

 PDF
PDF Citation
Citation Print
Print


Introduction
Head and neck cancer, including oral cancer, is the sixth leading type of cancer worldwide (1), representing approximately 6% of all solid tumors. Approximately 90% of diagnosed cases of head and neck cancers are oral squamous cell carcinoma (SCC), which is the most common malignancy in the oral cavity (2). In recent decades, the five-year survival rate after treatment of oral cancer is still approximately 50% for most countries (3-5), which indicates a poor prognosis for the developing world (6). Several chemotherapeutic agents for the treatment of oral cancer have been tested individually or in combination with other anticancer drugs, such as cetuximab, cisplatin, docetaxel, 5-fluorouracil, methotrexate, and paclitaxel (7-10). However, even in patients that are most sensitive to these chemotherapeutic drugs, they experience tumor recurrence due to the emergence of genetically modified cell populations, which ultimately leads to drug resistance and tumor re-growth (11). Indeed, the recurrence or metastasis rates for oral squamous cell carcinoma vary from 18% to 76% in patients undergoing standard treatment (12). The CSC theory provided a supportive explanation for drug resistance and subsequent tumor recurrence after chemotherapy. CSCs were first identified and characterized in the bone marrow of acute myeloid leukemia and described as key “seeds” for tumor initiation, growth, and therapy resistance (13). Currently, CSCs have also been found in almost all types of solid tumors, including those in the brain (14), colon (15), gastric (16), head and neck (17), lung (18), pancreatic (19), and prostate cancers (20), and are defined as a small subpopulation of tumor cells with stem cell-like characteristics. Unlike bulk tumor cells, they have both tumor-initiating potential and the capacity to reconstitute the bulk tumor with cellular heterogeneity, as observed in healthy tissues (21, 22). Furthermore, oral CSCs are commonly defined by several unique genomic and functional properties: (a) oral CSCs are intrinsically resistant to the majority of conventional chemotherapeutic agents; thus, the remaining CSC subpopu-lation can subsequently regenerate tumors in patients (23-25), (b) oral CSCs can be characterized by certain pluripotency-associated markers (22, 26-28), and (c) CSCs can survive and form 3D tumor spheres in suspension culture conditions (29-31). These functional characteristics of CSCs indicate that the majority of traditional therapeutic strategies, such as surgery with chemotherapy and/or radiation, can only kill the bulk tumor cells but leave rare, oral CSC subpopulations untouched. In this context, new therapeutic strategies that can effectively eradicate oral CSCs would ultimately improve treatment outcomes and enhance the quality of life of patients with oral cancer (32). However, there is not enough information currently available to make a conclusive statement regarding oral CSC-targeted therapeutic strategies. In this study, we provide current information about several genomic and functional properties of oral CSCs and discuss various therapeutic approaches targeting oral CSC-specific signaling pathways and pluripotency-associated markers involved in the maintenance and drug resistance of CSCs.
Go to : 
The Origin of CSCs from Somatic Stem Cells or Differentiated Cells
Given the functional similarities between somatic stem cells and CSCs, researchers have attempted to determine whether CSCs arise from tissue-resident stem/progenitor cells or differentiated cells that acquire stem cell-like characteristics during the malignant process. However, the origin of cancer stem cells remains elusive, because the origin of CSCs varies greatly depending on the malignancy (33, 34). Tissue-resident stem/progenitor cells exhibit unique biological features, such as long-term reconstitution, self-renewal, and transdifferentiation capacity. Functionally, stem cells self-renew asymmetrically but also give rise to daughter cells that are committed to lineage-specific differentiation to reconstitute specific tissues and maintain homeostasis. The tissue-resident stem cells that give rise to cancer is an attractive hypothesis, given that the multistage theory of tumor development requires actively dividing and long-lived stem cells in which consecutive genetic mutations can accumulate (35). Similarly, intermediate progenitor cells, which are more abundant in specific tissues than stem cells and commonly undergo limited cell proliferation, aberrantly acquire enhanced self-renewal capacity, and ultimately become the long-lived target that acquires consecutive genetic mutations (36). Indeed, a number of studies have hypothesized that tissue-resident stem cells or certain cells with stem cell characteristics can directly give rise to CSCs. Some characteristics and functions of leukemic CSCs with hierarchically arranged subpopulations (similar to those observed with normal bone marrow stem cells) support the stem-cell origin hypothesis (37). Interestingly, in some tissues, fully differentiated cells can also revert to the stem cell phenotype through dedifferentiation or reprogramming (38). Therefore, some researchers have hypothesized that cancer cells could be derived from mature and fully differentiated cells that re-acquire stem cell-like properties under certain conditions. In the “differentiated cell origin model”, the requisite of consecutive oncogenic mutations may lead to the dedifferentiation of mature cells into pluripotent stem cell-like cells. The corresponding mechanisms or physiological conditions that determine which cell types would undergo dedifferentiation have not been completely elucidated. Surprisingly, this dedifferentiation process can be accelerated by simple genetic modifications (39, 40). Indeed, Takahashi et al. (40) induced the dedifferentiation of fully differentiated somatic cells into pluripotent stem cells with multidifferentiation potential by introducing only four pluripotency-associated genes (Oct3/4, Sox2, c-Myc, and Klf4). Regardless of their cellular origin, whether tissue-resident stem/progenitor cells or differentiated somatic cells, CSCs are defined as the subpopulation of cells that retain stem cell-like characte-ristics. Similar to normal stem cells, CSCs can divide symmetrically or asymmetrically, giving rise to both other CSCs and more differentiated progenies that ultimately reconstitute the bulk tumor through differentiation into many different cell types.
Go to :
Therapeutic Implications of Oral CSCs in Cancer Therapy
The majority of cancer cells in heterogeneous bulk tumors have limited proliferative potential, tumor-forming capacity, and resistance to chemotherapy. The diverse phenotypic heterogeneity and plasticity among cancer cells in bulk tumors have been explained by the CSC model of hierarchically organized tumors (42). After their identification from leukemia, CSCs were first isolated and characterized from solid breast cancer tumors. The CD44+/CD24−/low and epithelial cell adhesion molecule-positive phenotypes may be typically considered pluripotency-as-sociated characteristics in breast CSCs (43, 44). As few as 100 cells exhibiting these cellular properties can grow rapidly in vitro and effectively form new tumors in vivo after reconstitution (43). The evidence of oral CSCs has been first suggested by the study, which showed that even just a small fraction of oral squamous cell carcinoma cells with “stem-like” characteristics is able to rapidly reconstitute a new bulk tumor after reconstitution (45). The isolation of oral CSCs from tumor masses has mainly been conducted using the cell surface marker CD44, which is an important cell surface marker for isolating stem-like cells from breast cancer (46, 47). Although CD44 was originally identified as a receptor for hyaluronic acid, it has also been suggested as a putative marker for various types of stem cells (48, 49) and a key regulator for the maintenance of the properties of various CSCs (50, 51). Aldehyde dehydrogenases (ALDH), a family of intracellular enzymes that catalyze cellular detoxification and subsequent drug resistance via oxidation of intracellular aldehydes, has been proposed as a putative biomarker for oral CSCs (52). Interestingly, ALDH-positive subpopulations isolated from head and neck squamous cell carcinoma have typical CSC characteristics and enhanced tumorigenic potential in vivo (53, 54). Overall, acquired resistance to standard chemotherapy or radiation therapy is an important cause of treatment failure. Emerging evidence indicate that CSC subpopulations are more resistant to standard chemotherapeutic drugs when compared to non-CSC subpo-pulations (55). CSC-mediated drug resistance has been previously demonstrated in multiple types of cancer, such as brain (56), breast (57), colorectal (58), leukemias (59), skin (60), and pancreatic (19) cancers. In addition, CSC-mediated radioresistance has also been reported in brain (61) and breast (62) cancers. Oral CSCs are also commonly resistant to various conventional chemotherapeutic agents (23-25) and radiation therapies (27, 63, 64) that target proliferating cells (Fig. 1). Therefore, the remaining resistant oral CSCs can proliferate extensively and reconstitute new tumors in patients and may ultimately lead to the failure of durable clinical responses. Therefore, research efforts are underway to develop novel therapeutic strategies for selectively eradicating resistant oral CSCs without affecting normal cells (65-67).
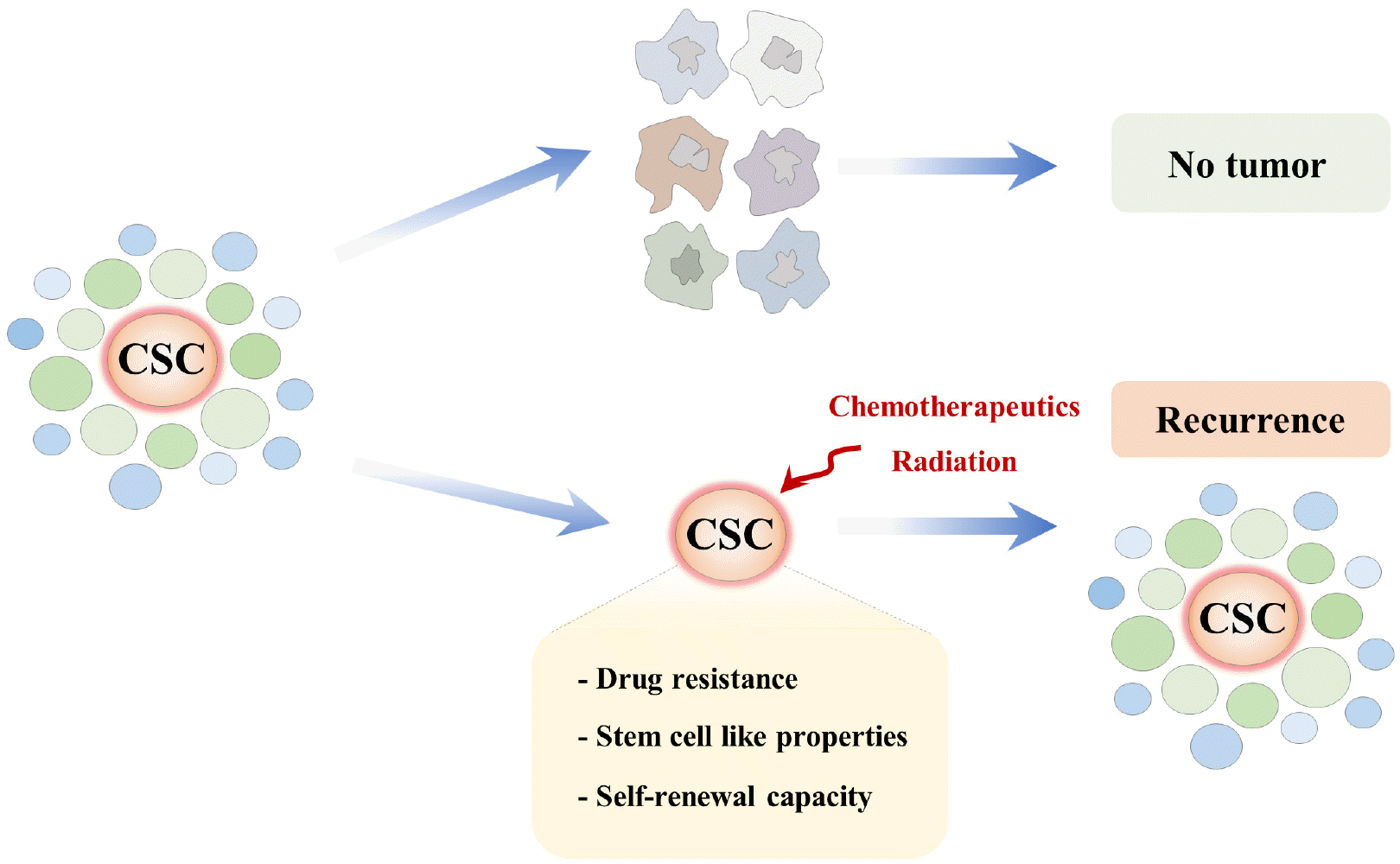 | Fig. 1Schematic diagram summarizing the therapeutic strategies targeting oral CSCs. The diverse phenotypic heterogeneity and plasticity among cancer cells in bulk tumors have been explained by the CSC model of hierarchically organized tumors. Unlike bulk tumor cells, CSCs are intrinsically resistant to the majority of conventional chemotherapeutic agents; thus, the remaining CSC subpopulation can subsequently reconstitute tumors in patients with cellular heterogeneity.
|
Go to :
Several Strategies for the Selective Targeting of Oral CSCs
Therapeutic efficiency could be enhanced by selectively targeting subtle expression differences in surface markers and alterations in various signaling regulators between CSC subpopulations and non-CSC subpopulations. Currently, many researchers have identified several promising therapeutic targets for oral CSCs, including signaling pathways, cell surface markers, pluripotency-associated genes, and transcription factors that can selectively eliminate oral CSCs and subsequently reduce the risk of cancer recurrence (Fig. 2).
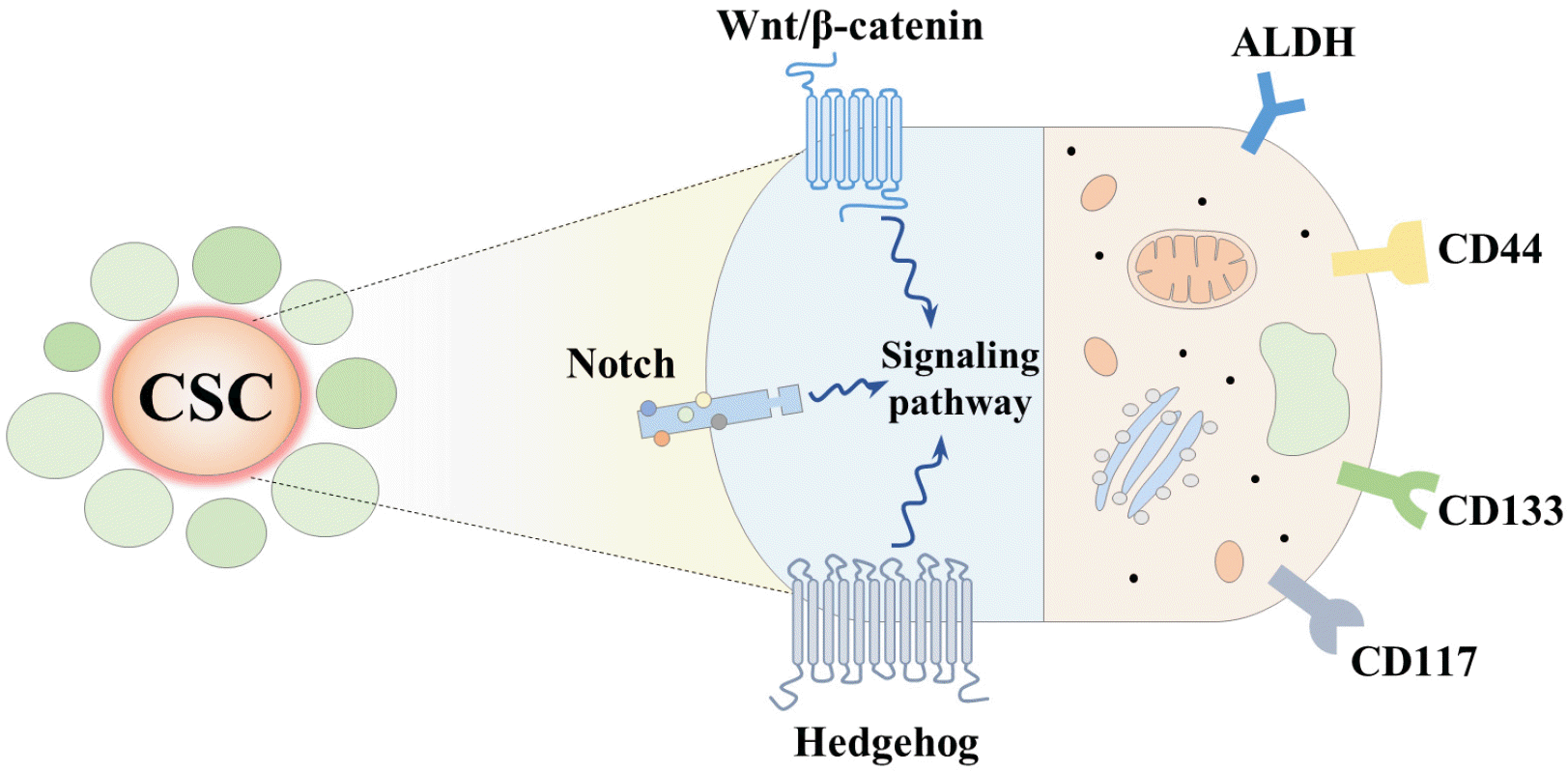 | Fig. 2Schematic diagram summarizing the therapeutic strategies targeting oral CSC-specific surface markers or signaling pathways. Therapeutic efficiency could be enhanced by selectively targeting subtle expression differences in surface markers and alterations in various signaling regulators between CSC subpopulations and non-CSC subpopulations. For this reason, CSC markers can be used to identify CSC-enriched subpopulations and various therapeutic approaches effectively inhibit oral CSCs by targeting pluripotency-associated genes (ALDHs, CD44, CD133, and CD177) and CSC-specific signaling pathways (Wnt/β-catenin, Notch, and hedgehog signaling pathways).
|
Pluripotency-associated surface marker targeting strategy
A rare subpopulation of CSCs can be identified and isolated from the tumor mass by using single or combinations of multiple surface markers, thus providing a more effective CSC-targeted therapeutic strategy (68). Currently, magnetic cell sorting, flow cytometry, fluorescent antibody staining, and real-time PCR are commonly used to isolate and characterize heterogeneous CSCs within a tumor mass. Although considerable progress has been achieved in identifying CSCs on the basis of their specific surface makers, the development of selective CSC therapies remains a challenge, largely due to the many common properties between CSCs and normal stem cells.
Aldehyde dehydrogenase: Aldehyde dehydrogenases (ALDHs) are a superfamily of intracellular enzymes that play important roles in cellular detoxification and subsequent drug resistance by metabolizing various intracellular aldehyde derivatives. Over two decades ago, ALDHs were first known to confer resistance to chemotherapeutic alkylating agents such as cyclophosphamide in hematopoietic and leukemic stem cells (69). ALDH-positive subpopulations were found in breast (70) and brain (71) cancers. In these tumors, cells with high ALDH activity were characterized as having enhanced self-renewal capacity and subsequent tumorigenic potential in vivo, which are typical characteristics of CSCs. Indeed, many recent studies have revealed that high levels of endogenous ALDH activity may be associated with increased in vitro clonogenic activity, tumorigenic properties, and drug resistance in oral CSCs (27, 53, 72, 73). ALDH1 protein expression levels are positively correlated with the tumor formation ability of neck squamous cell carcinoma and negatively correlated with patients’ responses to ongoing treatments (27). Consistently, as few as 5×102 ALDH1-positive cells of head and neck squamous cell carcinoma were able to reconstitute visible tumors in vivo, and these subpopulations showed increased 3D sphere-forming ability in vitro, higher migratory capacity, and enhanced radiation resistance (27, 53). Interestingly, the ALDHhigh subpopulation significantly overlaps with the CSC surface marker CD44-expressing populations (50.6%∼74.4%). Conversely, when CD44-positive subpopulations were sorted for ALDH activity, only 9.8%∼23.6% of the CD44+ cells overlapped with high ALDH activity (53). This result suggests that ALDH activity can be used as a selective marker for CSCs in head and neck squamous cell carcinoma (53). In addition, ALDH+ stem-like populations were also associated with epithelial-to-mesenchymal transition (EMT), which is a key process in metastasis during malignant progression in head and neck squamous cell carcinoma (27).
CD44: CD44 is a transmembrane receptor for hyaluronic acid, which is highly expressed in many cancers, and regulates cell migration and invasion processes (74). The CD44-hyaluronic acid signaling axis can promote tumor progression and subsequent metastasis by increasing self-renewal capacity, cell survival, and drug resistance (75, 76). CD44 can also bind to several growth factors and some metalloproteinases (MMPs), such as MMP-2, MMP-9, and MMP-14, resulting in an increased ability for metastasis (77, 78), angiogenesis (79), and drug resistance (80). CD44-positive subpopulations with high tumorigenic potential have been identified in multiple types of CSCs, such as in cervical (81), prostate (82), lung (83), breast (43), colon (84), ovarian (85), gastric (86), bladder (87), and pancreatic (88) cancer. In this context, the CD44high subpopulation with elevated tumorigenicity and metastatic ability is an attractive therapeutic strategy for the treatment of multiple cancer types. Emerging evidence has revealed the association of CD44 with tumor aggressiveness and its prognostic impact on patients with oral squamous cell carcinoma (89, 90). Consistently, it has been described that certain CD44 (i.e., v3, v6, v10) variant isoforms seem to act as metastasis genes and are associated with metastasis and cancer progression in oral squamous cell carcinoma (91). CD44-positive cell subpopulations express high levels of Bmi-1, which plays a key role in the self-renewal capacity of various stem cell types and is involved in tumorigenesis (22). Since then, many previous studies have suggested that CD44-positive cells exhibit a significantly higher potential for 3D sphere-forming ability in vitro, higher migratory capacity, and drug resistance in both primary tumor tissues and cell lines of oral squamous cell carcinoma (22, 92-96). In addition, the frequency of CD44-positive cells was significantly associated with poor prognosis and higher rates of recurrence and metastasis after radiation therapy in patients with oral squamous cell carcinoma (92).
CD133: CD133 (also known as AC133 and prominin-1), a 5-pass transmembrane glycoprotein with a molecular mass of 120 kDa, has been identified as a novel marker for hematopoietic stem/precursor cells and endothelial progenitor cells in various organs (97). However, several previous studies have indicated that CD133 can also be used as a marker for the identification of CSCs in many solid tumors, including hepatocellular carcinoma (98), oral squamous carcinoma (99), prostate carcinoma (100), renal cell carcinoma (101), and thyroid carcinoma (102). Consistently, CD133-positive subpopulations from the head and neck squamous cell carcinoma cell line Hep-2 have a markedly increased capacity for tumor formation in vivo when compared with CD133-negative cells (103). Previous studies have revealed a significant correlation between enhanced levels of CD133 and poor prognosis in patients with oral squamous cell carcinoma (104, 105). Moreover, CD133-positive cells showed increased drug resistance, self-renewal ability in vitro, tumorigenesis in vivo, and expression levels of pluripotency-associated genes, such as ALDH, NANOG, OCT4, and SOX2 (28). Elevated expression of CD133 was observed in oral cancer stem-like cells from patients with oral squamous cell carcinoma (104). Its expression is also correlated with increased migration, tumorigenicity, and the expression level of the ABC transporter gene ABCG2 (104). Additionally, oral cancer patients who are NANOG/OCT4/CD133 triple-positive were predicted to have the worst survival prognosis. However, due to differences in research design, sample size, and target population, the functions and mechanisms of CD133 in various aspects of oral CSCs are still not clear, and further investigation is necessary.
CD177: CD117, also known as the c-kit receptor, is a transmembrane protein with tyrosine kinase activity that plays a pivotal role in the maintenance and proliferation of hematopoietic stem cells (106). CD117 depletion in the bone marrow or spleen leads to a significant decrease in the number of cells from the erythrocyte and lymphocyte lineages (107). The CD117 signaling cascade is activated when CD117 binds to its cognate ligand, the stem cell factor (SCF), leading to the activation of the MAPK cascade (cRaf/Mek/Erk), the JAK/STAT pathway, and PI3K signaling events (108). The signaling pathways stimulated by CD117 are involved in various important cellular functions, such as self-renewal, apoptosis/survival, differentiation, and cell migration (109). CD117-positive cells were observed in the bone marrow, lung, spleen, and thymus (110). Previous studies have shown that CD117 is overexpressed in a number of solid cancer types, such as breast cancers (111), gastrointestinal stromal tumors (112), germ cell tumors (113), salivary gland tumors (114), and small cell lung cancer (115). Ongkeko et al. (116) analyzed the expression levels of CD117 in 44 patients with primary oral squamous cell carcinoma. Although its expression did not correlate with disease-free survival, the expression of CD117 was significantly higher in oral squamous cell carcinoma of the pharynx than in laryngeal cancers. Tsai et al. (117) demonstrated that chemoresis-tance to cisplatin was possibly correlated with the upregulation of CD117 and the ABC transporter gene (ABCG2) in oral cancer stem-like cells. However, experimental evidence regarding the reactivity of oral squamous cell carcinoma to CD117 expression is limited and contradictory. While Mărgăritescu et al. (118) and Galbiatti-Dias et al. (119) used CD117 with other stemness-related genes to identify and isolate oral cancer stem-like cells from oral squamous cell carcinomas, Barth et al. (120) analyzed CD117 expression in squamous cell carcinomas of the oral cavity, pharynx, and larynx and found that its expression was restricted only to stromal spindle cells.
Pluripotency-associated signaling pathway targeting strategy
The most frequently used strategy for the isolation, characterization, and targeting of CSCs within the tumor mass is based on specific cell surface biomarkers, such as CD133, CD44, and CD24 (34, 68, 121). However, these pluripotency-associated markers are not uniquely specific for CSCs, and they frequently overlap with their tissue-resident stem cells as well as normal somatic cells in many organs (122). Therefore, the use of single cell surface markers is considered under debate for selectively targeting CSCs within the tumor mass, because many studies found no cells with their specific characteristics (123). Alternatively, CSCs have also been identified and eliminated by selectively targeting key CSC-specific signaling pathways involved in their maintenance and various tumorigenic functions. Although the relationship between certain signaling pathways and several CSC functions still needs to be fully characterized, some of them appear useful for developing CSC-specific therapeutic strategies that avoid the deleterious side effects caused by affecting normal somatic cells or tissue-resident stem cells.
Wnt/β-catenin signaling pathway: Wnt proteins are a group of secreted glycoproteins that bind to cell surface receptors, including their cognate Frizzled (Fzd) receptors, and cause an accumulation of β-catenin in the cytoplasm (124). It regulates numerous processes essential for embryogenesis, tissue homeostasis, and cancer development (41, 125). Wnt/β-Catenin-mediated signaling, which is implicated in controlling various aspects of CSC-related tumorigenicity, has also been found to be involved in the maintenance, survival, metastasis, and drug resistance of various CSC types, including acute myeloid leukemia (126), breast (127), colon (128), liver (129), and lung (130) cancer. Iwai et al. (131) suggests that the Wnt/β-catenin signaling pathway plays a critical role in the oncogenesis of oral squamous cell carcinoma. They also demonstrated that the aberrant cytoplasmic accumulation of β-catenin can enhance the invasion and migration of oral squamous cell carcinoma by upregulating MMP-7 expression and inducing epithelial-mesenchymal transition (EMT) (131). Consistently, Yang et al. (132) reported that introducing a β-catenin gene into oral squamous cell carcinoma cells using a retrovirus vector-mediated transfection system resulted in a significantly increased resistance to TNF-α-induced apoptosis in transfected cells. Warrier et al. (133) also clearly demonstrated that Wnt/β-catenin signaling can increase the in vitro sphere-forming capacity and drug resistance of oral CSCs. They also reported that oral CSCs from squamous cell carcinoma are chemosensitized by naturally occurring Wnt inhibitors (sFRP4), via increasing apoptosis and/or reducing pluripotency (133). All-trans-retinoic acid (ATRA) reduced the self-renewal capacity of oral CSCs in vitro and subsequent tumorigenicity in vivo by inhibiting Wnt/β-catenin signaling from squamous cell carcinoma (134).
Notch signaling pathways: Notch signaling is an evolutionarily conserved intercellular signaling pathway that regulates various aspects of development and disease progression (135, 136). Therefore, dysfunction frequently results in a variety of congenital anomalies and diverse pathological disorders (41). The Notch receptors are four conserved single-pass transmembrane proteins (Notch1-4) and contain multiple arranged epidermal growth factor-like repeats (EGFR) that bind to Notch ligands (137). Among their family members, Notch 1 and 2 share the highest degree of similarity and are the most widely expressed in a large number of fetal and adult tissues, while Notch 3 is primarily limited to vascular smooth muscle cells, and Notch 4 is most predominantly expressed in endothelial cells (138). While the pro-oncogenic functions of dysregulated Notch signaling have been relatively well characterized in multiple types of cancers (138-140), its role in many aspects of CSCs is just emerging. Notch1 mutations are found in approximately 10%∼15% of patients with oral squamous cell carcinoma, suggesting its possible roles in the unique biological features of oral CSCs, such as long-term reconstitution, self-renewal, and transdifferentiation capacity (141-143). Shrivastava et al. (144) demonstrated that Notch1 was highly expressed in oral squamous cell carcinoma-derived 3D sphere-forming cells compared to monolayer cells. Consistently, they also observed the activation of Hes1, a well-known target of Notch signaling, in oral squamous cell carcinoma-derived sphere-forming cells as compared to adjacent monolayer cells (144). Moreover, Lee et al. (145) found that prolonged exposure to tumor necrosis factor alpha (TNF-α), a major proinflammatory cytokine, significantly enhanced multiple oral CSC-associated characteristics such as self-renewal capacity, pluripotency-associated genes, drug resistance, and tumorigenic potential in vivo by activating the Notch-Hes1 signaling cascade. Zou et al. (146) also reported that Notch2 expression was markedly increased in ALDH-positive CSC-like subpopulations in tongue squamous cell carcinoma. These results suggest that activation of the Notch signaling pathway can be mechanistically associated with the various characteristics of oral CSCs.
Hh signaling pathways: Hedgehog signaling was initially discovered as a critical segment polarity gene of pattern formation during early embryonic development in Drosophila, and its dysfunction frequently results in critical developmental anomalies and diverse pathological disorders (147, 148). It plays an essential role in regulating diverse cellular functions such as cell growth, survival/apoptosis, cell migration/invasion, and embryonic cell differentiation (149-152). While only one Hh gene has been identified in Drosophila, three different Hh family members have been found in vertebrates: the Desert Hedgehog (DHh), Indian Hedgehog (IHh), and Sonic Hedgehog (SHh) (153-155). Upon binding to its cognate receptor Patched-1 (a 12-pass transmembrane glycopro-tein), Hh initiates signal transduction through the transcription factor Gli, either dependently or independently (156). Many previous studies on several different types of human cancer, such as breast cancer (157), chronic myeloid leukemia (158), colorectal cancer (159), glioblastoma (160), lung cancer (161), multiple myeloma (162), and pancreatic cancer (163), have indicated that the Hh signaling network can possibly increase the self-renewal capacity, drug resistance, and tumorigenic potential of various CSC types. Consistently, Takebe et al. (164) provided a comprehensive survey of the Hh signaling pathway as a major regulator of many fundamental functions of CSCs, such as drug resistance, tumorigenic potential, pluripotency, and self-renewal capacity. Wang et al. (165) demonstrated that Shh is highly expressed in approximately 70% of oral squamous cell carcinoma specimens. In addition, statistical analyses showed that Shh overexpression is highly associated with the enhanced expression of the well-known Hh target gene Gli-1 and Hh receptor Ptch, suggesting that Hedgehog signaling is likely activated in oral squamous cell carcinoma cells (165). Consistently, Ptch expression is significantly associated with recurrence rates in patients with oral squamous cell carcinoma (165). Therefore, targeting dysregulated Hh signaling activity may provide effective diagnostic and therapeutic strategies for the treatment of oral cancers.
Hippo-YAP signaling pathways: The Hippo-YAP signaling cascades have been found as important oncogenic signaling pathways in various types of cancers (166). Its signaling activity is regulated by key downstream transcription co-activator Yes-associated protein 1 (YAP1) with PDZ-binding motif (TAZ) (167). In addition, YAP1 is essential transcription factor for maintaining undifferentiated state of embryonic stem cells by increasing the expression levels of various pluripotency-associated factors (168, 169). Importantly, significantly deregulated Hippo-YAP signaling pathway is widely observed in multiple types of human cancers including oral squamous cell carcinoma (170, 171). Indeed, local amplification of 11q22 locus in the human YAP1 gene is observed in 8.6% of patients with head-and-neck squamous cell carcinoma (172) and it signaling activity is closely related with poor prognosis and malignant phenotypes both in vitro and in vivo (172, 173). Endogenous YAP1 hyperactivation promotes surprisingly rapid and highly reproducible tumorigenesis of head-and-neck squamous cell carcinoma (174). Importantly, Li et al. (175) found that Hippo-TAZ signaling cascades enriched in CSCs subpopulation (CD44+/CD133+) by promoting the expression of pluripotency-associated transcription factor SOX2 in head neck squamous cell carci-noma. Similarly, several lines of evidence suggested that YAP1 could transcriptionally induce the expression of reprogramming factor SOX2 through a physical interaction with pluripotency-associated gene OCT4 to promote self-renewal capacity of CSCs subpopulation in lung cancer model (176).
Go to :
Oral CSCs and Their Potential Clinical Implications
Since the identification of CSC subpopulations in oral squamous cell carcinoma (22), a number of studies have described the positive correlation between the presence of oral CSCs and poor clinical outcomes (177, 178). Chen et al. (179) showed that the relative expression levels of ALDH1 and CD44 were significantly higher in high-grade oral squamous cell carcinoma. They also found that cucurbitacin I (JSI-124) can effectively induce the apoptosis of CD44+/ALDH1+ oral squamous cell carcinoma subpopulations through STAT3 signaling pathways (179). Likewise, Chen et al. (27) also demonstrated that ALDH1-positive cell subpopulations from oral squamous cell carcinoma have higher tumorigenic potential and are more resistant to chemotherapeutic agents and radiation than adjacent ALDH1-negative cell subpopulations. In this context, chemotherapy and radiation, the most commonly used therapeutic strategies together with surgery, often fail, as they do not effectively eliminate quiescent oral CSCs, which can reconstitute the entire bulk tumor. Therefore, it appears that establishing new adjuvant therapeutic approaches that can effectively eliminate oral CSCs within the tumor mass may provide a more effective treatment strategy to overcome therapeutic resistance and subsequent recurrence. Identifying highly selective oral CSC surface markers, as well as establishing effective therapeutic strategies, still requires intensive investigation (180, 181). Currently, therapeutic attempts to selectively target oral CSCs have not yet been used in clinical applications. Moreover, various pluripotency-associated CSC surface markers are not exclusively unique for oral CSCs, and they overlap with their tissue-resident stem cells as well as normal somatic cells (182). Therefore, a better understanding of CSC characteristics and the development of novel therapeutic approaches that selectively target oral CSCs are urgently needed to increase the clinical outcome in patients with different types of malignan-cies.
Go to :
Conclusions
After the identification of CSCs from leukemia over 40 years ago, they were first isolated and characterized from solid breast cancer tumors. CSCs not only play an essential role in cancer initiation, maintenance, and tumor progression, but they are also particularly important for mediating resistance to chemotherapeutic drugs and radiation, subsequently leading to the failure of these conventional therapeutic approaches. In this context, identifying and selectively targeting CSCs with specific pluripotency-associated signaling pathways and/or surface markers is a potential therapeutic strategy for inhibiting various types of cancer. However, no single cell surface marker that can specifically target the oral CSC subpopulation is currently available. The identification of specific surface markers or their signaling regulators is the first step in uncovering the characteristics and functions of oral CSCs. Indeed, combinations of a set of putative cell surface markers and/or signaling pathways representing the pluripotency-associated phenotypes will be helpful in achieving a better chance of developing new therapeutic alternatives for patients with oral cancer.
Go to :
 XML Download
XML Download