

 PDF
PDF Citation
Citation Print
Print


Introduction
Cardiovascular diseases remain one of the significant mortalities worldwide. Ischemic heart disease (IHD) is still one of the five major factors leading to premature death in China (1). Millions of patients suffer from heart failure or new or recurrent myocardial infarction (MI) every year. IHD results in CMs’ gradual death through apoptosis and/or necrosis and causes fibrotic replacement of the dead CMs. Subsequently, fibroblasts proliferate and migrate to the impaired area and remodel the myocardium through extracellular matrix deposition, resulting in increased tissue stiffness and decreased contraction. Excessive myocardial fibrosis is a vital driver in the course of various heart diseases and heart failure.
The minimal regenerative capacity of the human heart has extensively promoted new techniques for producing CMs in vivo and in vitro. With the ground-breaking scientific discovery of human embryonic stem cells (hESCs) (2) and human-induced pluripotent stem cells (hiPSC) (3, 4), researchers focus on developing reliable methods that affect the induction of stem cells differentiation into the cardiovascular lineage in recent decades. According to the enormous data, the above two forms of human pluripotent stem cells (hPSC) can produce numerous contractile CMs (5-7).
So far, although the cardiomyocytes generated by the hPSC (hPSC-CMs) display lots of resemblance with human primary CMs, hPSC-CMs have shown immature developmental status representing embryonic or fetal stages CMs. These hPSC-CMs exhibit spontaneous contraction, fetal ion channels, fetal electrophysiological properties (8), and fetal type gene expression patterns. Many recent studies have sought approaches to overcome this developmen-tal hindrance and further improve hPSC-CMs maturation to closer to native adult cardiac tissue by long-term culture, electrical and biomechanical stimulation methods, recently reviewed (9, 10). Furthermore, some investigators also develop an advanced three-dimensional (3D) cell culture system (that is, co-culture of non-CMs and extra-cellular matrix) to recapitulate in vivo environment (11, 12). Despite extensive efforts, we are still far from achieving PSC-derived CMs characteristics comparable to native or mature CMs in vitro (Fig. 1).
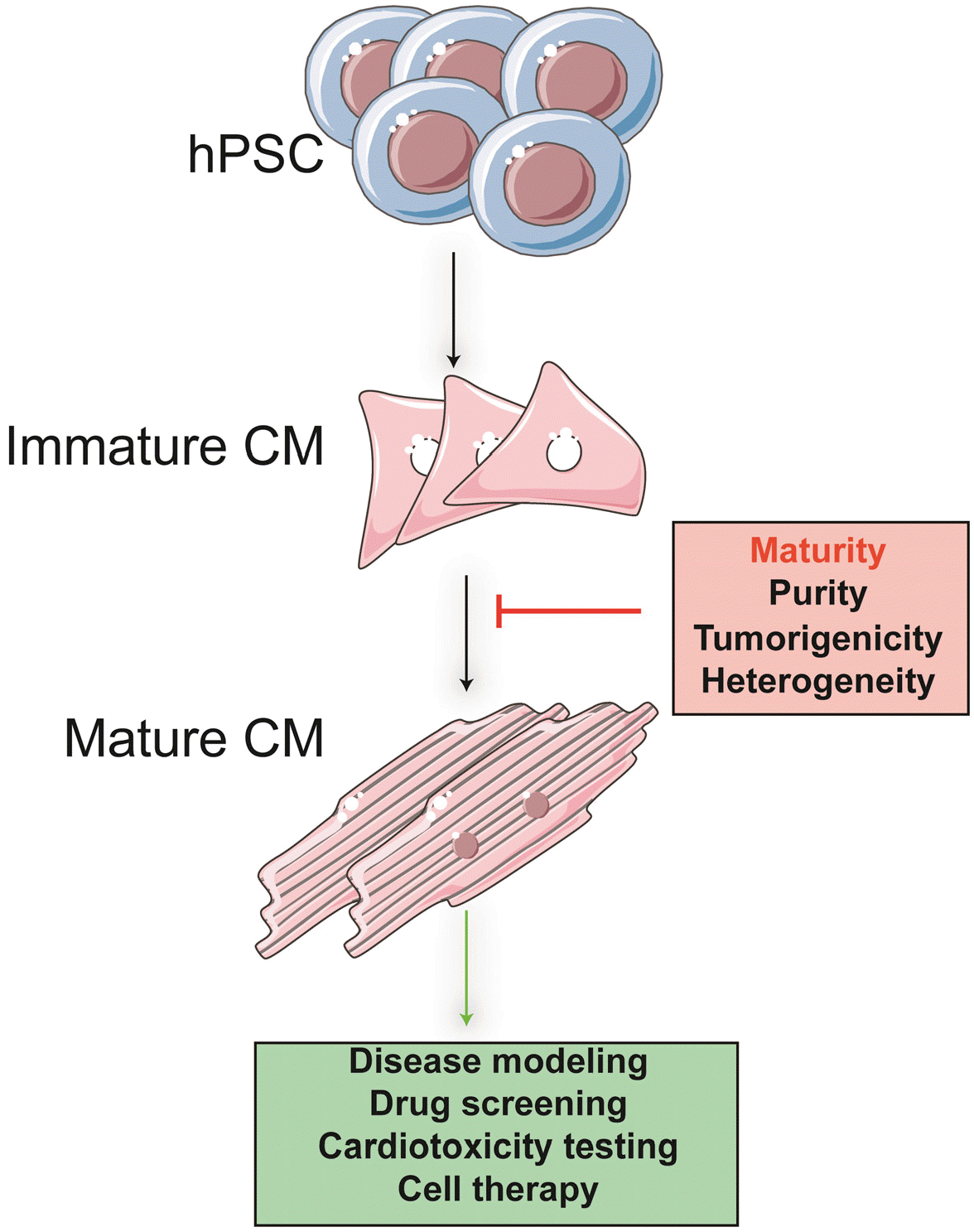
Thus, an in-depth understanding and identification of the molecular mechanism of cardiac maturation is necessary, which may help us better improve the maturation of hPSC-CMs, thereby achieving translational medicine applications.
Go to : 
Hallmarks of Cardiac Maturation
Major characteristics of CMs maturation are summarized in Table 1.
Table 1
Hallmarks of cardiac maturation
| Characteristics | Functional assessments | Key references | |
|---|---|---|---|
| Structure | Myofibrils massive expansion | Sarcomere length | (14-17, 20, 23, 26) |
| More clear banding | Fractional shortening | ||
| Improved sarcomere filament alignment | Contraction velocity | ||
| M-line more clear | |||
| Sarcomeric isoform switching | |||
| Metabolism | Increased number and size of mitochondria | Oxygen consumption rate | (47-50, 52, 53, 56) |
| Densely organized cristae | Mitochondrial membrane polarization | ||
| Mitochondrial membrane polarization | |||
| Glycolytic genes downregulation | |||
| Calcium handling & electrophysiology | More negative resting membrane potential | Calcium transient | (66-72, 76, 79) |
| Longer plateau phase of action potential | Peak height | ||
| Faster upstroke velocity | Departure velocity | ||
| T-tubulea formation | Action potential | ||
| Low automaticity | |||
| Cardiomyocyte proliferation | Decreased proliferation rate | Brdub | (84-89, 91-95, 97) |
| Repressed cell cycle regulators | Ki67 | ||
| Maturational hypertrophy | pH3c | ||
| Polyploidization |
![]()
Go to :
Sarcomere
The highly ordered tissue of striated muscle is the premise of rapid and unidirectional development of force and movement in the process of cardiac and skeletal muscle contraction (13-15). Myofibril is a 1-μm diameter, long cylindrical structure in striated muscle. A bunch of myofibrils provides the contractile function to skeletal muscle cells. Myofibrils are comprised of thick filaments and thin filaments. The components of thick filaments are myosins, and the main components of thin filaments are actins, supplemented by tropomyosins and troponins.
Under the electron microscope, the whole length of each myofibril presents regular bright and dark bands. The light and dark zones contain thinner and parallel filaments called myofilaments. The light band is isotropic, so it is called band I. The myofilament is thinner with a diameter of about 5 nm, which is called the thin myofilament. Part of the myofilament is located in the bright band, and the other part is in the dark band, inserted between the thick myofilaments. The dark band is also called band A. The myofilaments in the dark zone are larger with a diameter of about 15 nm, so they are called thick myofilaments. There is a bright narrow band in the center of the dark band called the band H; there is a thin film in the middle of the H band called the M membrane. There is also a thin film in the center of the bright band, called Z-membrane, or Z-line; The area of myofibril between two adjacent Z lines is called sarcomere, which is the basic unit of muscle contraction and relaxation.
In the process of myofilament gliding, when the intracellular Ca2+ is increased due to the excitation of CMs, Ca2+ binds to troponin on the filaments, which changes its configuration, thus pulling the troponin to roll and shift, exposing its covered binding sites. Meanwhile, Myosin ATPase on the cross-bridge can catalyze the decomposition of ATP, and the release of energy drives muscle contraction. When intracellular Ca2+ concentration is reduced, troponin dissociates from Ca2+ and returns to its resting position. The cross-bridge cannot contact the thin muscle filament, which makes the muscle enter the relaxation process.
Research has shown hESC-CMs and hiPSC-CMs between 20 and 40 days manifest poorly arranged contractile apparatus, exhibiting stunted myofibril density and orientation and fluctuating Z-disc alignment (15). However, substantial improvements in the myofibrillar density, alignment, and morphology are observed over prolonged in vitro culture, as shown in late period hESC-CMs and hiPSC-CMs (80∼120 days). An integral element of myofibril maturation is myofibrillar proteins isoform switching, such as titin, myosin heavy chain, α-actin, and the troponin complex. Due to extensive alternative splicing, the N2BA isoform of TTN is expressed in fetal CMs and hPSC-CMs, whereas the stiffer N2B isoform becomes restricted to adult CMs (16).
Another well-known sarcomere component is the myosin heavy chain, and it is expressed under two distinct isoforms in rodents, among which MYH7 is the predominant fetal isoform during fetal development, while MYH6 is the adult isoform in the adult hearts (17). On the contrary, fetal human CMs and hPSC-CMs initially express MYH6, while in adult CMs, MYH7 replaces it (18, 19). Likewise, slow skeletal muscle isoform (TNNI1) is predominantly expressed in fetal CMs, then switches to cardiac troponin I (TNNI3) as CMs mature (20-22). Studies have shown MLC-2A transcript expression is observed in both atria and ventricle during heart development in humans. In contrast to the mouse, it does not become chamber restricted; instead, it is widely expressed throughout heart development until adulthood. MLC-2V is identified as a maturation marker, which is not detected in the aria and slow-conducting tissues (23, 24). It is also reported that hiPSC-CMs present either one of these myosin light chain isoforms or both, to a distinct degree in the cultures (25). The isoform of myomesin (EH-myomesin) peaks in fetal CMs, but the isoform of myomesin lacking the EH domain can be detected in mature CMs (26). SMA is abundantly expressed in early embryonic phases in rodents and becomes undetectable in the adult heart (27, 28). hESC-CMs have been confirmed to express SMA in vitro but less in human fetal CMs (28). Desmin protein is the muscle-specific intermediate filament critical for maintaining sarcomeres (29) and is observed only in a fraction of hESC-CMs (25). By contrast, the typeⅢ intermediate filament protein vimentin is observed in fetal CMs and all hESC-CMs in the culture but not in adult rat CMs (25).
Contractile force, the critical feature of sarcomere contraction, results from fine regulatory interaction among electrical activation, calcium treatment, and myofilament activation. However, it is one of the least studied parameters, although the methods of contractile forces of CMs have been greatly developed. To the best of our knowledge, contractile force assays include magnetic beads (30), atomic force microscopy (AFM) (31, 32), micropost arrays (MPAs) (33-36), traction force microscopy (TFM) (37), optical edge detection (38), flexible cantilevers (39, 40), and strain gauges (41). Out of these techniques, MPAs and cantilevers have been used successfully to measure forces and assess cell maturation from CMs (33, 34, 36, 42). An early study reported peak isometric twitch tension was 44.0±11.7 mN/mm2 in human myocardium, peak twitch tension was 56.4+4.4 mN/mm2 in rat myocardium, 46.1± 2.6 mN/mm2 for rabbit myocardium (43). hPSC-CMs in a 3D collagen matrix with uniaxial mechanical load further increase the active force. However, this construct generates around 0.08 mN/mm2, about 550 times less than the adult human myocardium (44). Similarly, engineered heart tissue (EHT) consisting of neonatal rat CMs with collagen I and matrix factors revealed a high ratio of twitch (0.4 to 0.8 mN/mm2) to resting tension (0.1 to 0.3 mN/mm2) (45). Another group quantified the contraction force of hiPSC-CMs and hESC-CMs by using atomic force microscopy (AFM); they observed these two different sources of CMs contract with the comparable mechanical properties with contraction forces of 0.49±0.45 nN and 0.23±0.11 nN, respectively (32). In addition, individual CMs (which were plated onto polyacrylamide gels with an elastic modulus of 4 kPa and surfaces functionalized with chemically cross-linked gelatin) were detected using a method of dynamic traction force microscopy (46). Selected hESC-derived CMs contracted with an average axial force of 139±29 nN and a total force of 144±33 nN. Notably, these values did not change obviously, even if the culture time was prolonged to 90 days. In contrast, NRVCs contracted with an average axial force of 202±47 nN and a total force of 222±54 nN by employing the same method (46).
Go to :
Metabolism
Fetal nutrition relies predominantly on the placenta’s functional ability to intake glucose and lactic acid. During postnatal development, the transition to maternal milk nutrition raises the lipid diet content, and the increase of oxygen concentrations in circulating blood leads to metabolic reprogramming (47). Subsequently, roughly 50% of the total ATP is derived primarily from anaerobic glycolysis in the fetal heart. In contrast, by postnatal day (P) 7, β-oxidation of fatty acids is a major contributor to energy supply, facilitating more excellent ATP production (48, 49).
During maturation, striking changes of shape, morphology, and the biochemical content of cardiac mitochondria enable an amble and steady ATP production rate for contraction. PSC-CMs have immature mitochondria localized in the perinuclear region, exhibiting disorganized and fewer quantities and smaller sizes (Fig. 2). By contrast, mitochondria in adult CMs are well organized in a highly mature network that is about 40% of the cell volume (50) and are arranged in a straight line in the orientation of the sarcomeres and attached to SR, leading to efficient ATP transport (51, 52). Cristae, the inner membrane invaginations that offer a sufficient surface area for efficient mitochondrial respiration, present few and poorly aligned within PSC-CMs (53) while closely compacted and organized within adult CMs (54, 55). Meanwhile, compared with late fetal stages, the copy number of mitochondrial DNA (mtDNA) raises three times in adult hearts.
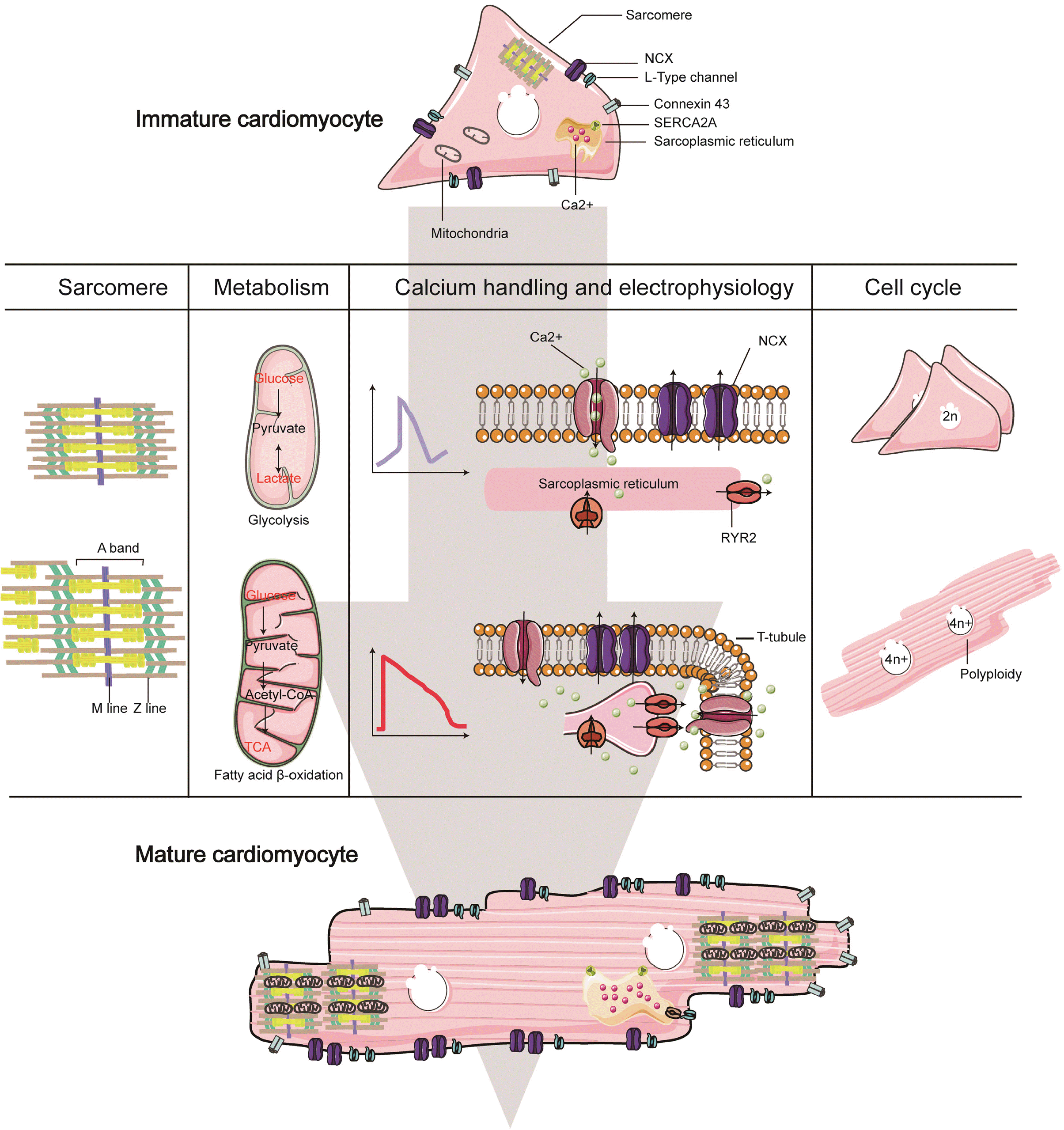 | Fig. 2Major characteristics of cardiomyocyte maturation. Dynamic chan-ges of structure and function of cardiomyocytes occur during maturation. Major characteristics of human pluripotent stem cell-derived cardiomyo-cytes (representing immature cardio-myocyte) and adult-like cardiomyo-cytes (representing mature cardio-myocyte) as discussed in the text.
|
Mitochondrial fusion and fission can direct mitochondrial morphology and size. For instance, disturbance of mitochondrial fusion proteins mitofusin 1 (MFN1), mitofusin 2 (MFN2), or optic atrophy 1 (OPA1) in adult CMs results in a plethora of small, round mitochondria (56). Fission-promoting protein DRP1 overexpression in CMs induces mitochondrial fragmentation (57), partially phenocopied by conditional interruption of MFN1/2.
Changes in metabolic maturation are directly involved in transcriptional activation of genes engaged in fatty acid metabolism and β-oxidation. A crucial part of the peroxisome proliferator-activated receptor (PPARs) in the synergistic cardiac metabolism regulation has been investigated. In common with nuclear receptor gene family members, the PPARs are ligand-dependent transcription factors. The binding of agonist ligands to the receptor leads to PPARs target genes expression level alteration (58). Proliferator–activated receptor γ coactivator-1 (PGC-1) gene expre-ssion is initiated in the postnatal mouse heart, which is identified as a crucial regulatory player by modulating the cardiac mitochondrial number and function response to heart energy demands (59).
Furthermore, inhibition of fatty acid β-oxidation by ETO in the postnatal mouse heart from postnatal days 2 to 4 preserves endogenous CMs proliferation, whereas retards CMs hypertrophic growth and maturation at P5 and P7. On the contrary, fatty acid β-oxidation activation in the infant mouse heart leads to maturation enhancements and gives rise to binucleated CMs at P5 (60). These findings reveal in different cellular-context, PPARs have a different impact on cellular function.
Likewise, another group in the nuclear receptors superfamily, the estrogen-related receptors (ERRs), have also been shown a significant role in supporting the metabolic transition of developing CMs. Supporting evidence shows ERRs have three family members, ERRa (Esrra), ERRb (Esrrb), and ERRg (Esrrg) (61, 62). ERRγ and PGC-1β are vastly upregulated in tissues with high energy requirements such as the heart, kidneys, and brown adipose tissue (63, 64). A recent study shows knockdown of the expression of ERRα and γ in the heart after birth in mice causes cardiomyopathy with an arrest in mitochondrial maturation (65). Using RNA-seq and ChIP-seq, results reveal that ERRγ activates transcription of genes involved in nearly all aspects of postnatal developmental maturation, including mitochondrial energy transduction, contractile function, and ion transport. In common with PPARs, ERRγ expression is also augmented in patients with hypertrophic cardiomyopathy (HCM) and cardiac hypertrophy animal models. Besides, overexpression of ERRγ in the heart induces cardiac hypertrophy. Thus, the promising results indicate functional collaboration between ligand-dependent and orphan nuclear receptors in stimulating CMs maturation.
Go to :
Calcium Handling and Electrophysiology
The sustained contraction and relaxation of the heart are strictly controlled by electric pulses and the oscillation of cytoplasmic Ca2+ concentration. Inward and outward channels of ions result in action potential, the form of the electrical signal that occurs during each cardiac cycle. When CMs are excited, depolarization and repolarization occur, forming action potential (AP). During the change of membrane potential, the ion channels undergo the transition of closing, opening, and deactivation. AP is traditionally characterized by five phases (0∼4) (66), and phase 0 is rapid depolarization, resulting from Na+ rapid influx. Phase 1 is the initial stage of rapid repolarization, which is caused by the transient outflow of K+. The plateau stage of the two phases is slow repolarization, which is caused by the influx of Ca2+ and a small amount of Na+ and the outflow of K+. Phase 3 is the end of fast repolarization caused by the outflow of K+. The AP duration from phase 0 to phase 3 is called action potential duration (APD). Phase 4 is the resting phase, and the membrane potential of the nonautonomous cells is maintained at the resting level. The intramembrane potential is negative to the extracellular potential in the resting state, which is about - 90 mV. It is in the polarized state caused by the high concentration of K+ efflux from CMs. A complete discussion of AP phases in atrial and ventricular CMs can be found in ref (67). In contrast, field potential (FP) is used to describe the membrane potential of CMs measured by multi-electrode array system (MEA). The detected FP signal comprises the spatiotemporal electrical activity of a group of cells; therefore, it is the sum of all the channel currents. In this regard, the FP is similar to the ECG signal that records the change of body surface voltage caused by the current flowing throughout the heart. The FP waveform reveals different characteristics magnified by a pronounced transient spike during membrane depolarization (68), followed immediately by mild slope associated with calcium influx, terminated with repolarization based on potassium efflux. FPD has been demonstrated to correlate with the APD90 of the APs (68). hPSC-CMs together with the MEA system can be widely used to test cardiac drugs effects and represent a promising in vitro model for cardiac electrophysiologic studies (69-73).
Atrial (neonatal and adult), as well as ventricular (embryonic, neonatal and adult) murine CMs, show substantial changes in AP morphology during development (74-77). The inward rectifier potassium current IK1 I nsufficiency is widely acknowledged as the main limitation for hiPSC-CMs. During murine fetal development, the total amplitude of IK1 in ventricular CMs and the activation kinetics increase, and the IK1 inward rectifier property becomes stronger. IK1 plays a crucial role in the spontaneous AP in fetal ventricular CMs (78). They block IK1, resulting in significant depolarization.
Moreover, iPSC-CMs can reiterate the cellular electrophysiological phenotype caused by SCN5A mutations. Remarkably, hiPSC-CMs reveal a relative increase in the adult Nav1.5 over the fetal Nav1.5 after extended culture (79). Some studies report that after>60 days in culture, L-type calcium channel in hPSC-CM is remarkably comparable to L-type calcium channel densities measured in adult human ventricular (80). The action potential of immature hPSC-CMs does not display an exact plateau phase (Fig. 2).
In contrast, mature ventricular CMs open the L-type Calcium channels (Cav1.2) to permit high levels of calcium influx, which generates a plateau phase (80). The slow recovery kinetics of transient outward potassium current (Ito) coupled with a depolarized MDP are responsible for an AP notch deficiency in the majority of hiPSC-CMs. Electrophysiological analysis has also recognized that Ito is small or virtually nonexistent in neonatal CMs from other mammalian species (81).
The action potential is transmitted to the trigeminal structure of the transverse tube and the terminal cistern of the bilateral muscle ganglia through the transverse tube system. The signal is transmitted to the nearby sarcoplasmic reticulum through the transverse tube, which results in the opening of calcium channels on the terminal cistern, where it triggers the cardiac ryanodine receptor 2 (RYR2) to liberate Ca2+ from the sarcoplasmic reticulum (SR); thus the calcium ions in sarcoplasmic reticulum follow the concentration gradient and enter the cytosol with low calcium concentration at rest (82). When excitation-contraction coupling occurs, the concentration of calcium ion in the cytosol can be increased by 100-fold.
Adult CMs are somewhat more giant cells, and Ca2+ signals have a high homogeneity (82, 83). By contrast, hiPSC-CMs express the same components for calcium handling, and the spatial distribution of Ca2+ signals is highly heterogeneous. During in vitro maturation, calcium store load steadily is increased. Notwithstanding, <40 days post beating, hESC-CMs still express functional intracellular calcium handling components, such as CaV1.2, CaVβ2, RyR2, and IP3R, even if its level is significantly lower than those of primary adult CMs.
Go to :
Cardiomyocyte Proliferation
Due to its relationship with cardiomyocyte proliferation, the study of cardiomyocyte maturation is also of great significance. Studies in humans and rodents demonstrate limited regeneration of CMs in adulthood, whereas CMs present proliferative capacity in the fetus (84-86). Tri-iodo-L-thyronine (T3), which is believed as a major stimulant of CMs maturation, suppresses the proliferation of fetal CMs in vitro (87). The deactivation of thyroid hormone signaling diminishes the polyploidization of mouse CMs, postpones the exit of the cell cycle, and preserves the heart regeneration potential of adult mice (88). Similarly, overexpression of YAP5SA, whose target genes encode cell cycle regulators, induces adult CMs reversion to a fetal-like cell state (89). Although miR-199a promotes the dedifferentiation and proliferation of CMs, drops the infarct range, and recovers the contractile function of CMs in infarcted pigs, the uncontrolled expression of this miRNA ultimately causes abrupt death (90). Thus, we should pay more attention to the experiment of promoting cardiac proliferation in the clinic.
Cardiomyocyte cell cycle withdrawal is observed within the first postnatal week of life in mice. Studies in humans have demonstrated the number of CMs remains stable over the human lifespan (91). During maturation, another hallmark is cardiomyocyte polyploidization. In rodents, >75% of CMs become polyploid and largely binucleated resulting from DNA synthesis and nuclear division without cytokinesis (92). By contrast, a similarly high percentage of human and other primate CMs are polyploid containing high DNA contents ranging from 4c to 16c due to DNA synthesis without karyokinesis (93) (Fig. 2). Moreover, the number of polyploid cells improves after myocardial infarction and other injuries. Cardiomyocyte polyploidization is likely negatively correlated with regenerative capacity. Inactivation of tnni3k causes mononuclear diploid CMs to enhance cardiomyocyte proliferation. In turn, overexpression of tnni3k in zebrafish promotes myocardial polyploidy and impairs heart regeneration (94). Relevant evidence shows that ect2 induces zebrafish cardiac polyploidy, an obstacle to the proliferation of CMs (95). However, though polyploidy is related to the terminal differentiation of CMs, polyploid hepatocytes devote robustly to liver regeneration despite ploidy. Thus, it remains obscure about the regulation and unique function of polyploids and their impact on cell physiology.
The classic regulators of cell cycle progression, including cyclins and CDKs, are also engaged in the cardiac cell cycle. These regulators are highly expressed during embryonic cardiac development and then decline at different rates during cardiomyocyte maturation (96). Likewise, the expression levels of cyclins and CDKs are diminished during hPSC-CM differentiation (97). A recent study has reported that overexpression of four cell cycle regulators, cyclin-dependent kinases 1 and 4 (CDK1 and CDK4), cyclin B1, and cyclin D1, induces cell division explicitly in adult mitotic mouse, rat, and human CMs. Once the cell-cycle regulators are delivered after myocardial infarction, the mice will reveal significant cardiac function (98). Overall, such knowledge not only deepens our understanding of cardiac maturation but may also shed light on the discoveries of therapeutic targets to induce host CMs division under disease conditions or to expand transplanted cells after transplantation into damaged hearts.
Go to :
Cardiac Maturation Regulation
Cardiac maturation involves a variety of molecular events coinciding. However, the functional implications of these cellular changes and interactions have only begun to be explored at the molecular level.
Go to :
Transcriptional Regulation
Transcriptome
Microarray analysis demonstrates that the gene expression profile of hPSC-CMs resembles that of the fetal heart in the first trimester. However, after inducing maturation under culture conditions containing the T3 hormone, its gene expression profile is analogous to that of the fetal heart in the second trimester (99). A finding from over 200 microarray data sets from different heart development stages indicates that PSC-CMs mature in the early stage but stop maturing even after 20 days of culture in the late embryonic stage. They found that transcriptional regulators, including PPARs, are misregulated in PSC-CMs, which is the leading cause of maturation stagnation (97). Recently, improvements in single-cell RNA-sequencing technology have profound insight into the regulatory networks in CMs maturation. Single-cell RNA sequencing is performed on more than 1200 mouse cells at seven developmental time points ranging from embryonic day 9.5 to postnatal day 21 (100). They subsequently observe the hES-D20 cells equivalent to E14.5 ventricular CMs, while hES-year1 cells equivalent to E18.5 ventricular CMs. Moreover, they also observe maturity heterogeneity of individual CMs even at the same target time point. This finding suggests that cardiomyocyte maturation state can be observed and assessed at a single-cell level. By taking advantage of extensive single-cell transcriptomic analyses of in vitro cardiac differentiation, another group found HOPX (101), as a critical regulator of heart development, is not efficaciously triggered during monolayer-based cardiac differentiation. Overexpression of HOPX increases cell size and a penal of known regulators of hypertrophy. Loss of HOPX function attenuates hypertrophic growth and maturation.
Master regulators
Currently, many studies have reported on factors that regulate diverse respects of heart maturation. We have already discussed the structure, metabolism, and electrophysiological characteristics of cardiomyocyte maturation in previous sections. However, a central open problem is whether these mature steps are regulated separately, whether they have a standard regulatory process or primary and secondary regulatory relationships. Although some studies reveal sarcomere maturation is upstream of most other cardiomyocyte maturation aspects, the relationships between the multiple aspects of CM maturation remain mostly unresolved. Here, we highlight a few factors that are known for regulating nearly every aspect of cardiomyocyte maturation. These factors are considered as the critical node in the cardiac mature regulatory network (Fig. 3).
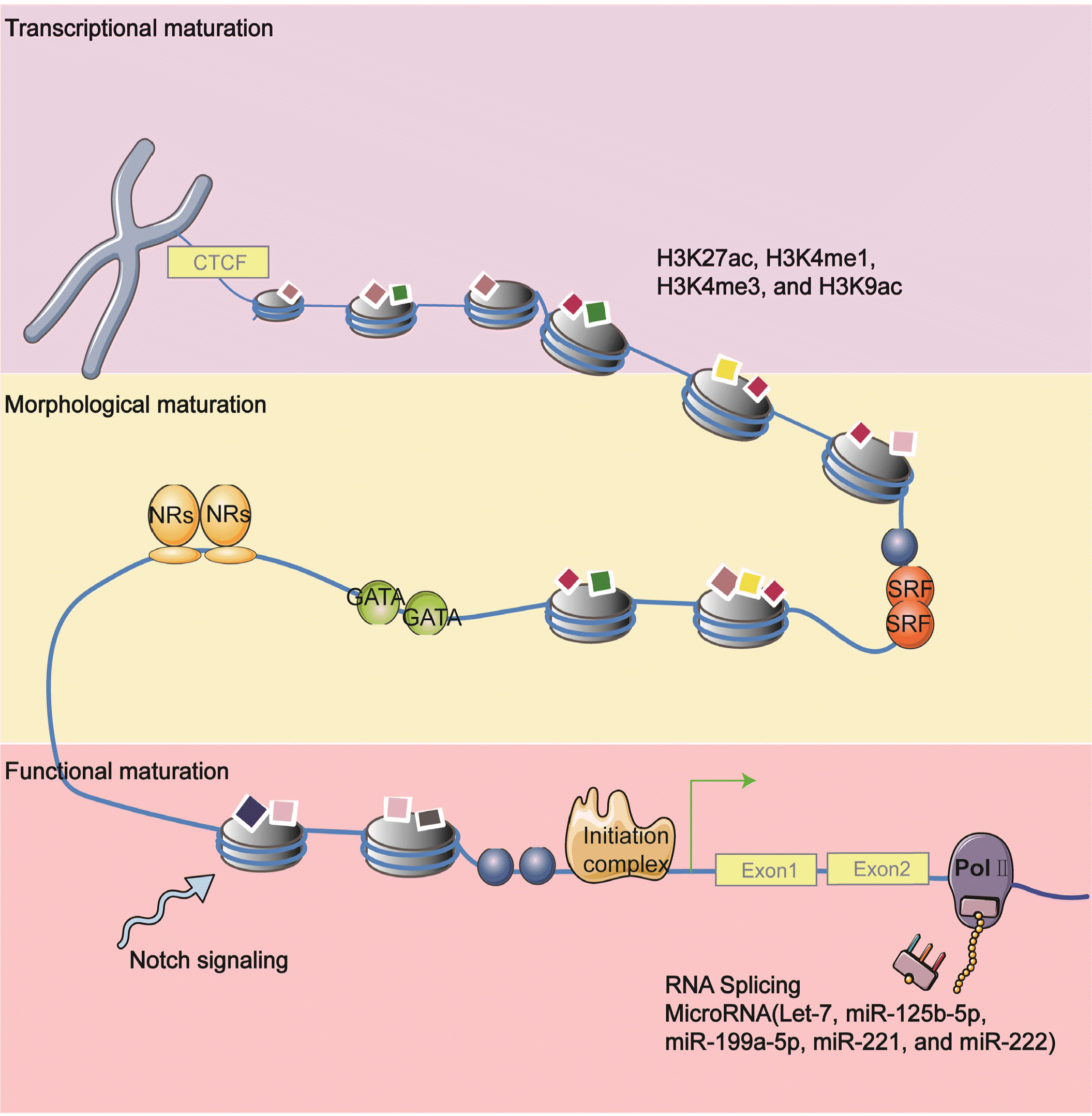
SRF
SRF, a MADS-box-containing transcriptional factor, has been characterized extensively recently (102). In early embryogenesis, SRF has an essential role in mesoderm formation during gastrulation (103). Also, the cardiac-specific absence of SRF in the embryo leads to severe cardiac defects (104). Besides its impact on development, disruption of SRF in the adult heart induces progressive impairment of left ventricular function and progression to dilated cardiomyopathy (105). Intriguingly, SRF is increasingly well implicated as crucial nodes in the regulatory network of cardiac transcription and regulated the cardiac transcriptome (106). Guo and colleagues recently discovered that SRF could coordinate almost all aspects of CM maturation. They first observe a significant reduction of mitochondria and metabolism genes after deleting SRF by exploiting CASAVA. BioChIP-seq further validates SRF directly regulates genes that control sarcomere expansion, mitochondrial biogenesis, transverse-tubule formation (106). However, overexpressed SRF in neonatal CMs also dramatically blocks T-tubule formation and causes defects in morphological maturation. Thus, the SRF dosage should be carefully balanced in promoting CM maturation.
Nuclear receptors
Thyroid hormone T3 profoundly impacts cardiomyocyte maturation, including regulating fetal-to-adult titin and myosin heavy chain isoform transition (107), increasing cardiomyocyte width polyploidization, augmenting expression of phospho-mTOR, ANP, and SERCA2a, reducing proliferation and cyclin D1 protein (88).
T3 functions via THRA and THRB nuclear receptors, which are the central thyroid hormone receptors. Mutation of THRA impairs cardiomyocyte maturation. Likewise, glucocorticoids have a critical role in late gestational heart maturation (108). Glucocorticoids bind to glucocorticoid receptors (GRs), which belong to the nuclear receptors superfamily. Mice deficient in glucocorticoid receptors in CMs and vascular smooth muscle reveals aberrant cellular behavior, including poorly aligned and disorganized myofibrils with only a few sarcomeres (109). Additional nuclear receptors (NRs) play a central role in regulating metabolic maturation. Such factors are PPARs (peroxisome proliferator-activated receptors), which interact with the retinoic acid X receptor and activate downstream targets engaged in fatty acid and carbohydrate metabolism by binding to the promoter region. It has been believed that PPARα is activated by fatty acids and a primary cardiac fatty acids metabolism regulator. PPARα-mediated activation of fatty acid β-oxidation promoted the proliferation of CMs at P4 in infant mice, while this phenomenon did not exist in P2 and P5. However, in P5, PPARα-mediated activation of fatty acid β-oxidation enhances the hypertrophic growth and maturation of CMs. These facts highlight the role of PPARα in distinct cellular functions during perinatal environmental changes (60). PPARγ coactivator–1 (PGC-1) promotes transcription via complex assembly that anchors active nuclear receptors with chromatin remodeling complexes. Mice deficient in both PGC-1α and PGC-1β demonstrated signatures of a maturational defect, including reduced growth, arrested mitochondrial biogenesis, and persistence of a fetal pattern of gene expression (110). Of note, depletion of PGC-1α and PGC-1β during postpartum heart development leads to significant defects in mitochondrial maturation and structure, related to decreased expression of genes involved in mitochondrial fusion-fission, such as Mfn1, Opa1, and Fis1, resulting in progressive, lethal cardiomyopathy (111). More recently, a study shows PGC1 is essential for postnatal CM growth, calcium handling, and mitochondrial activity beyond metabolism, which suggests that PGC1 has a multi-faceted role in coordinated cardiomyocyte maturation (112). Mechanistically, PGC1/PPAR signaling can regulate Yap1 and SF3B2, which are upstream regulators of cellular hypertrophy and calcium handling.
Apart from ligand-dependent nuclear receptors, ligand-independent (orphan) nuclear receptors such as the ERRs (estrogen-related receptors α, β, and γ) might also be essential for the maturational transition of developing CMs by directly regulating genes vital for mitochondrial functions, CM contraction, calcium homeostasis, and conduction (65, 113).
Therefore, NRs deserve more attention as potentially promising candidates for metabolic variations and transcriptional regulation during cardiac maturation.
Go to :
Posttranscriptional Regulation
miRNAs have been established as critical factors in coordinating the sophisticated regulatory network in cardiomyocyte maturation. For example, miR-1 is the most abundantly occurring cardiac microRNA (miRNA) in mature CMs, verified to facilitate their electrophysiological maturation (114). Moreover, Let-7 is the most upregulated miRNA family member during the one-year culture of hESC-CMs in vitro (115). Cell size, sarcomere organization, contraction force, and respiratory capacity enhancements are observed after let-7 family overexpression in hESC-CMs. By contrast, miR-200c overexpressing hESC-CM reveals Ca2+ influx inhibition (116). It is reported that microRNA maturation cocktail (that is, overexpression of Let7i and miR-452 and knockdown of miR-122 and miR-200a) while significantly enlarges hiPSC-CMs cell area and creates a more mature transcription profile (117). Similar to the cocktail strategy, delivering four microRNAs, miR-125b-5p, miR-199a-5p, miR-221, and miR-222 (termed as miR-combo), to m/hESC-CMs also leads to improvement of sarcomere alignment and calcium handling, mitochondrial cristae formation, and enhance expression of cardiomyocyte maturation markers (118).
Epigenetic regulation is central to establish and maintain vast various cellular functions. Epigenetic marks or factors, such as DNA methylation and histone tail modifications, are dynamically changed during cardiac development and maturation. Emerging evidence indicates the epigenetic alterations are closely associated with cardiomyocyte maturation. As expected, epigenetic changes are relevant to transcriptional activity and silencing. In postnatal CMs, fetal cardiac genes are decked with repressive chromatin configuration, characterized by hypermethylated and H3K27me3. Whereas actively expressed genes in adult CMs are hypomethylated and maintain active histone modifications, such as H3K27ac, H3K9ac, H3K4me1, and H3K4me3 (119, 120). Valproic acid, the histone deacetylase inhibitor, increases active histone modifications H3K4me3 on the whole genome level, induces hypertrophic growth, and augments cardiac gene expression hPSC-CMs (121), but it does not have an impact on electrophysiological properties. DNMT3A/B-mediated DNA methylation can inhibit the slow skeletal troponin I subtype (Tnni1).
Similarly, genes involved in converting fetal into adult energy metabolism are also methylated after birth. H3K27me3 is particularly related to suppressed and demethylated genes in CMs (119). In another study, early cardiac progenitor cells stimulated with polyinosinic-polycytidylic acid (pIC) have been shown to augment the maturation of CMs (122). Mechanistically, pIC treatment regulates early Notch signaling and increases the epigenetic activating modification H3K9ac in cardiac myofilament genes promoter regions. In a recent study, in vivo CRISPR screening identifies RNF20/40 (123), which monoubiqui-tinates H2B at lysine 120 by exerting E3 ligase activity. RNF20/40 is essential for CMs maturation, which is proven to regulate metabolism during CM maturation directly.
Chromatin organization is believed as another fundamental regulatory layer of CM maturation. Chromatin-remodeling protein, BRG1, maintains cardiomyocyte in an embryonic state. When encountering cardiac stress in adulthood, BRG1 is reactivated and cooperates with histone deacetylase (HDAC) and poly ADP–ribose polymerase (PARP) to induce myosin heavy chain isoform conversion (124). CTCF (CCCTC-binding factor), one of the best described architectural proteins, hearts of its mutant reveal mitochondrial function and protein production genes upregulation; however, mitochondrial do not mature correctly (125).
In conclusion, these findings suggest that histone and DNA modification play a significant role in regulating cardiomyocyte maturation. Nevertheless, we are still in the early stages of understanding how epigenomes regulate this sophisticated progress; the target and mechanism of CM maturation need to be further explored.
Cardiac proteomics promotes our understanding of regulating CM maturation. As anticipated, the global proteome of hESC-CMs is fetal-like, and hESC-CMs have the lowest level of sarcomeric protein, a protein engaged in energy transfer. Pathway analysis highlighted the peroxisome proliferator-activated receptor α signaling (PPARα) as a key regulator for cardiac maturation. Activation of PPARα with molecular agonist significantly increases fatty oxidative enzyme activity, hyperpolarizes mitochondrial membrane potential, and induces a more organized morphology (126). Using a mass spectrometric approach to investigate protein expression in vitro over 30 days of hiPSC-CM, the finding suggests proteins associated with protein translation/synthesis and ubiquitination are reduced, followed by an increase in oxidative phosphorylation and a decrease in glycolytic proteins. Although most proteins involved in excitation-contraction coupling are increased, this is insufficient to cause functional enhancement because of no change in calcium transient amplitude (126). In a recent study, Cai et al. (127) develop an unbiased proteomics strategy integrating high-throughput top-down targeted proteomics and bottom-up global proteomics to appraise precisely hPSC-CM maturation. This finding identifies several candidate maturation-re-lated factors critical for sarcomere tissue, cardiac excitability, and calcium homeostasis.
Go to :
Approaches to Acquire CM Maturation
The ability of hPSC-CM to mature into an adult-like phenotype after transplantation denotes that the standard cell culture conditions lack key elements in the in vivo environment (128). On the other hand, isolated adult CMs are observed to either die or dedifferentiate after being cultured in a cell medium for several days (129). The above limitation has been recognized for many years. Thus, many researchers are attempting to improve the in vitro systems to promote mature CMs better (Fig. 4).
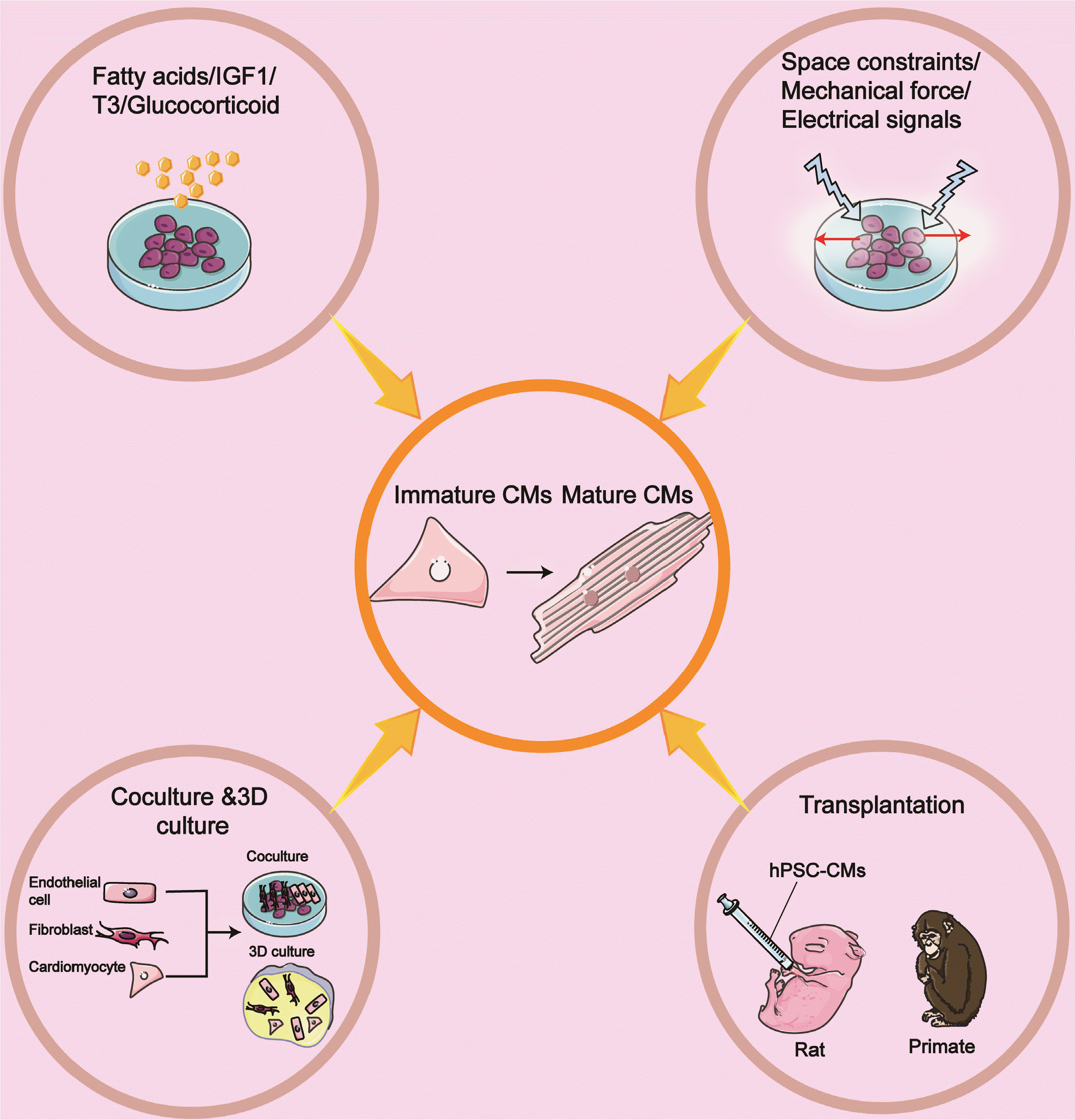
Go to :
Biophysical Stimuli
Using matrigel-coated polyacrylamide substrates with a physiologically relevant stiffness in rectangles [length: width=1.5∼1.7:1], micropatterns make CMs more mature concerning electrophysiology, calcium flow direction, mitochondria organization, and T-tubules formation. hPSC-CMs growth on hydrogel also displays action potential propagation and myocardial contractility (130). hPSC-CMs island geometry has a significant effect on action potential and calcium dynamics. hiPSC-CM in larger islands demonstrate maturity enhancements, explained by nearest-neighbor contact interactions (131). Similar findings were observed when GSK-3β inhibitor withdrawal combined with high cell-cell contact (132).
The mechanical force also modulates maturation. hPSC-CMs cultured on physiologically stiff 16 kPa show sarcomeric alignment and SERCA2a spreading and relocalization (133). hiPSC-CMs seeded on Matrigel with >0.4 mm thickness exhibit enhanced sarcomere alignment, rod-shaped and robust hiPSC-CM shortening (134). A recent study reports CMs derived from early-stage display significant plasticity instantly after the occurrence of spontaneous contractions, under the conditions of physical conditioning with growing density for four weeks, while cardiac tissue generated from human iPSCs display adult-like characteristics, such as organized ultrastructure, physiological sarcomere longitude and mitochondria density, the presence of transverse tubules and oxidative metabolism (135). However, electromechanical properties have not reached the maturity degree recapitulating the in vivo adult myocardium.
Applying electrical signals can also substantially enhance cell function. Suprathreshold electrical field stimulation and chronic pacing at a constant rate both ameliorate the neonatal rat ventricular myocytes’ functional phenotype (136, 137).
The above strategies have also been applied for hPSC-derived cardiac tissues. Nunes and colleagues have generated a platform termed biowire that combines 3D and electrical stimulation with an increased frequency ranging from 1 to 6 Hz to create a microenvironment favorable to cardiac maturation (138). Strikingly, these stimulated conditions remarkably improved cell and myofilament structure enhanced electrophysiological and Ca2+ handing and upregulated potassium inwardly-rectifying channel gene. However, M-lines, T-tubules defects, and downregulation of structural proteins mRNA are observed in this system. The same group developed the second version, named Bioware II (139), which enables electrophysiologically distinct atrial and ventricular tissues. Electrical stimulation matures CMs via increasing connexin expression and adapting an autonomous beating rate; this adaptive effect can maintain for up to 2 weeks.
Go to :
Biochemical Stimuli
The hormone-insulin-like growth factor 1 (IGF-1) signaling has vital roles in regulating several cellular processes, including contractility, metabolism, hypertrophy, autophagy, aging, and apoptosis in the heart (140). The canonical IGF1 pathways involve MAPK(RAS/RAF/MEK) and PI3K/AKT/Mtor pathways (141). IGF1 binds to the IGF1 receptor (IGF1R), a cell surface tyrosine kinase receptor needed in physiological stresses-induced hypertrophy (142) to activate pathways. IGF1, in turn, activates PI3K–AKT1 signaling to promote the proliferation of CMs derived from hESCs (143). Cardiac hypertrophy with an increase in myocyte size and enhanced systolic function is observed in the heart of overexpressing IGF1R mice (144). hESCs-CMs in engineered cardiac tissues (ECTs) stimulated with IGF1 and neuregulin 1β (NRG1) exhibit increased area and improved force-frequency relationship (145).
A switch from glycolysis to fatty acid metabolism is the hallmark of postnatal cardiomyocyte maturation, which is exploited for developing methods to improve in vitro CM maturation. Fatty acid supplementation can further advance hPSC-CM maturation, accompanied by enhanced force generation and augmented mitochondrial respiratory reserve capacity (146). In one study of a synergistic impact, galactose and fatty acid are added and vastly improved hPSC-CM maturation, characterized by a higher oxidative metabolism, increased myofibril density, and alignment enhanced AP durations, and higher upstroke velocities (147). Glucose deprivation of hESC-CMs is more elongated and displays functional maturation at the metabolic, electrophysiological, and biomechanical levels. In terms of mechanism, nucleotide biosynthesis via the pentose phosphate pathway is the pivotal regulator of the promitotic/anti-maturation effect of glucose (148). Mean-while, glucose aberrantly induces hypoxia-inducible factor 1-alpha (HIF1α) and its downstream glycolysis-related genes. HIF1α/LDHA inhibition results in oxidative phosphorylation improvement. Conversely, this promotes metabolic and functional maturation of hPSC-CMs (149).
As described above, the administration of T3 or glucocorticoid facilitates cardiomyocyte maturation (150, 151). The synergetic effect of biochemical signals on cardiomyocyte maturation has also been investigated. During the cardiac differentiation, combined thyroid and glucocorticoid hormones motivate T-tubule development and more ventricular-like EC coupling. Treatment CMs with thyroid hormone, dexamethasone, and IGF1 in 3D cardiac microtissues (CMTs) yields a more adult-like cardiac performance (152).
Go to :
Co-Culture and 3D Culture
CMs account for about 25∼35% of all cells in the heart. Conversely, non-CMs include endothelial cells, hematopoietic-derived cells, immune cells, and fibroblasts, which make up the vast majority of the heart’s cell (153). Co-culture of non-CMs and CMs promotes the maturation of hPSC-CM. For example, it is reported that non-CMs in embryoid bodies (EBs) drive electrophysiological maturation of early-stage cultures of hESC-CMs (154). Co-culture enables hiPSC-CM to be more mature partially through paracrine factors secreted from non-CMs (155). Fibroblasts have stage-specific roles in modulating cardiac function. Embryonic cardiac fibroblasts induce the proliferation of CMs, whereas adult cardiac fibroblasts promote myocyte hypertrophy (156). Co-culture of hPSC-derived ECs with hPSC-derived cardiac progenitor cells (CPCs) can increase CM size, enhance sarcomere proteins and their organization (157).
The use of hiPSC-CM and other cardiac cell types for 3D culture is a promising approach to improve maturation. Many research groups have demonstrated 3D culture systems (such as engineered tissues and organoids) can improve maturity (135, 158-160). For example, hESC-CMs are mixed with human primary cardiac microvascular endothelial cells and cardiac fibroblasts in spheroid microtissue, which leads to greater Ca2+ transient amplitudes and enhances spontaneous contraction rate, and remarkably accelerates contractile function (161). Similar results are obtained in another study. The tri-cellular cocultivation of hiPSC-CMs, cardiac fibroblasts (CFs), and cardiac endothelial cells also enhances maturation in three-dimensional microtissues (MTs) (162). They found cyclic AMP (cAMP) is responsible for connexin 43-mediated hiPSC-CM maturation. As mentioned above, additional biochemical (T3, dexamethasone, IGF1, palmitate) and biophysical stimulation (electrical pacing; mechanical stress) on these 3D contexts are essential to produce adult-like CMs (152, 163).
In summary, these studies demonstrated that co-culturing hPSC-CMs with fibroblasts and endothelial cells mimics the intercellular crosstalk environment in vivo. 3D systems further boost maturation in vitro. These technologies can significantly promote CMs maturation and are beneficial to discover novel cardiomyocyte maturation regulators. Notably, tissue-engineering techniques are challenged to widely spread because it is expensive, time-consuming, and challenging to understand downstream mechanisms. Therefore, we will need additional studies to develop a more economical and convenient technique for clinical application.
Go to :
Transplantation
Neonatal and adult rat heart transplantation can promote the maturation of hiPSC-CMs. In both stage hearts, engrafted cells develop partially mature myofibrils accompanied by cell enlargements, sarcomere lengthening, and more cardiac troponin I expression (164). However, compared with the host cells, engrafted derivatives are still much more minor. This phenomenon could be related to species incompatibility. A landmark study from the same group reported that after transplantation hESC-CMs into the myocardial infarcted heart, cardiac function is recovered at unprecedented levels accompanied by forming electromechanical junctions with the host heart (165).
Together, these studies demonstrate that hearts can provide the native environmental cues essential for guiding PSC-CMs to mature toward a nearly adult-like state. The cues might come from chemical signaling through gap junctions, paracrine factors from neighboring cells, and systemic circulation factors (such as metabolic or hormone-related). Further analysis will be needed to explore the novel factors and investigate their impact on morphological and functional maturation.
Go to :
Conclusions
In conclusion, we review the main features of cardiomyocyte maturation and the known regulators in this sophisticated process. Although the significant differences between immature CMs and mature CMs have been well documented, the details of the molecular mechanisms involved in the transition from immature to mature states remain ripe for discovery. Indeed, accumulated evidence shows maturation can be a complex trait governed by multiple signaling networks in the cytoplasm and nucleus. Therefore, the research in this field should focus on individual characteristics and how to synchronize the events.
On the other hand, although individualized intervention has promoted the development of CMs to a more mature phenotype in vitro, these methods are not enough. It seems that combinatorial approaches might be necessary.
With the development of the model system and the increasingly close cooperation between basic scientists and tissue engineers, a more comprehensive map of cardiomyocyte maturation can be guaranteed in the foreseeable future. This work is essential for designing better strategies to mature PSC-CM, stimulate cardiomyocyte regeneration, and treat diseases involving defects in cardiomyocyte maturation.
Go to :
 XML Download
XML Download