

 PDF
PDF Citation
Citation Print
Print


Introduction
Diabetes mellitus (DM) is a severe chronic metabolic disease reflected by high glucose levels that induce oxidative stress. Globally, DM affects >400 million people, and cases are expected to increase by 50% by 2030 (1-3). Micro- and macrovascular traits are the main diabetic complications and are the key mortality risk factors for diabetic patients (4). Previous research has shown that adipose-derived stem cell (ADSC) transplantation therapy promotes diabetic wound closure (5). However, high glucose microenvironments in diabetic patients inhibit ADSC survival, resulting in poor treatment outcomes (6). Therefore, efficient mechanisms that enhance stem cell survival in these inclement microenvironments must be delineated.
Increasingly, studies have observed that hypoxia pretreatment promotes ADSC proliferation via increased vascular endothelial growth factor (VEGF) levels (7); however, the regulatory mechanisms for this effect remain unclear. A recent study observed that circular RNAs (circRNAs) actively participate in pathological mechanisms across different disorders. CircRNAs are more stable than linear RNAs and are highly expressed in some tissues; thus, they are considered a more promising potential biomarker than linear RNAs (8). Previously, circRNAs were reported to function as miRNA ‘sponges’ that naturally and competitively sequester and suppress miRNA activity (9-11). However, how circRNAs function in ADSC-mediated diabetic wound healing remains unclear. Therefore, we employed high-throughput sequencing, RT-qPCR and a diabetic wound mouse model to study circRNAs in ADSC acceleration of diabetic wound healing.
Go to : 
Materials and Methods
Ethics statement
The Animal Care and Use Committee of Peking Union Medical College Hospital Hospital approved the investigation protocol (XHDW-2020-01). We performed all postoperative animal care and surgical interventions according to the NIH guide for Care and Use of Laboratory Animals.
ADSC isolation and identification
ADSCs were isolated from mouse adipose tissue as previously described (12, 13). We observed no uninduced differentiation during our cell culture expansion. We induced osteogenic differentiation in 3-week ADSC cultures in Dulbecco’s modified Eagle’s medium (DMEM) (Invitrogen, Carlsbad, CA, USA), supplemented with 10% fetal bovine serum (FBS) (Invitrogen, Carlsbad, CA, USA), 0.1 μM dexamethasone, 50 μM ascorbate-2-phosphate and 10 mM β-glycerophosphate. We induced adipogenic differentiation by culturing ADSCs for two weeks in DMEM supplemented with 10% FBS, 10 μM insulin, 0.5 mM isobutylmethylxanthine, 200 μM indomethacin and 1 μM dexamethasone. ADSC osteogenic or adipogenic differentiation was investigated using alizarin red (Invitrogen, Carlsbad, CA, USA) and oil-red O (Invitrogen, Carlsbad, CA, USA) staining. We grew normoxic ADSCs cultures in 95% air (20% O2) and 5% CO2. For hypoxic pretreatment, ADSCs were cultures in 93% N2, 2% O2 and 5% CO2.
Strand-specific and high-throughput RNA-Seq library construction
Total RNA from ADSCs and hypoxic-pretreated ADSCs was isolated using TRIzol reagent (Invitrogen, Carlsbad, CA, USA). Approximately 3 μg total RNA per sample was prepared using a VAHTS Total RNA-seq (H/M/R) Library Prep Kit from Illumina (Vazyme Biotech Co., Ltd, Nanjing, China). This kit isolated ribosomal RNA and removed the remaining RNAs, including non-coding RNA and mRNA. We then performed an RNA purification step using RNase R (Epicenter, 40 U at 37℃ for 3 h), followed by TRIzol. An RNA-seq library was established using a KAPA Strand RNA-Seq Library Prep Kit (Roche, Basel, Switzerland) and expose them in order for extensive codifying with Illumina HiSeq 4000 from Aksomics, Inc. (Shanghai, China).
Diabetic wound induction
We used BALB/C mice to induce a diabetic state via a single intraperitoneal injection of 60 mg/kg streptozotocin (STZ, Sigma, USA) dissolved in 0.1 M citrate buffer (pH 4.5). On day 7 after STZ administration, we validated the presence of diabetes by assessing fasting blood glucose levels in blood samples from tail veins. A mouse with a fasting blood glucose level of >250 mg/dl was considered diabetic. Animals were maintained for one month and were used for subsequent analyses of posterior blood glucose stabilization. Following diabetes confirmation, we anesthetized mice by intramuscular injection of 30 mg/kg sodium pentobarbital. Once anesthetized, the hair was shaved from the dorsal leg area and the region sterilized using povidone iodine solution. A sterile biopsy punch was used to generate a 4-mm full-thickness excisional wound. After this procedure, we randomly allocated mice to subcutaneous injection with 100 μl PBS containing 1×105 ADSCs, or an equal volume of PBS at four sites near the wound (25 μl/site). We euthanized mice after three weeks and harvested skin samples for histopathological analyses.
CD31 immunohistochemistry
We fixed, processed, and blocked skin sections for immunofluorescence staining. Sections were incubated overnight at 4℃ with an anti-rabbit CD31 antibody, followed by incubation with biotinlyated goat anti-mice IgG antibody (Vector laboratories). Following secondary antibody incubation, we incubated sections in avidin:biotinylated enzyme complex (ABC; Vector Laboratories) for 30 min. We then treated sections with 3,3’-diaminobenzidine substrate (Vector Laboratories), and the section produced a brown stain. We mounted slides using Fluka Eukitt quick-hardening mounting medium (Sigma Aldrich). Negative controls contained no primary antibodies. Images were captured using a Carl Zeiss MIRAX MIDI via a Plan-Apochromat 20×/0.8 NA objective lens and Marlin F146.C camera, and images were processed in MIRAX Viewer Version 1.11.49.0 software (Carl Zeiss Microimaging GmbH, Oberkochen, Germany).
RNA overexpression or interference
We induced miR-18a-5p overexpression using miR-18a-5p mimics. We purchased a circ-Astn1 overexpression vector and HIF-1α silencing vector from RiboBio (Guangzhou, China). We performed all cell transfections using Lipofectamine 2000 (Thermo Fisher Scientific), following a previous method (14).
Western blot assay
Skin tissues were lysed, and lysates were centrifuged at 12,000 rpm at 4℃ following addition of a protease inhibi-tor. The protein concentration was determined with a Pierce bicinchoninic acid assay (BCA) kit (Thermo Fisher). Proteins was separated by 10% SDS-PAGE and transferred to PVDF membranes. The primary antibodies used to assay protein expression were VEGF (1:6S00), HIF-1α (1:600) (all Santa Cruz Biotechnology, Dallas, TX, USA), Anti-GAPDH (1:1,000, Sigma-Aldrich). Horse-radish peroxidase-conjugated secondary antibody (1:1,000, Abcam, USA). An ECL chemiluminescent kit (Millipore, Burlington, MA, USA) was used to read the bands.
Quantitative real-time polymerase chain reaction (RT-qPCR)
Total RNA was isolated from wound tissues and cells using a TRIzol reagent kit. We synthesized and amplified cDNA using a TaqMan miRNA Reverse Transcription Kit. RT-qPCR reactions were performed using a TaqMan Human miRNA Assay Kit, using the 2−ΔΔCT method to detect expression fold changes. We used U6 and GAPDH genes as internal references. Primers were: circ-Gcap14, F: 5’-GCAGCTATGAGTCCTCTG-3’, R, 5’-CTTCCATGGGC TATAAGGTG-3’; HIF-1α, F: 5’-AGAGTCAAGCCCAGAG TCAC-3’, R, 5’-TGGGACTGTTAGGCTCAGGT-3’; VEGF, F: 5’-GCACCCATGGCAGAAGGAGGAG-3’, R, 5’-GTGC TGACGC-TAACTGACC-3’; miR-18a-5p, F: 5’-CAGTAAA GGTAAGGAGAGCTCAATCTG-3’, R: 5’-CATACAACCA CTAAGCTAAAGAATAATCTGA-3; U6, F: 5’-AGTAAGC CCTTGCTGTCAGTG-3’, R: 5’-CCTGGGTCTGATAATG CTGGG-3’; GAPDH: F: 5’-GTCTCCTCTGACTTCAACA GCG-3’, R: 5’-ACCACCCTGTTGCTGTAGCCAA-3’ (Gene Pharma, Shanghai, China).
Luciferase reporter assay
We cloned wild-type (wt) and 3’-UTR mutant (mut) HIF-1α, and wt and mut circ-Gcap14 into pMIR firefly luciferase-expressing vectors. We co-transfected vectors into 70% confluence HEK293T cells. We used 500 ng pMIR-HIF-1α-wt/pMIR-HIF-1α-Mut or pMIR-circ-Gcap14-wt/pMIR-circ-Gcap14-mut combined with 50 nM miR-18a-5p mimics. We assayed luciferase activity using the Dual-Luciferase Reporter System (Promega, Madison, WI, USA) and performed five independent assays.
Statistical analyses
We expressed continuous parameters using the mean± standard deviation (SD), and used one-way variance analysis (ANOVA) to compare data in GraphPad Prism 5.1. A p value≤0.05 indicated statistical significance.
Go to :
Results
The hypoxic pretreatment increases the therapeutic effect of ADSCs to accelerates diabetic wound closure
ADSCs can effectively repair diabetic wounds (15). We observed that ADSCs possessed classical cobblestone-like morphology (Fig. 1A). Immunofluorescence staining was positive for the mesenchymal cell markers CD44, CD90, CD29 and CD105, and negative for the endothelial markers CD31 and von Willebrand Factor (vWF) (Fig. 1B∼G). Alizarin red and oil red O staining also confirmed that ADSCs exhibited osteoblast and adipocyte differentiation abilities (Fig. 1H and 1I).
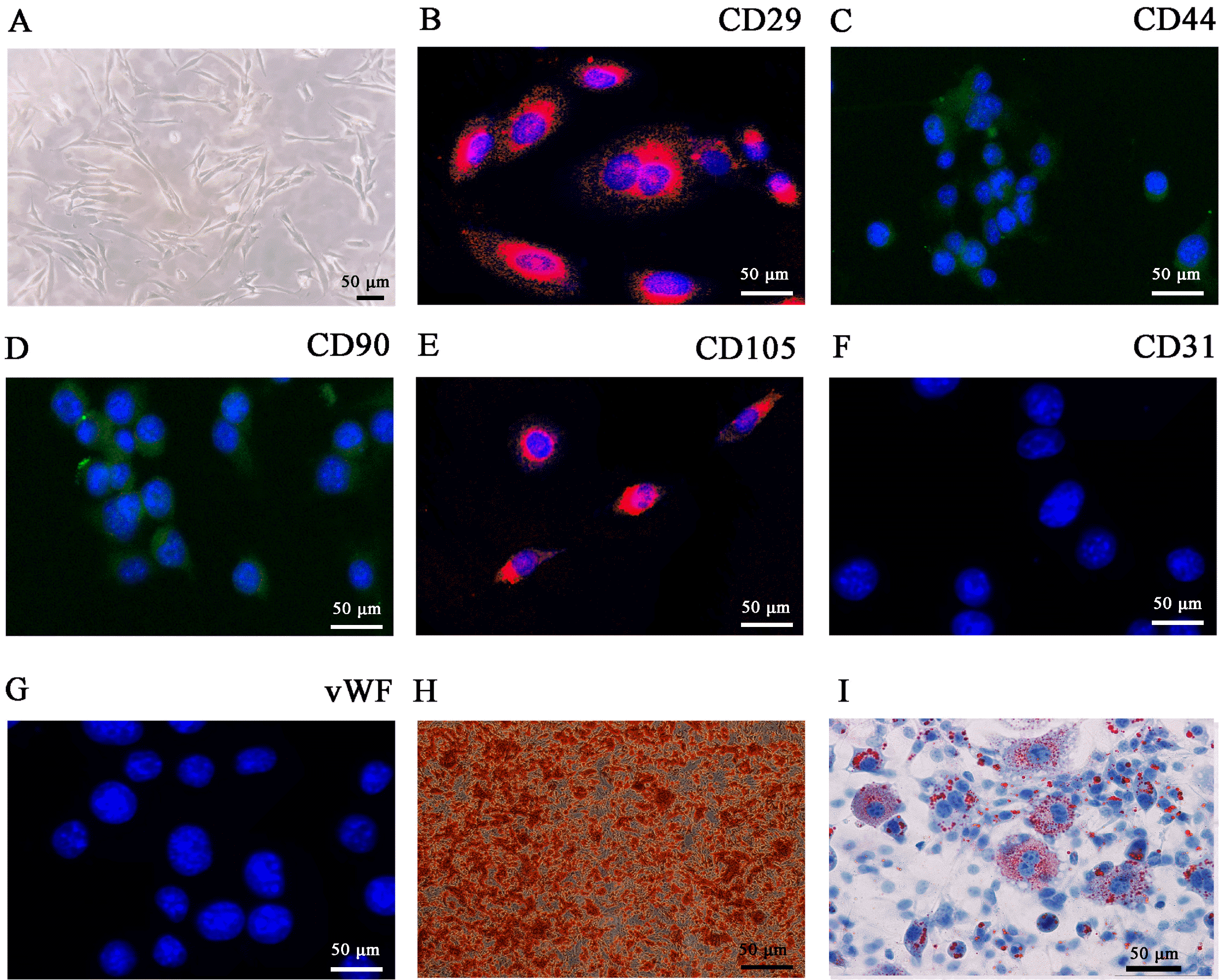 | Fig. 1Characterization of adipose-derived mesenchymal stem cells (ADSCs). (A) ADSCs show a typical cobblestone-like morphology. (B∼G) Immunofluorescence staining of cell surface markers. Antibodies were labeled with either fluorescein isothiocyanate (FITC, green) or phycoerythrin (PE, red). CD29, CD44, CD90, and CD105 staining was positive, whereas CD31 and von Wille-brand Factor (vWF) staining was negative. (H, I) Differentiation potential of ADSCs by oil red O (H) and alizarin red staining (I). Scale bar, 50 μm.
|
A previous study indicated that the hypoxic pretreatment of ADSCs improved therapeutic efficacy (7). To clarify if hypoxic pretreatment of ADSCs could indeed accelerate diabetic wound closure, mice were injected with STZ for one week to construct a diabetic mouse model. We anesthetized mice with sodium pentobarbital (0.5 mg/g), and made a 10 mm full-thickness round excisional skin wound on every mouse instep, with a sterile 10-mm punch biopsy tool. Following received control ADSCs or hypoxia-pretreated ADSCs, diabetic wound closure was detected 0, 7 and 14 day after surgery (Fig. 2A). Our results showed that hypoxic pretreatment of ADSCs accelerated diabetic wound closure (Fig. 2B and 2C). Immunohistochemical CD31 staining showed that this pretreatment promoted angiogenesis (Fig. 2D and 2E). RT-qPCR and western blot analyses showed that hypoxic pretreatment of ADSCs significantly increased HIF-1α and VEGF expression (Fig. 2F and 2G). These data suggested that hypoxic ADSC pretreatment accelerated diabetic wound closure, and increased angiogenic growth factor expression in our mouse model.
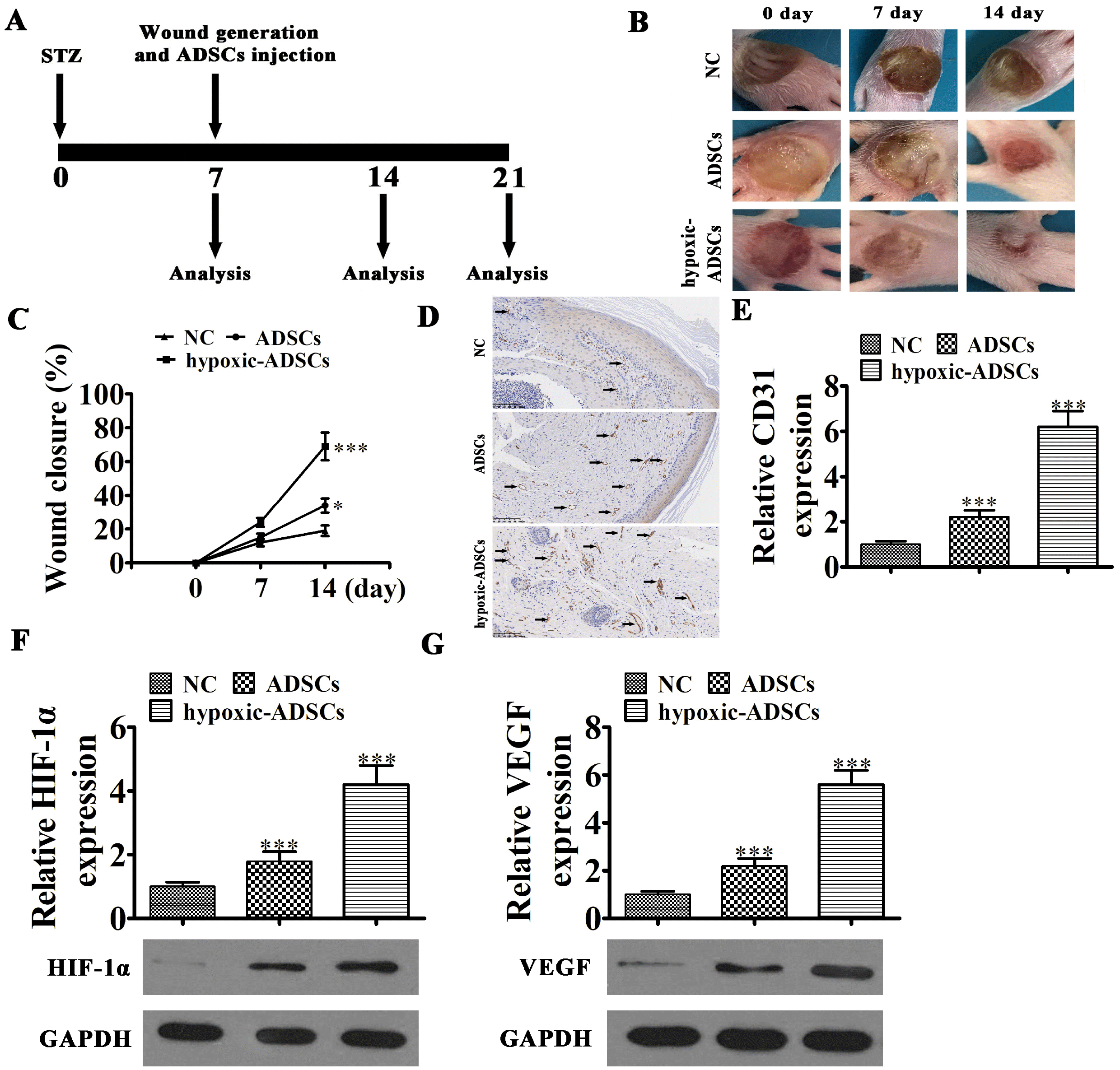 | Fig. 2Wound healing effects of hypoxia-pretreated ADSCs in mice. (A) Schematic of in vivo procedures. (B, C) Wound closure rates were quantified at indicated times after wound generation and ADSC transplantation. Data are presented as the mean±SD. *p<0.05, ***p<0.001 vs. NC group. (D, E) CD31 immunohistochemical staining detection the angiogenesis as black arrow show. Data are presented as mean±SD. ***p<0.001 vs. NC group. (F, G) RT-qPCR and western blot detection show the expression HIF-1α and VEGF in mRNA and protein level from tissue surrounding wounds in the diabetic mouse model. Data are presented as the mean±SD. ***p<0.001 vs. NC group.
|
Circ-Gcap14 played an important role in hypoxic-ADSC accelerated diabetic wound closure, and enhanced angiogenic expression in our mouse model
CircRNAs have important functions in microenviron-mental regulation (16). In our study, high-throughput sequencing was used to explore if circRNAs play a role in ADSC-accelerated diabetic wound closure. Our data revealed that ADSC hypoxic pretreatment resulted in several differentially expressed circRNAs (Fig. 3A). RT-qPCR analysis showed that circ-Gcap14 expression was significantly increased in hypoxic-ADSCs when compared with non-hypoxic cells (Fig. 3B).
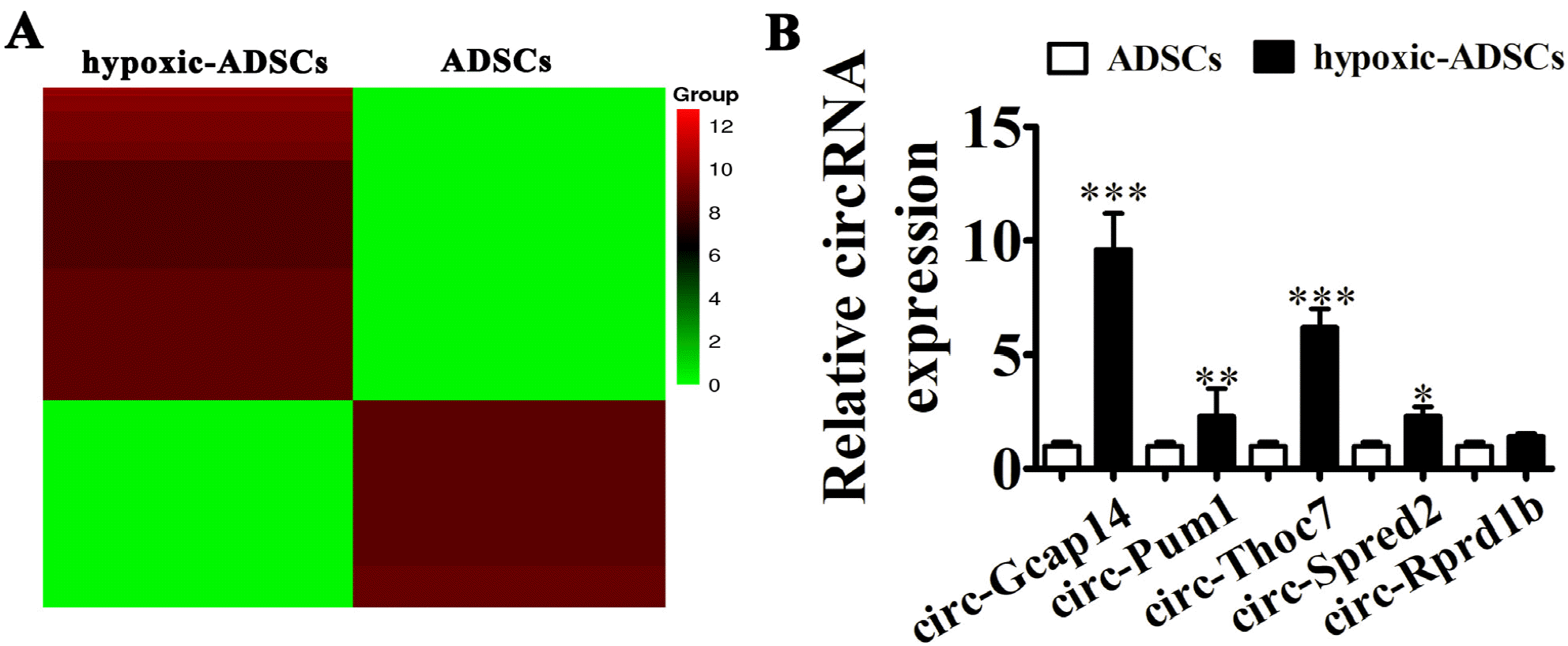 | Fig. 3Circ-Gcap14 is upregulated in hypoxia-pretreated ADSCs (hypoxic-ADSCs). (A) Heat map of upregulated and downregulated circRNAs with a ≥1.5-fold difference between hypoxic- and wild-type ADSCs. (B) RT-qPCR shows circRNA expression in both hypoxic- and wild-type ADSCs. Data are presented as the mean±SD. **p<0.01, ***p<0.001 vs. ADSCs group.
|
We then constructed a circ-Gcap14 overexpression and silencing vector, and transfected both into ADSCs. As expected, our data showed that circ-Gcap14 expression was significantly increased after transfection with the circ-Gcap14 overexpression vector, but was significantly silenced upon circ-Gcap14 downregulation (Fig. 4A). After ADSC pretreatment with hypoxia, circ-Gcap14 overexpression significantly accelerated diabetic wound closure, but circ-Gcap14 silencing decreased diabetic wound closure (Fig. 4B and 4C). Immunohistochemical staining for CD31 showed that circ-Gcap14 overexpression promoted angiogenesis, but circ-Gcap14 silencing decreased it (Fig. 4D and 4E). RT-qPCR and western blot analysis also showed that circ-Gcap14 overexpression increased VEGF and HIF-1α expression, while circ-Gcap14 silencing decreased this expression (Fig. 4F and 4G). These data suggested that circ-Gcap14 functions in hypoxic-ADSC acceleration of diabetic wound closure, and enhances angiogenic growth factor expression in our mouse model.
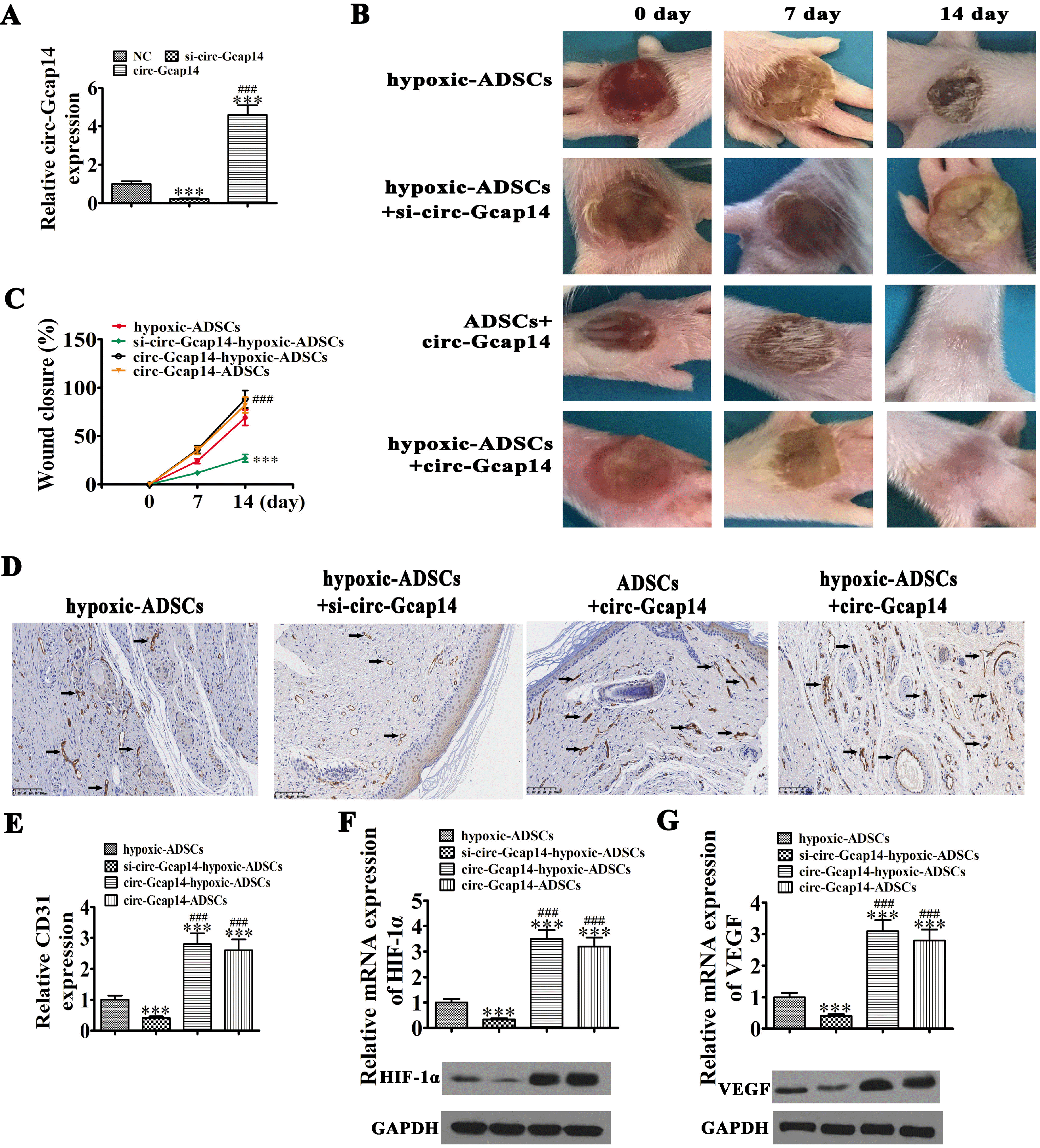 | Fig. 4Circ-Gcap14 plays an important role in ADSC mediated diabetic wound closure. (A) RT-qPCR shows circ-Gcap14 expression in ADSCs upon circ-Gcap14 downregulation or overexpression/silencing. Data are presented as the mean±SD. ***p<0.001 vs. NC. ###p<0.001 vs. si-circ-Gcap14. (B, C) The rate of wound closure was quantified at indicated times after ADSC transplantation. Data are presented as the mean±SD. ***p<0.001 vs. hypoxic-ADSC. ###p<0.001 vs. si-circ-Gcap14-hypoxic-ADSC. (D, E) CD31 immunohistochemical staining detection the angiogenesis as black arrow show. Data are presented as the mean±SD. ***p<0.001 vs. hypoxic-ADSC. ###p<0.001 vs. si-circ-Gcap14-hypoxic-ADSC. (F, G) RT-qPCR and western blot detection shows the expression of HIF-1α and VEGF in both mRNA and protein level from tissue surrounding wounds. Data are presented as the mean±SD. ***p< 0.001 vs. hypoxic-ADSC. ###p<0.001 vs. si-circ-Gcap14-hypoxic-ADSC.
|
The expression of circ-Gcap14 increased HIF-1α level by adsorption miR-18a-5p
Bioinformatics data showed that circ-Gcap14 regulates HIF-1α expression via miR-18a-5p inhibition. HIF-1α is involved in angiogenesis by promoting VEGF expression (17, 18). To validate interactions between circ-Gcap14, miR-18a-5p and VEGF, we created a suite of luciferase reporter (LR) vectors. MiR-18a-5p binding sites on circ-Gcap14, as well as sites with point mutations were added to prevent binding (Fig. 5A). By performing the luciferase assay in mut or wt circ-Gcap14 transfected HEK293T cells, we showed that miR-18a-5p suppressed circ-Gcap14 activity (Fig. 5B).
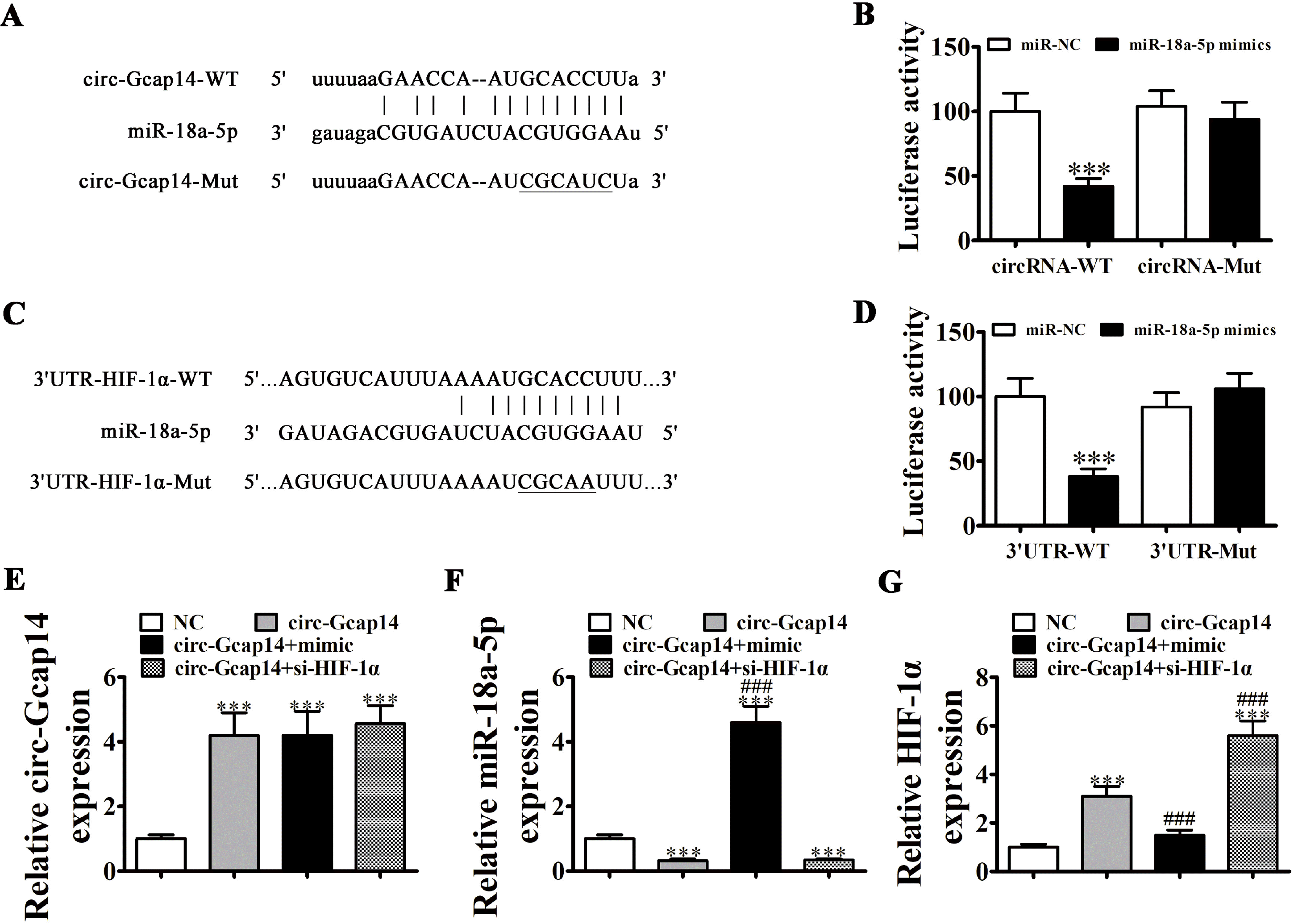 | Fig. 5miR-18a-5p and HIF-1α are downstream targets of circ-Gcap14. (A) Predicted binding sites for miR-18a-5p in circ-Gcap14. The mutated circ-Gcap14 is also shown. (B) Relative luciferase activity at 48 h post-transfection of HEK293T cells with miR-18a-5p mimics/NC or circ-Gcap14 wild-type/mutant. Data are presented as the mean±SD. ***p<0.001. (C) Binding site prediction of miR-18a-5p in the HIF-1α 3’UTR. The mutant version of the 3’-UTR-HIF-1α is shown. (D) Relative luciferase activity at 48 h post-transfection of HEK293T cells with miR-18a-5p mimic/NC or 3’UTR-HIF-1α wild-type/mutant. Data are presented as the mean±SD. ***p<0.001. (E∼G) RT-qPCR shows circ-Gcap14 (E), miR-18a-5p (F) and HIF-1α expression (G). Data are presented as the mean±SD. ***p<0.001 vs. NC group. ###p<0.001 vs. circ-Gcap14.
|
We next created the LR vector. Candidate miR-18a-5p binding sites regarding HIF-1α 3’-UTR and those with point mutations inserted to prevent binding were constructed (Fig. 5C). We transfected HEK293T cells with mut or wt HIF-1α 3’-UTR and assayed luciferase activity, demonstrating that wt miR-18a-5p suppressed HIF-1α activity (Fig. 5D). RT-qPCR analyses also showed that circ-Gcap14 overexpression promoted circ-Gcap14 and HIF-1α expression, but decreased miR-18a-5p expression (Fig. 5E), suggesting that miR-18a-5p and HIF-1α were downstream targets of circ-Gcap14. MiR-18a-5p upregulation suppressed HIF-1α expression, but HIF-1α overexpression did not reverse miR-18a-5p expression, suggesting that HIF-1α was a miR-18a-5p downstream target (Fig. 5F and 5G). These data showed that both miR-18a-5p and HIF-1α were downstream circ-Gcap14 targets.
Go to :
Discussion
In DM patients, acute skin wounds resulting from trauma usually turn to chronic non-healing wounds (19). Several methods have been tried to accelerate the diabetic wound healing process, but optimized therapeutic strategies are lacking (20). Previous investigations have suggested that ADSC transplantation therapy accelerates diabetic wound closure (13, 15). Thus, in this study, we observed that hypoxic pretreatment of ADSCs exerted greater acceleration effects on diabetic wound closure than non-hypoxic ADSCs. Under hypoxic pretreatment conditions, high-throughput sequencing identified circ-Gcap14 expression as critical in the process; downregulated/si-lenced circ-Gcap14 decreased ADSC therapeutic effects, whereas circ-Gcap14 overexpression increased these therapeutic effects, suggesting that circ-Gcap14 plays a role in the ADSC acceleration of diabetic wound closure.
Bioinformatics and luciferase reporter analyses found that HIF-1α and miR-18a-5p were circ-Gcap14 downstream targets. The overexpression of circ-Gcap14 promoted HIF-1α and miR-18a-5p downregulation. Previous studies have shown that VEGF is the main HIF‐1α downstream effector, inducing cell proliferation, migration and tube formation in endothelial cells (21-23). The HIF‐1α/VEGF signaling pathway is a critical target for angiogenic-related disease therapies, e.g. diabetic ulcers (24, 25). Our study also confirmed that circ-Gcap14 has an important role in hypoxic-ADSC acceleration of diabetic wound closure, and enhances VEGF expression.
In conclusion, our study indicates that the hypoxic pretreatment of ADSCs enhances wound healing in a diabetic mouse model via induction of the circ-Gcap14/miR-18a-5p/HIF‐1α/VEGF axis. Our study ascertained the therapeutic effects of circ-Gcap14 in an STZ-induced diabetic wound healing mouse model.
Go to :
 XML Download
XML Download