

 PDF
PDF Citation
Citation Print
Print


Introduction
Malignancies and cancer diseases account for a quarter of human death cases (1). The typical therapeutics for chances, including surgery, chemotherapy, and radiotherapy have not been able to be satisfying in treating the cancer patients. Although the methods for treating the malignancies and curing the cancer patients have greatly been improved in last years, in most of the cases there is not a proper response to the traditional therapeutics by the cancer cells. The tumor specificity is regarded to be the most important factor in limiting the efficacy of traditional cancer medications. Hence, it seems necessary for the medicine to search for the most efficient treatment approaches specifically targeting malignancies (2).
Mesenchymal stem cells (MSCs) are considered as the primary option for obtaining the stem cells for using in clinical and experimental applications. These cells have been obtained from various tissues, including brain, heart, and kidney suggesting a potential of promising candidates for several human diseases (3-7). MSCs are able to differentiate toward different cells and can simply be expanded in vitro, and therefore, have attracted the interests for utilizing in treatment options of that human diseases. They have the ability for self-renewal into mesenchymal lineages. MSCs exhibit pathotropic migratory features, establish them as potential candidates for selective delivering the drugs with the aim of tumor therapy (3, 4).
Tumor cells applies angiogenesis to be survived and proliferated (8). A number of growth factors and extracellular matrix proteins are responsible for tumor angiogenesis and, therefore, researchers have been attracted to utilize the targeted anti-angiogenic therapy of treatment of tumors (9, 10). Besides, several stem cells have been modified in vitro for expressing anti-angiogenic factors more efficiently (3). In this review article, we try to clarify the possible applications of MSCs through anti-angiogenic properties to treat cancer patients.
Go to : 
Characteristics of MSCs
MSCs are considered as the adult stem cells and naturally generated in the human body. MSCs were initially found in the bone marrow (BM) stromal matrix but they are distributed throughout the body (11, 12). MSCs are commonly found in several fetal and adult organs, like amniotic fluid, heart, skeletal muscle, adipose tissue, synovial tissue, placenta, pancreas. According to evidence, organs and tissues containing connective tissue have also MSCs in themselves (13). MSCs are considered as the primitive cells that are basically originating from the mesodermal germ layer and have been regarded as progenitor cells that have the potential to be developed to the connective tissues, skeletal muscle and vascular cells. MSCs have also the potential to be differentiated into cells of the mesodermal lineage, like fat, bone, and cartilage cells, but they can also differentiate into neuroectodermic and endodermic cells (14). MSCs have been considered as an important source in biomedicine due to the multilineage capacity (15). Because of an ease in acquisition, fast proliferative capacity, and the autologous transplantation potential, MSCs have been regarded as the first option of stem cells to be utilized in the regenerative medicine. MSCs may confer advantageous capacity for cell recovery in the harmed tissues (16). These stem cells has been reported to be involved in the modulation of immune cells and, hence, are attributed to be contributing factor in autoimmune disorders (17, 18). Studies have established a tumor specific migration and residence feature for MSCs, suggesting a positive capacity of these cells to be utilized as promising carriers of drugs for the aim of cancer therapy (19). Both pro- and anti-cancer characteristics have been attributed to MSCs (20); nonetheless, if MSCs are monitored efficiently, for example with anticancer agents, they could be applied in cancer therapy.
Go to :
Sources of MSCs
Several sources of adult tissues have been identified for MSCs extraction and the harvesting of MSCs is not subject to the ethical issues (3). MSCs are potentially able to develop to various tissue types that might be either within or across the germ lines (21). BM-derived MSCs have been attributed with the highest level of lineage plasticity, that can be differentiated to the almost all cell types (22, 23). That notwithstanding, the available data obtained from the preclinical studies have indicated that the BM-derived MSC may not be the best source for utilization in the clinic. Invasive procedures are used for the harvesting of BM that yields a little number of cells. On the other side, the number, differentiation capacity, and life time of BM-derived MSC are decreased as the individuals become older (24, 25). Adipose tissue and umbilical cord blood are considered as two alternative sources to harvest MSCs. Recently, MSCs obtained from adipose tissue have gained high attention to be used in preclinical and clinical evaluations, since tissue sampling is simple, the initial cell numbers is high, and the proliferation potential is satisfactory (26). However, phenotypes of MSCs, and the expansion and differentiation capacity of these cells originating from the adipose tissue are similar to those obtained from BM (27).
Furthermore, the umbilical cord blood as well as the wharton’s jelly have been implied to have a plenty of MSCs (28). The cells extracted from placenta in adherent layer, which is considered a noninvasive and simple approach, have been reported to have a fibroblastiod morphology, which represent similar surface molecules as BM-derived MSCs, such as CD90, CD13, CD49e, CD29, and CD54 (29). Umbilical cord blood-derived MSCs show the ability to proliferate at a higher rate in comparison to the BM and adipose tissue-derived MSCs (27, 30), which is probably because of high telomerase activity in the umbilical cord blood-derived MSCs (31).
Go to :
Circulation and Homing of MSCs
As the first step in the homing, MSC needs to migrate from the BM to the circulation and move toward other tissues (Fig. 1). It has been indicated that MSCs can mobilize from the BM as well as other tissues to the circulation upon the various injury situations, including hypoxia, normoxia, and inflammation (32, 33). It is not completely divulged that how MSCs migrate from the BM and cross through the endothelium and how home in the tissues. That notwithstanding, it is clear that a common role of MSCs is to home in and repair the injured organs. The wound healing ability of MSCs initiates with their migration to the signals released from the injured organs (34). Various inflammatory factors released from wounds have also been identified in the tumor microenvironment that have been attributed to the migration of MSCs (35). Among the mediators/receptors involved in the migration of MSCs are MCP-1/CCR2, SDF-1/CXCR4, HGF/c-Met, SCF-c-Kit, VEGF/VEGFR, PDGF/PDGFR, and HMGB1/RAGE (36). The involvement of the stromal cell-derived factor SDF-1 and the related receptor, CXC chemokine receptor-4 (CXCR4), in the migration of MSCs identified in surveys that either the receptor or the cytokine was knocked down (37-39). Furthermore, it was demonstrated that inhibition of CXCR4 and SDF-1 in mice cause marked decrease in the migration of exogenous stem cells to target organs (37). It has also been indicated that the blockade of CXCR4 resulted in an impaired homing of endogenous MSC into tumor sites, development of MSCs into myofibroblasts, and reduced MSC survival (40). It has been revealed that chemokines like CXCL12, CXCL13, CXCL16, and their receptors play a role in the mutual mobilization of MSCs to BM and generate the BM niche. In the efficient homing of MSC into the BM, CXCL16 play an important role. CCL22 has been shown to play the strongest chemotactic impression in the migration of MSCs from the BM into the blood (41). Tumor tissues represent CCL2 and CXCL16, suggesting that they are playing a vital role in the mobilization and migration of MSCs into tumor sites (42-44).
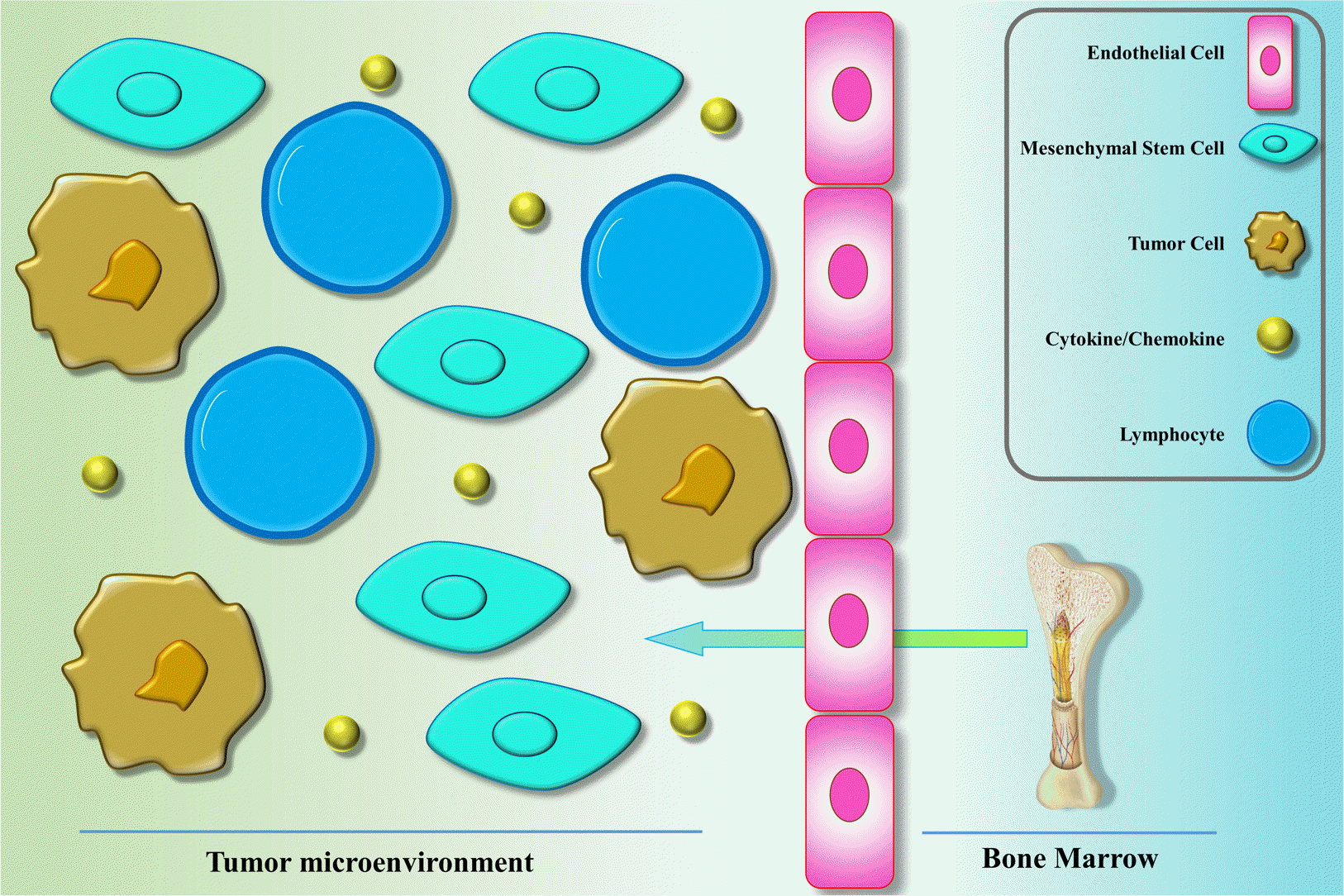
Matrix metalloproteinase (MMPs) enzymes poses proteinases function, which is needed for proteolytical cleavage of precursor proteins, such as adhesion molecules, growth factors, cytokines, as well as some receptors. It has been demonstrated that MMP-1 and tissue inhibitor of metalloproteinase-3 (TIMP3) play role in the migration of MSCs across the BM endothelium (45). Plus, increased MMP-2 levels in the serum was attributed to the C1q complement -mediated migration of umbilical cord blood-derived MSC into the wounded tissue (46).
It was demonstrated that MSCs were recruited to the regions of irradiation, and local irradiation may increase the MSC specificity to be homed in particular tissues (47). These observations about the migratory characteristics of MSCs testify the promising potentials for designing the therapeutic tools that exploit the tumoritropic features of MSCs through engineering these cells towards potent cells via delivering the compounds against tumors.
Go to :
The Behavior of MSCs in the Tumor Microenvironment
MSCs show different kinds of activities in various microenvironments because of the diversity in the signaling pathways involved in the stimulation of MSCs. In the clinical studies, employed MSCs are primarily naïve, obtained from normal tissues, and are usually harvested in vitro. Such naïve MSCs have the ability to interfere with the tumor cell growth in co-cultures with tumor cells. It was observed that naïve MSCs could decrease the expansion of leukemia cells. Such blocking property of naïve MSCs was seen to be dose-dependent, as the naïve MSCs was increased, the rate of inhibition of tumor growth was increased. It was suggested that naïve MSCs might decrease the expansion of tumor cells through mediators released from MSCs, such as Dickkopf-related protein 1 (DKK1), an inhibitor of Wnt signaling pathway in tumor cells (48). Interfering in the Wnt pathway was associated with the decreased expression of Cyclin D2 and c-Myc as well as increased expression of P27KIP1 and P21CIP1, that culminated in the interruption of tumor cell cycle (49-52). On the other side, naïve MSCs have the potential to activate the apoptosis in tumor cells (53) via increasing the expression of caspase 3 (50). Furthermore, naïve MSCs have the potential to decrease the expansion of tumor cells via inhibiting the angiogenesis. In this way, naïve MSCs increase the apoptosis rate in vascular endothelial cells and interfere with angiogenesis (54, 55).
On the other side, tumor supporting activities of MSCs have also been reported (56). An equilibrium between anti-inflammatory and pro-inflammatory phenotypes of MSCs determine the influence of MSCs on the progression or suppression of tumor cells (57). It has been shown that MSCs inside tumors might undergo functional modulations to alter from an anti-tumorigenic phenotype to pro-tumorigenic MSC (58, 59). MSCs with anti-inflammatory properties are involved in contributing to tumor development by several mechanisms. Tumor perturbations has been shown to be associated with recruitment of MSCs to tumor sites (60). MSCs are able to regress immune responses against tumor cells (61), induce angiogenesis in tumors (62, 63), promote epithelial-to-mesenchymal transition (EMT) resulting in metastasis (64, 65), and induce a resistance in tumor cells to different therapeutics (66). Furthermore, it has been indicated that MSCs promote tumor progression through modulating the metabolic settings of tumor cells. Particularly, prostaglandin E2 secreted by MSCs is able to abrogate the apoptosis of lymphoblastic leukemic cells (67). MSCs produce lactate in a tumor microenvironment with high oxidative stress conditions that is uptaken via tumor cells, resulting in promoted migratory ability of tumor cells (68). Naïve MSCs have also been attributed with the increased angiogenesis in the colon cancer cell lines (69, 70), differentiation of MSCs to vascular endothelial cells in melanoma (71), increasing the development of cancer stem cells (CSCs) that promote the tumorigenesis and metastasis (72) in breast cancer (73), enhanced proliferation of gastric cancer cell lines (72, 74). Moreover, naïve MSCs might support tumor progression through increasing the migratory abilities of tumor cells via production of chemokines, like CXCR4 (75), CCL5 (76, 77), intercellular adhesion molecules (ICAMs), and vascular cell adhesion molecules (VCAMs) (78). Moreover, by suppressing the immune responses, MSCs has been observed to enhance the tumor development (79).
In addition, MSCs generate a wide range of chemokines, cytokines, and growth factors that mediate paracrine- or autocrine functions in the tumor development (80). These factors are able to modulate the tumor microenvironment to a tumor supporting settings. Several inflammatory mediators secreted by MSCs, including CXCL1, CXCL2 or CXCL12, are able to indirectly enhance tumor development in several tumor models (81, 82). In the same way, MSCs-derived inflammatory chemokines and cytokines, such as IL-8 and IL-6, contribute the tumor development in colon cancer (83) and breast cancer (73) models. By a paracrine mechanism via IL-6 and CXCL7 secretion, MSCs was shown to migrate into breast cancer xenografts and promoted the development of CSC subpopulations (73).
It seems that MSCs may represent bidirectional behaviors on tumor development, and various studies have reported a disagreement on the MSCs effect on the tumor progression or inhibition. However, it is worthy to mention that there is a general agreement that MSCs are frequently contribute to the tumor development in comparison to suppressing tumor growth, which is widely attributed to the regenerative functions and immune system suppression by MSCs (84, 85).
Go to :
MSC-Derived Extracellular Vesicles
Extracellular Vesicles (EVs), like exosomes, are membrane-enclosed vesicle with a size of 40∼1,000 nm that are produced by several cells, such as MSCs (86). EVs primarily encompass microRNAs (miRNAs) as well as proteins, which is encapsulated by a lipid bilayer membrane (87-90). Secreted EVs are able to move toward other cells and bind them in order to deliver several signals to the targets (91). MSC-derived EVs are able to modulate several processes in the tumor cells, including metastasis, angiogenesis, and proliferation (92) as well as control immune cells (93, 94). Indeed, MSC-derived EVs might promote the tumor development or repress tumor progression (95).
Umbilical cord derived MSC-EVs was shown to meliorate the oxidative stress and cell apoptosis as well as promote cell proliferation in rats with cisplatin-induced nephrotoxicity (96). Additionally, Zhu et al. (97) indicated that MSC-derived EVs promoted tumor growth through promoting the expression of vascular endothelial growth factor (VEGF) in tumor cells by induction of the extracellular signal-regulated kinase1/2 (ERK1/2) pathway. Therefore, MSCs are able to promote tumor growth and angiogenesis by secreting EVs.
On the other hand, it was shown that BM-derived MSCs-EV caused cell-cycle arrest in the G0/G1 phase and induced apoptosis in tumor cells (98). Additionally, MSCs-EV interrupted the angiogenesis by suppressing the expression of VEGF in tumor cells (99). Furthermore, MSC-derived EVs containing miRNAs targeting VEGF were observed to suppress angiogenesis in human nasopharyngeal carcinoma cells (100). miR-143, which has been investigated in several malignancies (101), was indicated to be harbored by MSC-derived EVs an reduced the migration of osteosarcoma cells (102). As a consequence, MSC-derived EVs are able to repress the tumor cell proliferation and angiogenesis.
Go to :
Exertion of MSCs for Treatment of Tumor
Naïve MSCs (have not undergone any manipulation) have been reported to possess antitumor properties. This antitumor activity has been attributed to the mediators produced by MSC that have the potential to decrease the proliferation of tumor cells in different cancers, including breast adenocarcinoma, melanoma, hepatoma, lung cancer, and glioma (52, 103-105). In a mouse model of Kaposi’s sarcoma, the BM-derived MSCs were administered intravenously that resulted in the homing of these cells in the tumor sites and prevented the progression of tumor (106). Studies in vitro and in the mouse model of melanoma indicated that MSCs possess the ability to repress the angiogenesis (55). Furthermore, MSC administration to the melanoma mice model subcutaneously eventuated in increased apoptosis rate and decreased expansion of tumor (55). In addition, umbilical cord blood-derived MSCs overexpressing CD44 and CD133 were reported to increase apoptosis rate in the glioblastoma multiforme cells in vitro (107). As well, umbilical cord blood-derived MSCs administration resulted in underexpression of XIAP activating caspase-3 and caspase-9, leading to increased apoptosis rate in the glioma cells (108). To comply with these observations, umbilical cord blood-derived MSCs as well as MSCs from other sources indicated reduced expansion of the glioblastoma multiforme cells through tumor necrosis factor (TNF)-related apoptosis-inducing ligand (TRAIL) (109).
Go to :
Modulation of Angiogenesis for the Treatment of Cancer
The process of angiogenesis
In the embryonic development period, neovascularization or new blood vessel formation from pre-existing vessels is regarded as a major occurrence that leads to the construction of vasculature system that contains endothelial cells (ECs) (110). During the physiological situations, vasculogenesis is seldomly observed in the adults. That notwithstanding, angiogenesis may be seen in adults as a physiological event during pregnancy as well as the cyclical growth of vessels in the ovarian corpus luteum (111, 112). In the pathological conditions, such as tissue repair during wound healing and tumor growth, neovascula-rization is considered as a normal phenomenon (113). However, it should be noted that the structure and composition of the tumor vessels are different from those occurred in the normal vessels. It has been shown that ECs of tumor tissues express higher levels of CD109, CD137, CD276, and placenta growth factor (PlGF) compared with the ECs originating from the normal non-tumor organs. In spite of a bulk of studies with respect to the cellular and molecular characteristics of ECs from different tumors, there are disagreements with respect to the origin of ECs in the tumor conditions (114). It has been reported that endothelial progenitor cells (EPCs) are actively involved in the angiogenesis process of tumors (114). However, it is clear that the initiation of angiogenesis requires a divergence toward a profile of increased angiogenesis activators, while inhibited process of angiogenesis suppression. The critical stimulating mediators of angiogenesis include vascular endothelial growth factor (VEGF)-A, fibroblast growth factor (FGF), and hepatocyte growth factor (HGF), PlGF, and MMPs (115, 116). Adversely, thrombospondins, endostatin, angiostatin, and interleukin (IL)-12 are important endogenous inhibitors of angiogenesis (117). Taken together, angiogenesis is thought to be involved in the progression and development of tumors.
Anti-angiogenesis tumor therapy via interfering with VEGF pathway
Noting to the participation of the angiogenesis process in the development of tumors, it seems that the molecules involved in the tumor angiogenesis can be targeted as a therapeutic strategy. Inhibition or blocking of the growth factors or signaling pathways prerequisite for the development and progression of ECs is regarded as a logical mindset for prevention of tumor vasculogenesis and, therefore, tumor repression (113, 118, 119). VEGF is considered as an critical angiogenetic factor which is involved in both physiological and pathological situations (120). Genetic alterations in the VEGF gene have been reported to cause defects in the normal development angiogenesis and death in the embryonic period (120). Hypoxia inducible factor (HIF)-1α, through binding to the VEGF promoter, regulates the expression of VEGF (121). A hypoxic condition in various regions of tumors results in the increased expression of VEGF in tumor niche, leading to an enhanced blood supply and expedited expansion of tumor cells (122). Furthermore, VEGF is normally overexpressed in the microenvironment of numerous human tumors (123); however, VEGF receptors of VEGFR1, VEGFR2, and VEGFR3 have been shown to be highly expressed in the tumor associated ECs (124, 125).
The tumor mice model were administered with an monoclonal antibody targeting the VEGF that manifested as a remarkable deceleration in the tumor growth (126). Bevacizumab is a humanized monoclonal antibody to neutralize VEGF (127), was the first anti-angiogenic monoclonal antibody approved by the FDA (Food & Drug Administration) in 2003 for the therapy of metastatic colorectal cancer patients (128, 129). In addition, the promising effects of bevacizumab in treating patients with non-small cell lung cancer (NSCLC) (130) and metastatic breast cancer (131) were established. Upon these successful experiences, some other factors inhibiting the VEGF pathway, target either VEGF or its receptor, have been under clinical trial studies. As a soluble receptor of VEGF, VEGF-TrapR1R2, which carries the functional parts of the VEGFR1 and VEGFR2 (132), has been shown to have the ability to neutralize the circulating VEGF and be utilized in tumor therapy. In addition, it has been observed that VEGF-TrapR1R2 possesses better anti-tumor effects in relation to DC101, which is a VEGF receptor blocker. Studies have shown that the inhibition of VEGF signaling pathway, through the components that blocks the VEGF receptor, are efficiently involved in the repression of the tumor progression (133, 134). Receptor tyrosine kinase inhibitors (RTKIs), which targets molecules in the VEGF signaling pathways, including linifanib (135, 136), cabozantinib (137), axitinib (138), tivozatinib (139), vendatanib (140), sunitinib (141), pazopanib (142), and sorafenib (143) has been shown to be promising in the treatment of various tumors. In spite of promising effects of these therapeutics in the clinical trials, the detailed mechanisms underlying their beneficial anti-angiogenic effects have not fully been disclosed. Preclinical models have revealed that the inhibition of angiogenesis has been the main mechanism of angiogenesis inhibitors that target VEGF pathway. That notwithstanding, vascular normalization has also been implicated as another mechanism of anti-angiogenetic agents. This mechanism imply to the targeting of the non-functional vessels in tumors, that implement positive functions through decreased blood flow to the tumor, or efficient drug delivery to the tumor cells (144). Therefore, progression in designing agents with potential targeting of VEGF pathway and, therefore suppressing the angiogenesis, provided prosperous therapeutic strategy for tumors that have the potential to be further improved in combination with other tools and approaches.
Vascular disrupting agents (VDAs) have been considered as another class of anti-angiogenetic compounds above and beyond the VEGF inhibitors. To implement their anti-angiogenic functions to prevent tumor development, VDAs cause vascular collapse, manifesting as hypoxia, which in turn cause tumor necrosis (145). ASA404 is a VDA that has been shown to cause apoptosis in the ECs, and, thereupon, diminished blood flow to the tumor cells. Clinical studies is currently assessing the efficacy of ASA404 in the NSCLC patients (146).
Go to :
MSCs and Anti-Angiogenic Cancer Therapy
Angiogenesis is applied by tumors to proceed the process of growth (8). Angiogenesis can be inhibited through the exertion of MSCs as vehicles to deliver anti-angiogenetic drugs to the tumors and, therefore, limit the tumor progression. Such engendered MSCs have displayed a tropism to cancer organs and can deliver anti-angiogenic drugs to the tumor sites, while having little adverse effects (147). However, it should be noted that a systemic supply of anti-angiogenic drugs for a long time may present with adverse effects, such as drug toxicity, as well as decreased blood supply of the tissues, which in turn results in the poor delivery of chemotherapeutic drugs to the target sites (148).
Dysregulated balance of pro-angiogenic and anti-angiogenic mediators as well as growth factors in the tumor microenvironment has been attributed to the tumor-mediated angiogenesis (9, 149). As an endogenous inhibitor of angiogenesis in the tumors, endostatin has been exerted as an important anti-angiogenic agent in several malignancies (150). In a study, the human placenta-derived MSCs were manipulated to deliver endostatin and were injected into nude mice. These MSCs, which expressed endostatin, were observed to be resided in the tumor site and a significant reduction in the tumor size was reported without considerable systemic toxicity and adverse effects. The reduced tumor size was reported to be due to increased apoptosis in the tumor cells and blocked angiogenesis (150). Furthermore, a phase II clinical trial reported that delivery of anti-angiogenic compounds resulted in the correction of the abnormal structure and malfunction of the blood vessels, culminating in a remarkable decrease in the brain edema (151). This correction in the vessel structure was reported to be due to decreased vessel diameter and permeability (152, 153). Intratumoral administration of MSCs has been shown to be accompanied with the residence of these cells in the tumor sites and interfering with vasculogenesis, implying to a beneficial potential of MSCs for targeted anti-angiogenic drug delivery (154). The genetically-modified human adipose-derived MSCs expressing the interferon γ-induced protein 10 kDa (IP-10), which is a powerful chemoattractant, in a mouse model of metastatic melanoma resulted in inhibited tumor cell growth and limited angiogenesis and significantly associated with increased survival of mice (155).
Go to :
The Road ahead toward the Delivery of Anti-Angiogenic Inhibitors via MSCs
Estrogen, a female sex hormone, has been shown to stimulate angiogenesis through acting directly on ECs, as well as indirectly on endometrial cells. It has also been reported that estrogen triggers the expression of VEGF in the uterine stromal cells (156, 157). ER has been established as a potential target of various drugs for inhibition of the breast cancer progression (158). The ligation of estrogen to the estrogen receptor (ER) causes an increased proliferation; thereupon, endocrine cancer therapy attempts to prevent the binding of estrogen to the ER and block the aftermath signaling. Tamoxifen and raloxifene are considered as selective estrogen receptor modulators (SERMs) that are competitive inhibitors of estrogen and directly bind to ER. Interfering with the estrogen pathway has been indicated as an efficient strategy for the therapy of hormone responsive breast cancers, while conferring little toxic side effects (159). High expression of ERβ has been observed in the prostate (160). Implying to the possible response of prostate cancer to SERMs (161). Moreover, promising outcomes have been established about the estrogen therapy in the ovarian cancer patients (162). As well, the Women’s Health Initiative (WHI) trial indicated that estrogen is involved in the colon carcinogenesis (163). As a result, with respect to the role of estrogen and ERs in the process of angiogenesis and the promising outcomes of estrogen/ER pathway blockade, it seems that MSCs can be exerted as machine for the targeted delivery of drugs that interfere with this pathway. Moreover, monoclonal antibodies targeting ERs can be evaluated to inhibit angiogenesis in the tumor microenvironment. SERMs have been attributed with a number of adverse effects in the tissues that are not the target of such compounds. Hence, strategies for targeting the estrogen/ER pathways needs to be optimized to obtain best results. This is a hypothesis by the authors, requiring studies in the animal models as well as clinical trials to be validated.
It has been demonstrated that progesterone prevents the proliferation of ECs in vitro and interferes with the cell cycle in the human dermal endothelial cells (164). Progesterone receptor is expressed on the endometrial ECs and may play a role in the suppression of VEGF-induced proliferation of ECs (165). Therefore, targeting the progesterone pathway through manipulated MSCs seems potential therapeutic strategy for cancer treatment, a hypothesis that needs to be extensively studied.
The corpus luteum is considered as a temporary endocrine tissue in ovaries that are involved in the prosperous gestation of mammals. When a follicle undergone the rupture process upon ovulation, the corpus luteum is then converted to debris and undergoes a number of modifications, such as differentiation and growth, which rely on the angiogenesis process. It has been reported that some endogenous stimulatory and inhibitory compounds exist in the corpus luteum to regulate the angiogenesis (166). Follicular cells of ovaries produce the pigment epithelium–derived factor (PEDF), which is a physiological inhibitor of angiogenesis (167). PEDF possess an anti-angiogenic function in the corpus luteum, implying to its possible utility as an anti-angiogenesis therapy of cancer in the future, probably through delivering via MSCs as vehicles.
Go to :
Challenges in Application of MSC in Cancer Therapy
The therapeutic application of MSCs has been associated with multiple challenges that may limit the clinical efficacy of this approach. Among the drawbacks in exertion of MSCs are inappropriate homing of MSCs in the body and little accumulation of these cells in the target tumor (168). Additionally, MSCs may lose their therapeutic potential, gain immunogenicity properties, and obtain tumorigenic characteristics during physiological differentiation into mesenchymal lineages (169). It has also been shown that endogenous MSCs are able to produce several mediators and activate signaling pathways to promote the angiogenesis in tumor cells, like colorectal cancer (69) and melanoma (170). However, a study analyzed clinical 36 trials that used BM-derived MSCs through intravascular delivery in different subjects, including graft versus host disease, Crohn’s disease, ischemic stroke, cardiomyopathy, myocardial infarction, and healthy volunteers. This study indicated no associationship between the MSCs therapy and tumorigenic risk. Moreover, no severe adverse effects of the MSC therapy were observed (171). Most of the clinical trials have evaluated the safety and tolerability of the MSC therapy in cancers. Nonetheless, the adverse effects of MSC therapy in promotion of tumor, instead of suppressing it, has little been studied. The tumorigenic evaluation of MSC therapy needs long time follow-up of the patients, as carcinogenesis is a gradual process.
Go to :
Concluding Remarks
Several beneficial properties have been proposed for MSCs that make them appropriate candidates for cell-based therapeutic tool in cancer therapy. First, MSCs are able to mobilize and preferentially home in tumor tissues and interact with various cells present in tumor microenvironment. Furthermore, MSCs are easily available, have no or little immunogenic features, can be simply manipulated in vitro. Little have been surveyed with respect to the preclinical evaluation of the MSC potential to be exerted as vehicles to locally deliver/express a single therapeutic compound in cancers. Despite a promising experience in the promoted survival of cancer cases through anti-angiogenic therapy, the pooled findings testify a limited overall survival improvement (172, 173). These limitations stem from several issues. First, tumor microenvironment may exert mechanisms to resist against anti-angiogenetic molecules and, therefore, limit the efficacy of such therapeutics (174). Second, anti-angiogenetic drugs may develop vascular collapse that may result in a hypoxic state, which in turn may result in radio-resistance, chemo-resistance and anti-angiogenesis resistance (175, 176). These adverse effects may culminate in tumor metastasis and, therefore, negatively impress the clinical exertion of anti-angiogenic drugs. In spite of an improvement in our knowledge of MSCs in recent years, little has been attempted with respect to the employment of these cells to downmodulate the angiogenesis in the clinics. Therefore, filling this gape requires developing state-of-the-art technologies and new concepts in exertion of anti-angiogenic compounds conferring little side effects. To attain this end, engineering MSCs to specifically deliver the angiogenesis inhibitors could be still promising, particularly when combined with other therapeutics and targeting other pathways simultaneously. As well, searching for novel molecular targets to diminish the angiogenesis could be a part of answer to this gap.
Go to :
 XML Download
XML Download