areas were measured using Image-Pro software (Media Cybernetics, USA) and assessed according to the sum of the endocardial and epicardial length of the infarct zone in proportion to the sum of the endocardial and epicardial length of the entire left ventricle.
Tube formation assay
Tube formation was assessed using HUVECs, co-cultured with the supernatant of BMSCs treated with irisin, LY294002, irisin+LY294002, or control. HUVECs were seeded at a density of 5×104 cells per well and co-cultured with the BMSC supernatant. Tube formation images were acquired 4 h after transplantation, and the number of bran-ch points was quantified using the Image-Pro software.
Scratch wound-healing assay
The wound-healing assay was performed as previously described (10). Briefly, HUVECs were cultured in a 6-well plate (1×106 cells/well) for 48 h under normal conditions. After scratching the surface with a sterile 10-μl pipette tip, HUVECs were washed twice and cultured in DMEM (containing 1% fetal bovine serum [FBS]) for 24 h, containing an equal volume of BMSC supernatant treated with irisin, LY294002, irisin+LY294002, or an equivalent volume of DMSO. The wound areas were photographed with an inverted microscope (Leica, Germany), and cell migration was calculated as the average wound area in five random fields, compared with the zero-time point, using Image-Pro software.
Cell apoptosis assay
Following treatment with irisin for 24 h under normal conditions, BMSCs were cultured in DMEM with glucose and FBS deprivation under hypoxia (95% air/5% CO2, at 37℃) for 24 h.
TUNEL staining assay
Apoptosis was detected using the TUNEL Apoptosis Assay Kit (Beyotime Biotechnology, Shanghai, China) according to the manufacturer’s instructions. After fixation with 4% paraformaldehyde for 30 min, BMSCs were permeabilized with 0.2% Triton for 5 min at room temperature, followed by incubation with TUNEL reaction reagent for 1 h at 37℃ in the dark. Images were captured using a fluorescence microscope. Apoptotic cells were calculated as TUNEL-positive cells in proportion to the total number of cells.
Western blotting
Western blotting was performed to detect protein levels of p-PI3K, PI3K, p-Akt, and Akt. In brief, the proteins were obtained from cells using RIPA lysis buffer on ice, followed by centrifugation for clarification. Western blotting was performed as previously described (12).
Echocardiography
Echocardiography was performed to assess cardiac function on day 28 post-MI. After isoflurane inhalation, the mice were anesthetized and bound. Then, two-dimensional images were obtained using a Vevo 2100 system (VisualSonics, Inc., Toronto, Canada).
Statistical analysis
All experiments were independently performed at least three times, and data values are presented as the mean± standard error of the mean. Student’s t test was used to compare any two groups, and more than three groups were compared using one-way analysis of variance. *p<0.05 was deemed statistically significant.
Results
Irisin enhanced the protection of BMSCs in the MI mouse model
The therapeutic efficacy of BMSCs in ischemic heart disease has been clarified in several animal studies and clinical trials (13, 14). To confirm the protective role of irisin toward BMSCs, we treated BMSCs with irisin (200 ng/ml) for 24 h under normoxic conditions (BMSCs+irisin). DMSO, the solvent used for irisin, was used as the control (BMSCs+Con). In the MI mouse model, we transplanted BMSCs treated with irisin into the border zone immediately after MI surgery. Twenty-eight days post-MI, improved heart function was noted, with an increased ejection fraction (EF) and fractional shortening (FS), using two-dimensional echocardiography in the BMSCs+irisin group when compared with the BMSCs+Con and DMEM groups (Fig. 1a and 1b). Moreover, the infarct areas were examined by Sirius Red staining, presenting reduced infarct areas in the BMSCs+irisin group when compared with the BMSCs+Con and DMEM groups (Fig. 1c and 1d). These results revealed that irisin protected BMSCs in response to ischemic insults in vivo.
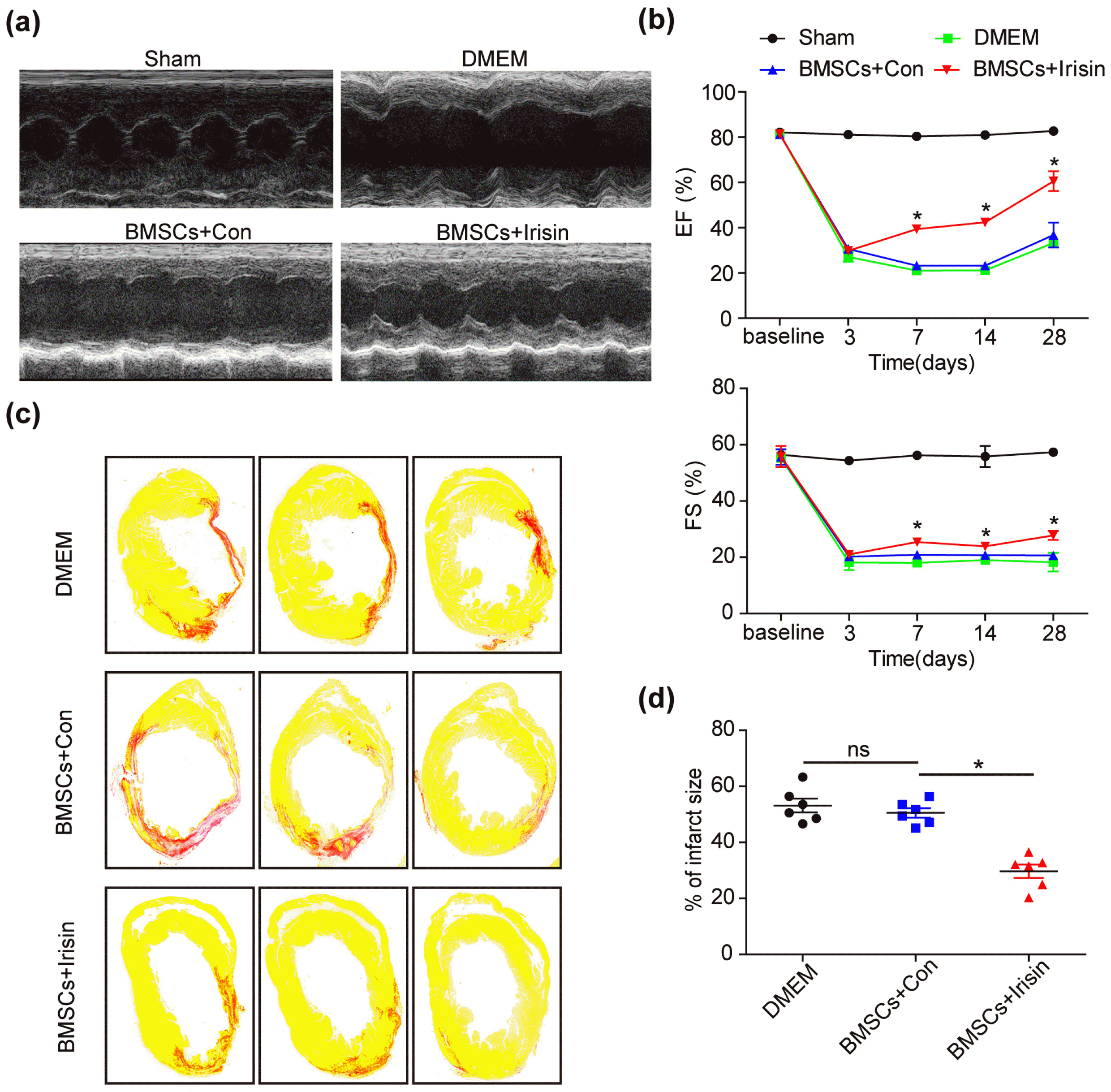
Fig. 1
Irisin-treated BMSCs rescued the cardiac function in MI mouse in vivo. (a) Representative images of echocardiography showing the improved cardiac function in BMSCs treated with irisin. (b) The ejection fraction (EF) and fractional shortening (FS) are gradually recovered in the BMSCs+irisin group when compared with other groups (n=8 for Sham, 5 for DMEM, 6 for BMSCs+irisin and BMSCs+Con). (c) The infarct area was examined by Sirius Red staining 28 days post-MI (n=6 for every group). (d) The infarct size in hearts was quantified using Image-Pro. All data were measured as mean±standard error of the mean (SEM). *p<0.05. BMSCs, bone marrow mesenchymal stem cells; MI, myocardial infarction; DMEM, Dulbecco’s Modified Eagle Medium.
![]()
Irisin regulated angiogenesis of BMSCs in vivo
The angiogenic efficacy of BMSCs mediated via paracrine signaling has been reported (4). Additionally, the role of irisin in angiogenesis was previously verified (9, 10). For further evaluation, we investigated the ability of irisin-regulated BMSC paracrine signaling to induce angiogenesis. On day 28 post-MI modeling, alpha-smooth muscle actin (α-SMA), a marker of vascular smooth muscle cells, was significantly elevated in the BMSCs+irisin group when compared with the BMSCs+Con and DMEM groups (Fig. 2a). Similarly, vascular endothelial cells markers, including von Willebrand factor (vWF) and CD31, were considerably enhanced in the BMSCs+irisin group when compared with the BMSCs+Con and DMEM groups (Fig. 2b and 2c). Interestingly, these vascular markers were marginally increased in the BMSCs+Con group when compared with the DMEM group. Blood vessels in the border zone were quantified (Fig. 2d), indicating that irisin further promoted the angiogenic efficiency in BMSCs, despite the moderate efficiency of BMSCs in angiogenesis.
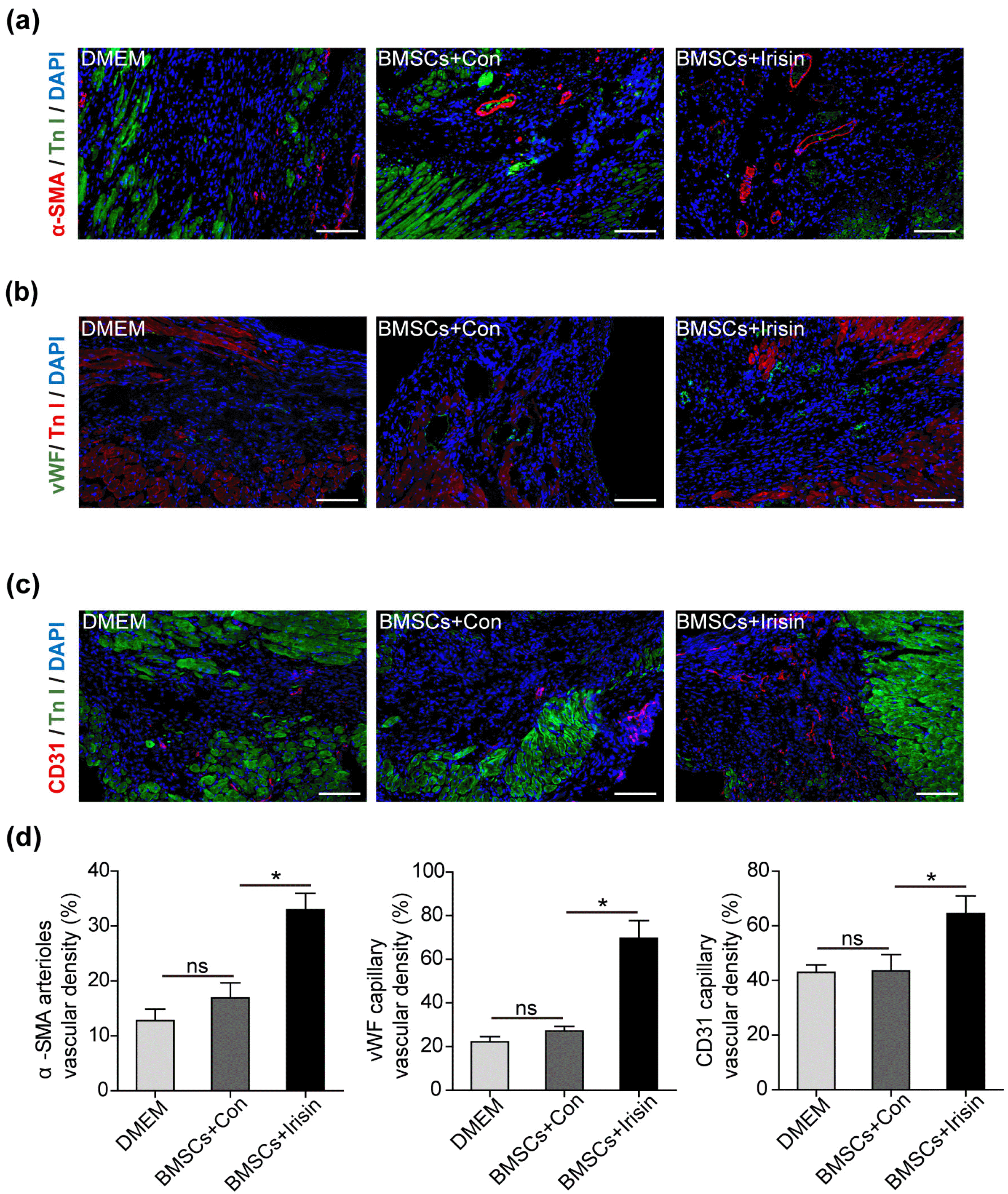
Fig. 2
Irisin-treated BMSCs enhance angiogenesis. (a∼c) Representative images of vascular markers including α-SMA, vWF and CD31 were captured in the border zone of irisin-treated BMSCs, control, and DMEM groups. (d) The vascular density was measured in at least 5 high-power fields per section (n=8 for DMEM, 7 for BMSCs+irisin and BMSCs+Con). Scale bar, 100 μm. Data are presented as mean±standard error of the mean (SEM). *p<0.05. BMSCs, bone marrow mesenchymal stem cells; DMEM, Dulbecco’s Modified Eagle Medium; α-SMA, alpha-smooth muscle actin; vWF, von Willebrand factor.
![]()
Irisin mediated the paracrine effects of BMSCs by primarily regulating the PI3K/Akt pathway
Previous studies have shown that the paracrine effects of BMSCs influence cellular fate (4, 15). To determine the role of irisin in mediating paracrine effects of BMSCs, conditioned media of BMSCs, with or without irisin, were collected; then, tube formation of HUVECs was assessed using the Matrigel assay, with cell migration examined using a wound-healing assay. In the BMSCs+irisin group, tube formation (Fig. 3a and 3c) and migration (Fig. 3b and 3d) of HUVECs were significantly increased when compared with the BMSCs+Con and irisin alone groups, but not in control alone; no difference was observed between the BMSCs+Con group and irisin alone group. The results indicated that irisin enhanced the paracrine effects of BMSCs, consistent with a previous study (16).
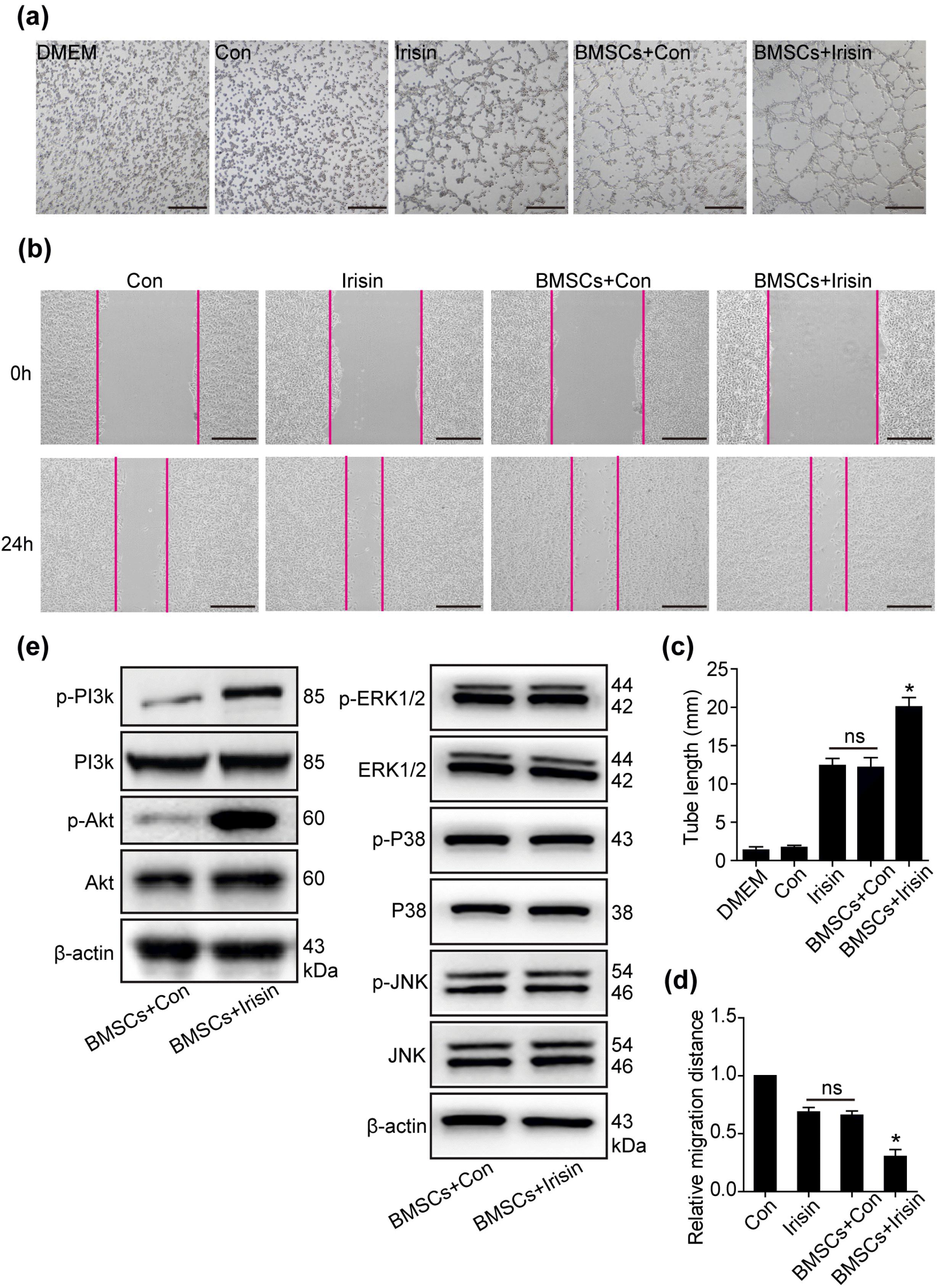
Fig. 3
Irisin promotes the paracrine efficacy of BMSCs through the PI3K/AKT pathway. (a) Conditioned media of equivalent cell numbers (1×106 cells) were collected for irisin-treated BMSCs and control groups. By employing HUVECs, tube forma-tion was assessed using the supernatant of BMSCs treated with irisin and control, irisin alone, and control alone. Scale bar, 50 μm. (b) The wound-healing assay was performed to demonstrate HUVEC migration using the above conditioned media, respectively. Scale bar, 243.2 μm. (c) The number of branch points was quantified by Image-Pro software and is shown in bar graphs (n=3 separate studies). (d) Quantification of HUVEC migration by Image-Pro software (n=3). (e) Western blot analyses of pPI3K, PI3K, p-AKT, AKT, p-ERK1/2, ERK1/2, p-P38, P38, p-JNK, and JNK pathway were detected in irisin-treated BMSCs and control groups. All data were measured as mean±standard error of the mean (SEM). *p<0.05. BMSCs, bone marrow mesenchymal stem cells; HUVECs, human umbilical vein endothelial cells.
![]()
In MI, the PI3k/Akt pathway is deemed the primary signal transduction pathway associated with cardioprotec-tion (17). In a cardiotoxicity model, irisin was found to be involved in cardiomyocyte apoptosis via Akt activation (6). Herein, we sought to monitor the level of PI3k/Akt in irisin-regulated BMSC paracrine signaling. In the present study, p-PI3k/PI3k and p-Akt/Akt were increased in irisin-mediated BMSCs when compared with the control group, with no differences in the p-ERK1/2/ERK1/2, p-P38/P38, and p-JNK/JNK pathways, revealing that PI3k/Akt was involved in irisin-regulated BMSC paracrine signaling (Fig. 3e). Furthermore, inhibition of the PI3k/Akt pathway (LY294002) was performed to suppress irisin-induced tube formation (Fig. 4a and 4b). In the wound-healing assay, the increased HUVEC migration, regulated by irisin-treated BMSCs, was blocked by LY294002 (Fig. 4c and 4d). However, a slight increased effect of tube formation and HUVEC migration by irisin despite LY294002 administration, suggesting that alternate signaling pathways are involved in irisin-regulated BMSC paracrine signaling (Fig. 4c and 4d). These data revealed that irisin promoted BMSC-mediated paracrine signaling via the PI3k/Akt pathway.
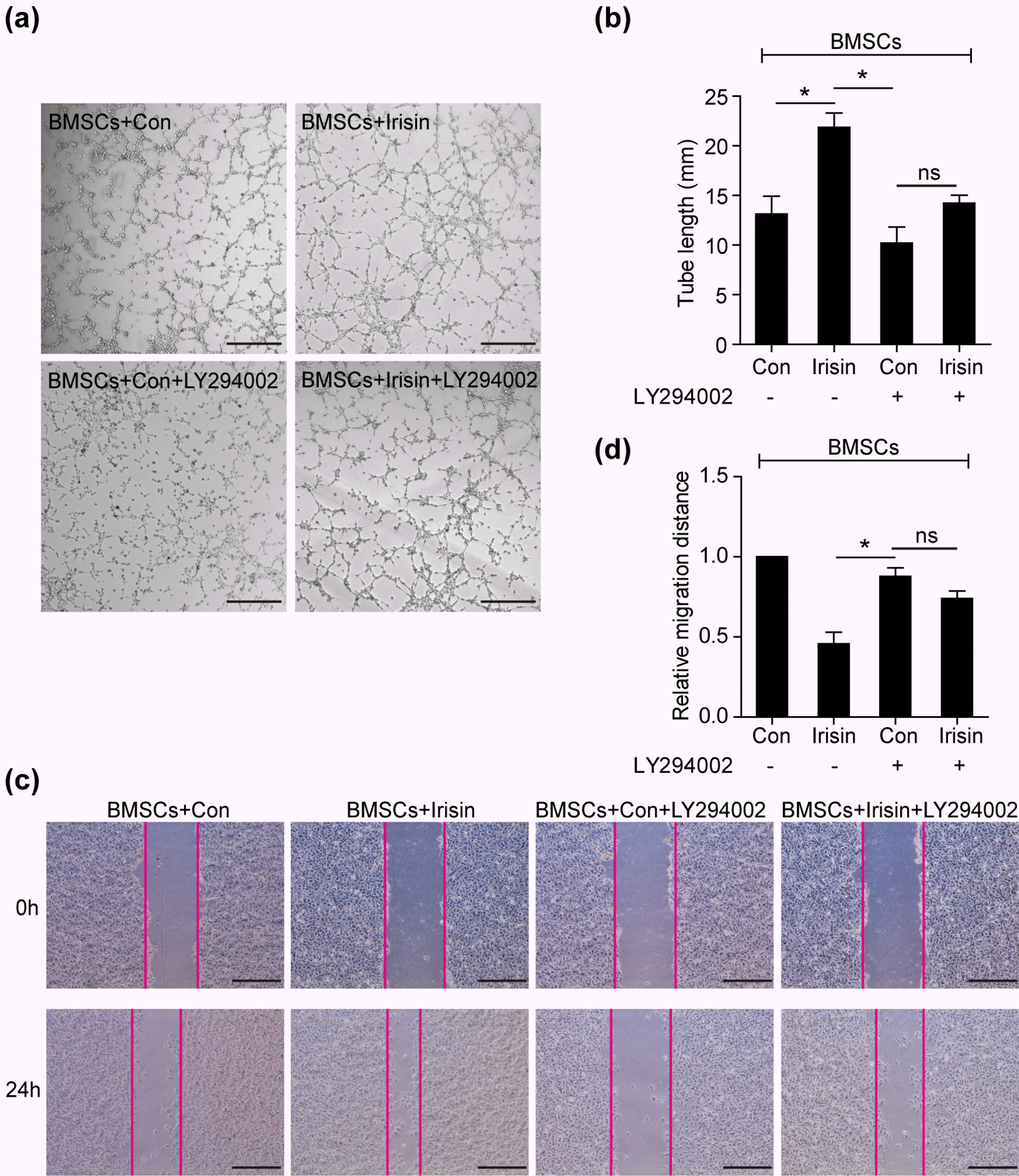
Fig. 4
The PI3K/AKT pathway is required to mediate the paracrine efficacy of BMSCs. (a) Representative images of tube formation were analyzed using HUVECs co-cultured with the supernatant of BMSCs treated with irisin, LY294002, irisin+LY294002, or control, respectively. Scale bar, 50 μm. (b) The number of branch points in the above groups was quantified by Image-Pro software and is shown in bar graphs (n=3). (c) The wound-healing assay assessed HUVEC migration using the above conditioned media, respec-tively. Scale bar, 243.2 μm. (d) Quantification of HUVEC migration (n=3) by Image-Pro software. Data are presented as mean±standard error of the mean (SEM). *p<0.05. BMSCs, bone marrow mesenchymal stem cells; HUVECs, human umbilical vein endothelial cells.
![]()
Discussion
In the present study, we observed that the irisin administration increased the angiogenesis of BMSCs in the border zone to protect cardiac function post-MI in vivo. Consistent with this phenomenon in vivo, irisin treatment promoted the paracrine efficacy of BMSCs in vitro. Mechanistically, we observed that irisin activated the PI3k/Akt signaling pathway, primarily improving the paracrine efficiency of BMSCs. Combined with these data, it was speculated that irisin could be a potential novel therapeutic agent for BMSC transplantation in tissue engineering and regenerative medicine for cardiovascular diseases.
Mesenchymal stem cells (MSCs) are a promising therapeutic option for immune-mediated diseases, cardiova-scular diseases, diabetes, and cancer (13, 18, 19). The paracrine effect of MSCs is reportedly considered the principal mechanism underlying MSC-based therapy, including exosomes and other paracrine factors such as basic fibroblast growth factor (bFGF) and vascular endothelial growth factor (VEGF) (13, 20). Based on the paracrine action, MSCs enhance angiogenesis via HIF-1α, miR-200b, and the leptin signaling pathway (17). Similarly, the paracrine function was observed in the present study, as revealed by the Matrigel assay.
In skeletal muscles, irisin is a myokine cleaved from FNDC5, especially after exercise. The role of irisin is multifunctional, with activities such as anti-inflammation, neuroprotection, bone formation, and metabolic regulation (21, 22). As a hormone secreted into circulation, irisin has been shown to afford protection against cardiovascular injury, reduce cardiac fibrosis, and improve cardiac function (23, 24). Moreover, irisin is considered crosstalk in muscle–adipose–bone–neuron connectivity mediated by paracrine or endocrine activity (25, 26). However, the functional role of irisin in mediating the paracrine effects of MSCs remains elusive. In the present study, we further demonstrated that irisin treatment enhanced the paracrine function of MSCs. Despite irisin functions discovered in several previous studies, our data revealed a new aspect of irisin-related paracrine effects in MSCs.
The therapeutic potential of BMSCs has been reported in several studies (27). However, the lower viability, inefficient homing, and migration remain major hurdles in the development of BMSC-mediated treatment strategies; therefore, numerous studies have attempted to improve the ability of BMSCs, including genetic modification and target tissue modification (28, 29). Although we failed to demonstrate the cardioprotection afforded by BMSCs, irisin showed a trend toward improvement when compared with DMEM. Meanwhile, we observed that irisin enhanced not only the angiogenesis of BMSCs but also their survival (Supplementary Fig. S1), which was consistent with a previous study demonstrating irisin-mediated improvements on BMSCs in an MI model (16).
The mechanism via which irisin exerts its paracrine function remains unclear. Mitogen-activated protein kinases (MAPKs), as a major signaling pathway of irisin, play a critical role in cellular mitosis, differentiation, metabolism, and apoptosis (5). However, we failed to determine the influence of irisin on the ERK1/2, P38, and JNK pathways in BMSCs. As irisin regulates neural differentiation and endothelial cell proliferation via the ERK1/2 pathway and mediates browning of white adipocytes via the P38 pathway (30), we speculated that irisin mainly activated these pathways during cell proliferation; however, we detected that irisin had no impact on BMSC proliferation as examined by the Cell Counting Kit 8 (CCK-8) assay (data not shown).
The regulation of the PI3k/Akt signaling pathway is involved in a large spectrum of cellular fates, including growth, proliferation, metabolism, and survival (31). Reportedly, improved MSC survival and paracrine signaling have been associated with the activation of the PI3k/Akt pathway in a mixture of moringin and cannabidiol-induced apoptosis and heat stress-induced skin injury models (32, 33). Moreover, inhibition of the PI3k/Akt pathway could trigger cancer stem cell death and alleviate ovarian cancer (34). However, irisin regulation via the PI3k/Akt pathway needs to be comprehensively investi-gated. Zhang et al. (35) have observed that irisin modulated apoptosis of pancreatic cancer cells by inactivating the PI3K/AKT pathway. Liu et al. (36) have reported that irisin inhibits doxorubicin-induced apoptosis in pancreatic cancer. Meanwhile, in liver cancer, irisin increased cell proliferation and invasion via activation of the PI3K/AKT pathway (37). In a mouse model of type 2 diabetes, Liu et al. (38) have demonstrated that irisin improves insulin resistance and insulin sensitivity via PI3K/AKT pathway activity. It has been suggested that irisin may bind to different cellular receptors to activate or inhibit the PI3K/AKT pathway in diverse cells. Therefore, our study determined the effect of irisin on the PI3K/AKT pathway following MSC transplantation in an MI mouse model. PI3K kinases are a family of enzymes that constitute three classes, Class I (p85/p110 and p101/p120), Class II (PI3KC2α, PI3KC2β, and PI3KC2γ), and Class III (VPS34), and p85 is the primary and canonical subunit of PI3K (39). We observed that the PI3K/AKT pathway was activated following irisin administration using the PI3K p85 antibody. However, irisin demonstrated marginal paracrine effects following administration of a PI3k/Akt pathway inhibitor, indicating other minor signaling pathways are involved in mediating the effects of irisin, such as PGC-1α and IGF-1 (40).
In conclusion, in the present study, we demonstrated that irisin promoted the angiogenesis and paracrine effects of BMSCs to improve cardiac function in MI model mice through the PI3k/Akt pathway. Accordingly, we propose a novel optimized MSC strategy for regenerative medicine.
Supplementary Materials
Supplementary data including one figure can be found with this article online at https://doi.org/10.15283/ijsc21005.
 XML Download
XML Download