

 PDF
PDF Citation
Citation Print
Print


Introduction
Myocardial infarction (MI) emerges as a major cause of physical disability and even mortality since it induces clinical manifestation like vomiting, continuous chest pain, nausea, depression, fatigue, feebleness, arrhythmias and tachypnea (1, 2). MI is strongly associated with the perilous factors embracing high lipid concentration, diabetes or hypertension, lack of physical exercise, unbalanced diet, overweight and smoking, which aggregate intravascular clots, eliminate oxygenation, disrupt vascular endothelium construction and aggravate atherosclerosis in MI patients (3). Furthermore, MI can trigger undesirable consequences, for example, myocardial failure, high rate of relapse and mortality (4). To avoid those poor prognostic responses, candidate treatment that targets MI severity is urgently required (1).
Hence, treatment of cells, especially mesenchymal stem cells (MSCs) attracts increasing attention in myocardial repair by their high differentiation and self-renewal ability and features of angiogenesis and immunomodulation (5). Besides, MSCs could practically strengthen heart function, decrease scar range and promote myocardial repair through suppressing cardiac fibrosis, mediating myocardial cellular behaviors and modulating immune responses (6). Significantly, when adipose-derived SCs (ADSCs) are introduced into MI area, myocardial self-renewal and repair are promoted (7). Moreover, compared with the other MSCs, ADSCs could better reduce MI range and scar size (8). However, although the administration of MSC contributes a lot to heart diseases, the survival rate of these cells is still a disappointment (9). In other word, to facilitate viability and migration of ADSCs might be a breakthrough in post-MI alleviation and repair.
A number of studies have suggested that the modification of microRNAs (miRs) to MSCs prevents cells from death or autophagy, restricting MI size and improving myocardial repair of MI tissues (10, 11). miRs are deemed as reliable biomarkers of MI as they regulate downstream genes or axis and affect myocardial function (12). miR-324 is downregulated in cardiomyocytes with hypoxia/reoxygenation-induced injury, inhibiting cell proliferation (13). However, research about the specific function of miR-324-5p in MI progression is elusive. Here, we found that miR-324-5p could promote the viability and motility of ADSCs, which is corroborated by a recent study that miR-324-5p encourages adipocyte differentiation and adipogenic level (14). What’s more, Chen et al. (15) have revealed that miR-324-5p protect myocardial tissues against MI injury by reducing mitochondrial fracture and oxidative stress damage, and enhancing endothelial progenitor cell viability. Besides, as a target gene of miR-324-5p, Toll-interacting protein (TOLLIP) is overexpressed in MI tissues and it degenerates inflammatory reaction of MI and spoils cardiac function (16). By means of these information, we hypothesize that miR-324-5p-modified ADSCs could greatly improve post-MI myocardial repair by targeting TOLLIP.
Materials and Methods
Experimental animal
Male Sprague-Dawley (SD) rats (4 weeks, 100∼110 g) (Beijing Vital River Laboratory Animal Technology Co., Ltd, Beijing, China, SYXK (Beijing) 2017-0033) were kept in a controlled environment with constant humidity (60%±5%), and temperature (22±3℃) for 12 h light/dark cycles with free access to food and water. All the operations were committed under anesthesia.
Separation and cultivation of ADSCs
ADSCs were separated and cultivated as previously reported (17). Briefly, inguinal adipose tissues of SD rats were separated and cleaned with phosphate buffer saline (PBS) to discard the residual blood. Then, tissues were cut into pieces (1×1 mm) which were detached with collagenase I. After the centrifugation at 4,000 g for 5 min, cell particles were suspended in Dulbecco’s modified Eagle’s medium (DMEM) consisting of 10% fetal bovine serum (FBS)/1% penicillin-streptomycin/2 mM L-glutamine at 37℃ for 48 h in a humidified atmosphere with 5% CO2. The medium consisting of non-adherent cells was refreshed by new medium every 3 days. The morphology of ADSCs was observed under a microscope. Cells were subcultured when they reached 90% confluence and experiments were performed using passaged 3 (P3) ADSCs.
Identification of ADSCs
Immunophenotyping of ADSCs was analyzed on a FACScalibur flow cytometer (BectonDickinson, Bedford, MA, USA) with CD90 (ab225), CD45 (ab6329), CD105 (ab2529), CD34 (ab81289) antibodies (all from Abcam Inc., Cambridge, MA, USA).
Subsequently, adipogenic and osteogenic differentiation assays were committed. Osteogenic differentiation went as: ADSCs at 80∼90% confluence were cultured in osteogenic induction medium, which included DMEM containing 10% FBS, ascorbic acid, β-glycerophosphate, and dexamethasone, from which ADSCs were cultured to induce osteogenic differentiation. Adipogenic differentiation was conducted as ADSCs were incubated in adipogenic induction medium, which included DMEM containing 10% FBS, insulin, indomethacin, dexamethasone, and 3-isobutyl-1-methylxanthine, from which ADSCs were cultivated to induce adipogenic differentiation. The medium was refreshed every 3∼4 days for 21 days. Subsequently, cells were fixed and stained with alizarin red or oil red O. All reagents were attained from Beijing Solarbio Science & Technology Co., Ltd. (Beijing, China).
Treatment of ADSCs
P3 ADSCs were incubated by DMEM with 10% FBS in the 6-well plates till 70% confluence. At this point, ADSCs were divided into the blank group (uninfected), the lentiviral vector (LV, Shanghai GenePharma Co, Ltd, Shanghai, China)-negative control (NC) group (infected with NC), and the LV-miR group (infected with miR-324-5p overexpression lentivirus). Then, the LV-NC group or the LV-miR-324-5p group was infected with 8 μg/ml polybrene (Solarbio) into ADSCs (18-20).
Cell counting kit-8 (CCK-8) method
Cells (5×103 cells/well) with various treatment were inoculated into 96-well plates at 37℃ for 24 h, 48 h and 72 h, respectively, with 10 μl CCK-8 reagent (Shanghai Beyotime Biotechnology Co. Ltd., Shanghai, China) supplemented into each well. Next, optical density at 450 nm was assessed using a microplate reader (Bio-Rad, Inc., Hercules, CA, USA).
Transwell assay
ADSCs were detached by trypsin, centrifuged and soaked twice in PBS. Afterwards, ADSCs were resus-pended in DMEM consisting of 10% FBS. Then, 100 μl cell suspension (1×104 cells) and 100 μl DMEM consisting of 10% FBS were supplemented into the 24-well plates, which were then placed in Transwell chamber and cultivated at 37℃ for 24 h. The Transwell chamber was subsequently removed, and the cells were immersed into PBS, fixed in 4% formic acid (Solarbio) for 10 min, and then stained with Hoechst 33258 (1 μg/ml, Solarbio) for 30 min in the dark. Finally, cells were rinsed 3 times in PBS, with the total number of migrated cells counted by a microscope.
Establishment and treatment of MI rat model
Rats were anesthetized with pentobarbital sodium (30 mg/kg, Sigma-Aldrich, Merck KGaA, Darmstadt, Germany) via intraperitoneal injection using a ventilator connected to trachea cannula. Heart was exposed in the fourth and fifth intercostal gap after left thoracotomy, when left anterior descending coronary artery (LAD) running in myocardial tissues consistent with the direction of the great myocardial vein was found, it was sutured and ligated with 6-0 silk thread. After LAD occlusion, the corresponding blood supply area was whitened, while infarction manifestations such as ST segment elevation was noticed as electrocardiogram displayed (11, 18, 20-23). Subsequent treatments were performed 30 min after ligation (22). Peri-MI areas were injected at multiple sites by PBS or 20 μl PBS containing 1×106 differently treated ADSCs or adenovirus (Ad)-NC (empty vector Ad, 0.5×109 pfu) or Ad-TOLLIP (TOLLIP Ad, 0.5×109 pfu) (Ad-NC and Ad-TOLLIP were both synthesized by Xinji Biotechnology Co., Ltd, Shanghai, China). The sham group received only thoracotomy without LAD. After ADSCs transplantation, all rats underwent echocardiography on the 28th day, and blood samples were collected, with 6 were used for reverse transcription quantitative polymerase chain reaction (RT-qPCR) and the remaining ones for hematoxylin and eosin (HE) staining and terminal deoxynucleotidyl transferase-mediated dUTP nick end labeling (TUNEL) staining.
Myocardial function assessment
Myocardial function of rats was evaluated using the Vevo2100 small animal ultrasound imaging system (VisualSonic, Toronto, Canada). In brief, rats were anesthetized with 40% oxygen (GH-reagent, Shanghai, China) and isoflurane (1.7%, final concentration), and an ultrasound probe was inserted into the short axis of the left chest to collect two dimensional echocardiogram of the papillary muscles. The left ventricular end-diastolic diameter (LVEDD) and LV end-systolic diameter (LVESD) were measured according to the myocardial contractility parameters (18, 20, 24). LV ejection fraction was calculated as (LVEDD)3−(LVESD)3)/(LVEDD)3)×100%) and LV fractional shortening equals to (LVEDD−LVESD)/LVEDD)×100%) (25).
Myocardial enzyme assessment
After the termination of echocardiography, orbital blood (1 ml) was taken, placed for 30 min and centrifuged at 3,000 g for 10 min at 4℃ (26, 27). Levels of myocardial troponin T (cTnT) (H149-4) and creatinine Kinase-MB (CK-MB) (H197) in serum were tested using the detection kits (NanJing JianCheng Bioengineering Institute, Nanjing, Jiangsu, China).
HE staining
The obtained myocardial tissues were paraffin-em-bedded and sliced at equal distances into sections (6 μm) for the following HE staining. Specifically, sections were deparaffinized through biological transparent reagent (Solarbio) and hydrated with graded ethanol double distilled water, stained with hematoxylin (Beyotime) for 5 min, rinsed using double distilled water, differentiated with 1% hydrochloric acid alcohol for 30 s and then subjected to another double distilled water wash, followed by eosin (Beyotime) staining for 3 min, successively dehydrated and transparent with graded ethanol absolute ethyl alcohol and biological transparent reagent (14, 28). In addition, sections were sealed and scanned with the digital scanning microscopy imaging system (Meyer Instruments Inc., Houston, TX, USA) to collect images.
TUNEL staining
Myocardial apoptosis was detected as myocardial tissue sections were assessed via the in situ cell death assay kit (Roche Applied Science, Mannheim GmbH, Penzberg, Germany) TUNEL staining. Next, the apoptotic nucleus and myocardial nucleus were stained with TUNEL or 4’, 6-diamidino-2-phenylindole. Apoptotic rate was calculated as number of TUNEL-positive apoptotic myocardial cells/total number of myocardial cells ×100% (21, 28, 29).
Dual-luciferase reporter gene assay
Luciferase reporter plasmids (pis CHECK2) containing TOLLIP wild type (WT) 3’untranslated regions (3’-UTRs) and TOLLIP mutant type (MUT) 3’-UTRs, respectively, were produced by General Biological System Co., Ltd. (Anhui, China). Lipofectamine 2000 (Invitrogen Inc., Carlsbad, CA, USA) was employed to co-transfect the reporter plasmids and miR-324-5p mimic or mimic NC (Shanghai Genechem Co., Ltd., Shanghai, China) to HEK293T cells (American Type Culture Collection, Manassas, Virginia, USA), which were inoculated into the 48-well plates. Then, luciferase activity of cell lysis buffer was examined through the Dual-Glo luciferase assay system (Promega Corporation, Madison, WI, USA).
RT-qPCR
The total RNA in ADSCs or myocardial tissues was extracted in line with the instructions of the TRIzol kits (Invitrogen). The total RNA concentration was determined using a spectrophotometer (German Eppendorf) and the purity of RNA was determined using 1% agarose gel electrophoresis. Then, the RNA was adversely transcribed into cDNA with the PrimeScript RT kits (Takara Biote-chnology Ltd., Dalian, China). The reverse transcription of miRNA was carried out by miRcute enhanced miRNA cDNA first strand synthesis kit (Beijing TianGen Biotech Co., Ltd. Beijing, China). RT-qPCR was performed when SYBR premix (Takara, Kusatsu, Japan) or miRcute enhanced miRNA fluorescence quantitative detection kit (SYBR Green) (TianGen Biotech) was applied on the Applied Biosystems 7500 PCR system (Carlsbad, CA, USA) following manufacturers’ instructions. The 2−ΔΔCt method (30) was appointed to calculate the relative expression of genes with glyceraldehyde-3-phosphate dehydrogenase (GAPDH) as the internal reference for TOLLIP and U6 as the internal reference for miR-324-5p. ΔΔCT=(CT value of target gene in experimental group− CT value of internal reference gene in experimental group)−(CT value of target gene in control group−CT value of internal reference gene in control group). Primer sequences of RT-qPCR were shown on Table 1.
Statistical analysis
SPSS 21.0 (IBM Corp. Armonk, NY, USA) was employed for data analysis and GraphPad Prism8.0 (GraphPad Software Inc., San Diego, CA, USA) for graph. Kolmogorov-Smirnov test indicated whether the data were in normal distribution. The results were displayed as mean±standard deviation. One-way analysis of variance (ANOVA) or two-way ANOVA for comparing each group, and Sidak’s multiple comparisons test or Tukey’s multiple comparisons test for pairwise comparisons after ANOVA. The p value was attained using a two-tailed test and p<0.01 indicated significant difference.
Results
Cultivation and identification of rat ADSCs
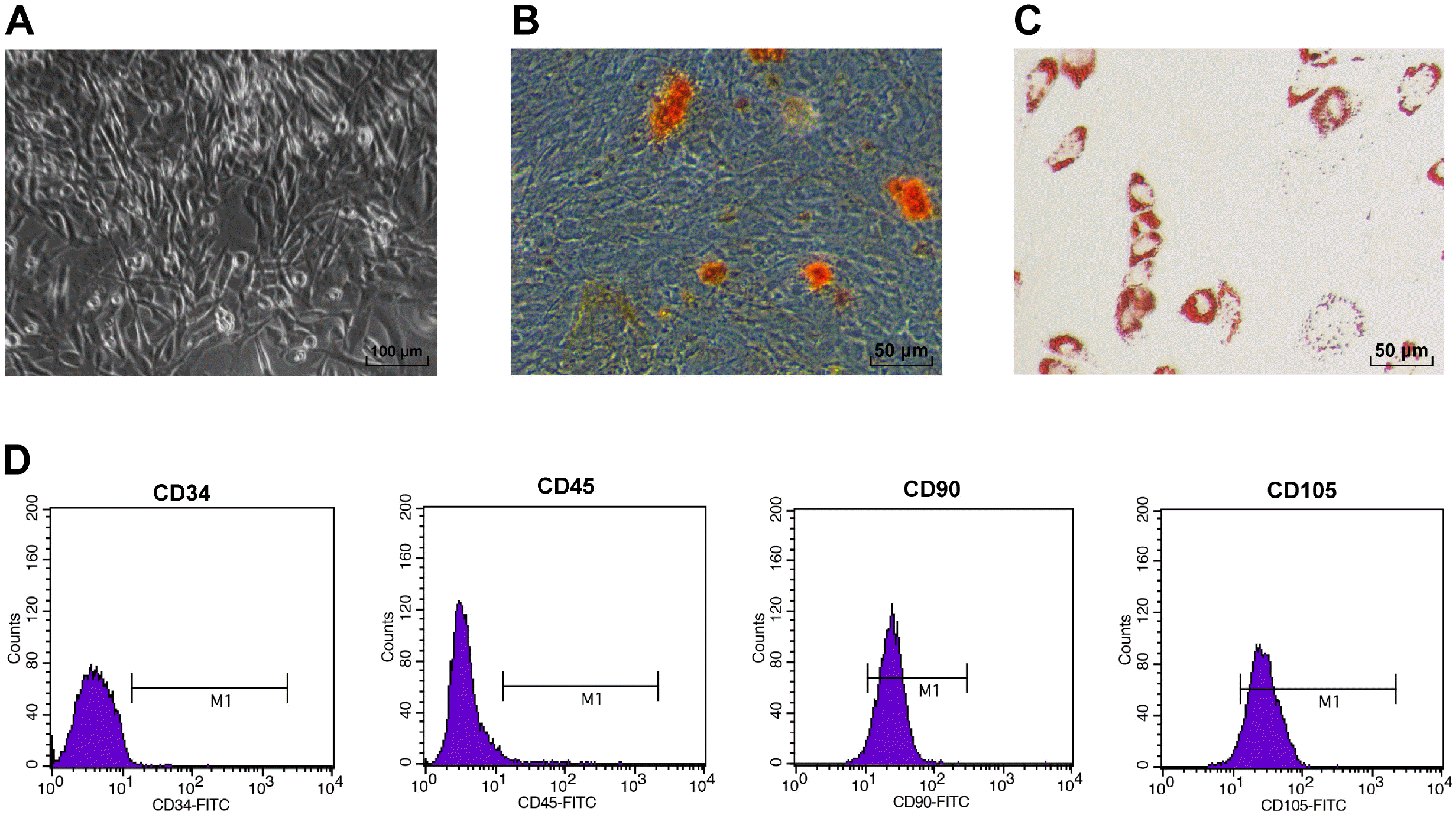
MI was a disease triggered by abrupt occlusion of coronary arteries and myocardial necrosis induced by ischemia and hypoxia (31, 32). Among various sources of SCs, MSCs, especially ADSCs, have been reported for the treatment of MI (7, 8). Although MSCs transplantation has witnessed a lot of successes in animal treatment, studies have shown that the cell survival rate after one week of transplantation was less than 10% (9, 33, 34). With the purpose of researching the role of ADSCs in MI, ADSCs were cultured, which exhibited the shape of long spindle under microscope (Fig. 1A). Induction differentiation carried out on ADSCs uncovered that cells were potent in getting adipogenesis and osteogenesis (Fig. 1B and 1C). Besides, CD105 (95.74%) and CD90 (90.56%), surface markers of MSCs, were overexpressed in ADSCs, while CD34 (1.21%) and CD45 (2.38%), markers of hematopoietic SCs, were not expressed (Fig. 1D). The above findings revealed that rat ADSCs were successfully cultured.
miR-324-5p promotes ADSC proliferation and migration
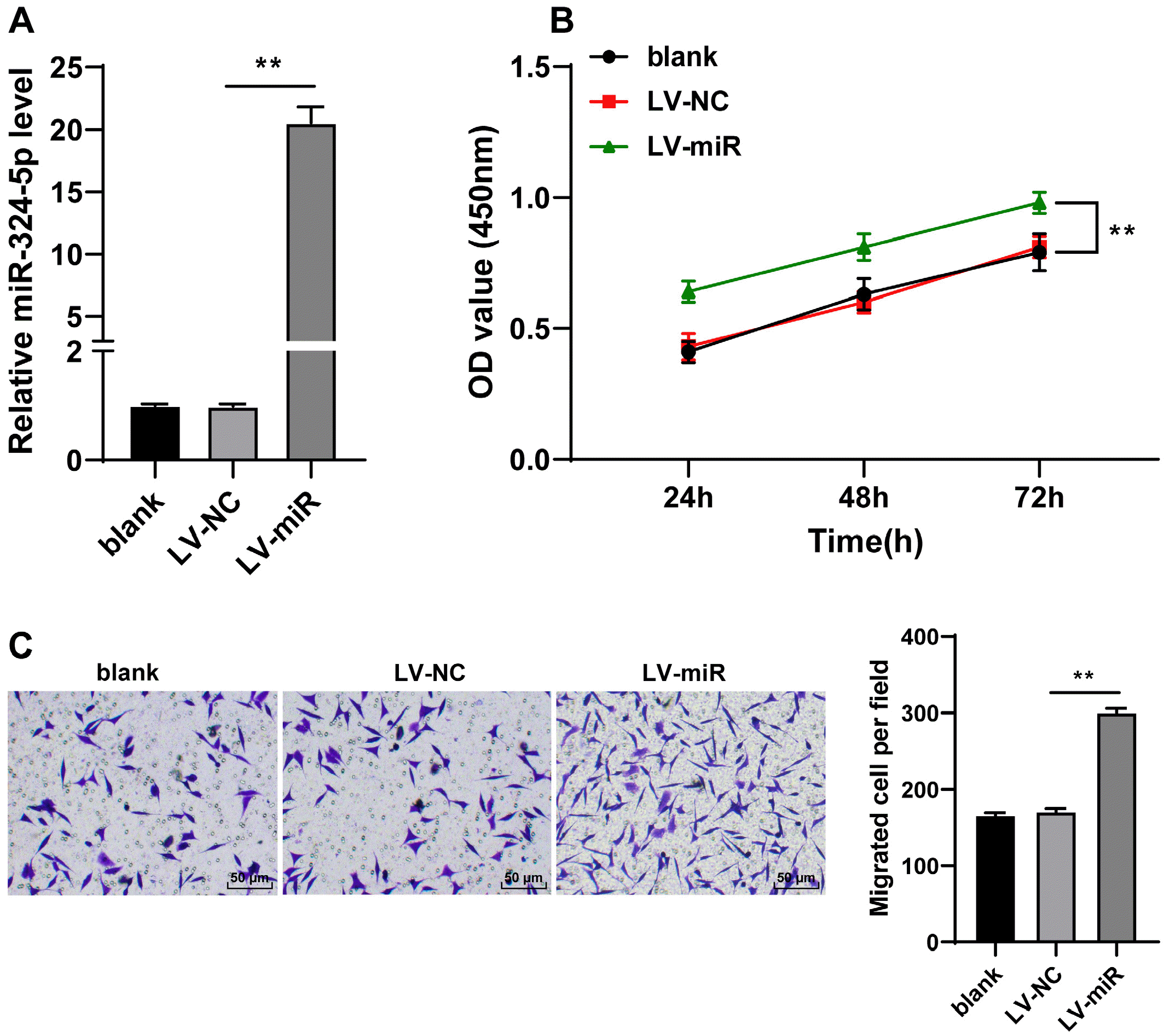
To explore the role of miR-324-5p on ADSCs, miR-324-5p expression was upregulated in ADSCs by the infection of ADSCs using LV-miR-324-5p (p<0.01, Fig. 2A). It was unmasked from CCK-8 method and Transwell assay that ADSC proliferation and migration were enhanced in the LV-miR group compared with the LV-NC group (p<0.01, Fig. 2B and 2C), indicating that overexpressed miR-324-5p enhanced ADSC dissemination.
miR-324-5p-modified ADSCs enhance post-MI repair in rats
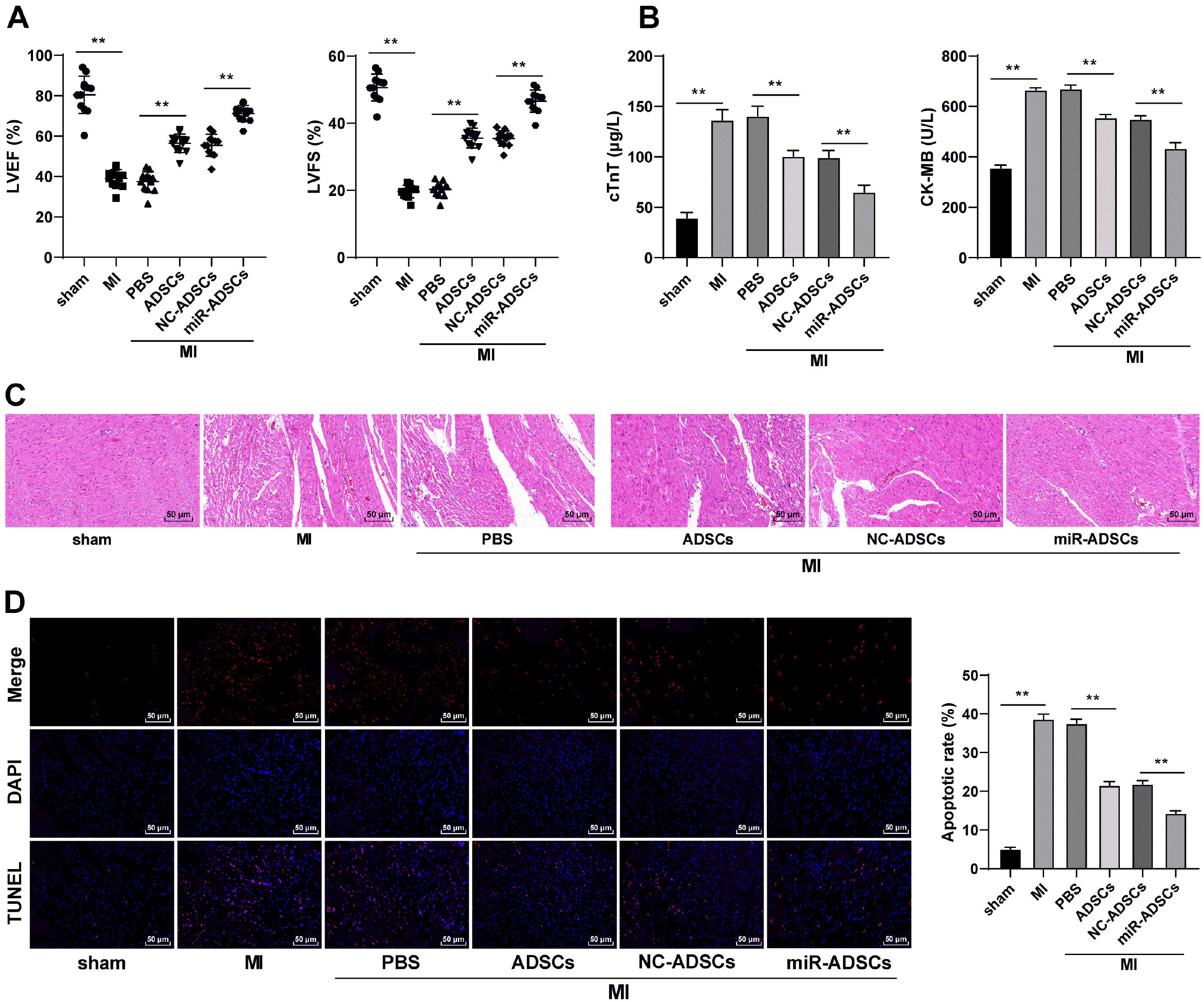
To test the function of miR-324-5p-modified ADSCs in MI treatment, MI model was established in rats, with ADSCs administrated into the peri-MI area at multiple sites. As shown in Fig. 3A, myocardial function of MI rats was evidently impaired, while ADSCs treatment alleviated myocardial function impairment and decreased the serum levels of cTnT and CK-MB in MI rats, but the function of miR-ADSCs was even more efficient (p<0.01, Fig. 3A and 3B). HE staining and TUNEL staining conduced on myocardial tissue sections displayed that the degree of MI was aggravated and apoptosis was increased in MI rats, which was reversed in the ADSC group and the miR-ADSC group, and the miR-ADSC group exhibited was more effective than the ADSC group (p<0.01, Fig. 3C and 3D). In summary, miR-324-5p overexpression in ADSCs could improve post-MI myocardial repair in rats.
miR-324-5p targets TOLLIP expression in MI rats
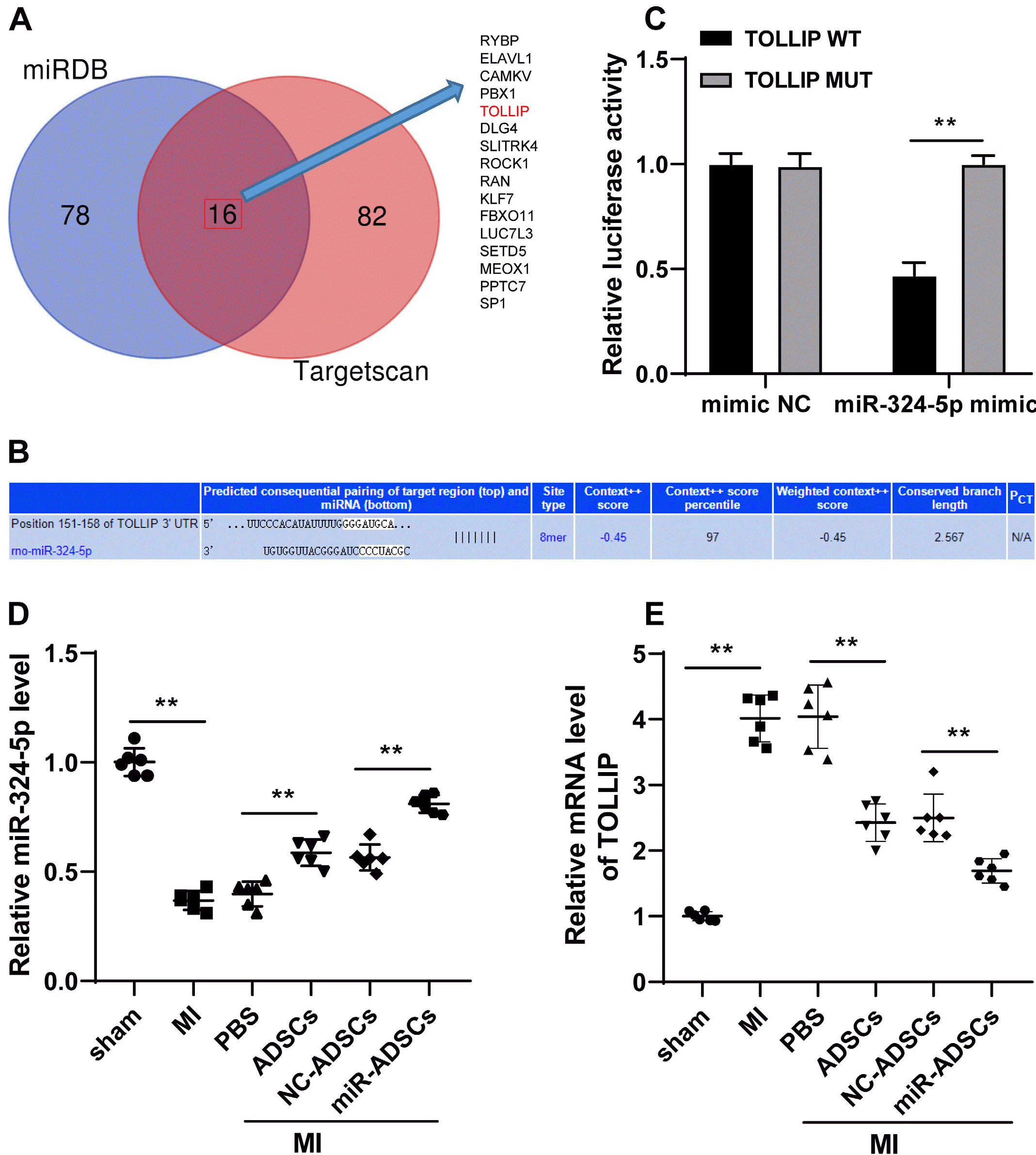
Next, to further reveal the downstream mechanism of miR-324-5p in myocardial tissues, the downstream target genes of miR-324-5p was predicted and intersected by miRDB and Targetscan databases, and 16 target genes were noticed (Fig. 4A), of which TOLLIP was reported to promote the occurrence of MI (16). Thus, dual-luciferase reporter gene assay was designed based on the binding site of miR-324-5p and TOLLIP (Fig. 4B), which showed the target binding relationship between miR-324-5p and TOLLIP (p<0.01, Fig. 4C). In addition, ADSCs treatment upregulated miR-324-5p expression and downregulated TOLLIP expression; while miR-324-5p overexpression led to downregulated TOLLIP (p<0.01, Fig. 4D and 4E). That’s to say, miR-324-5p targeted TOLLIP.
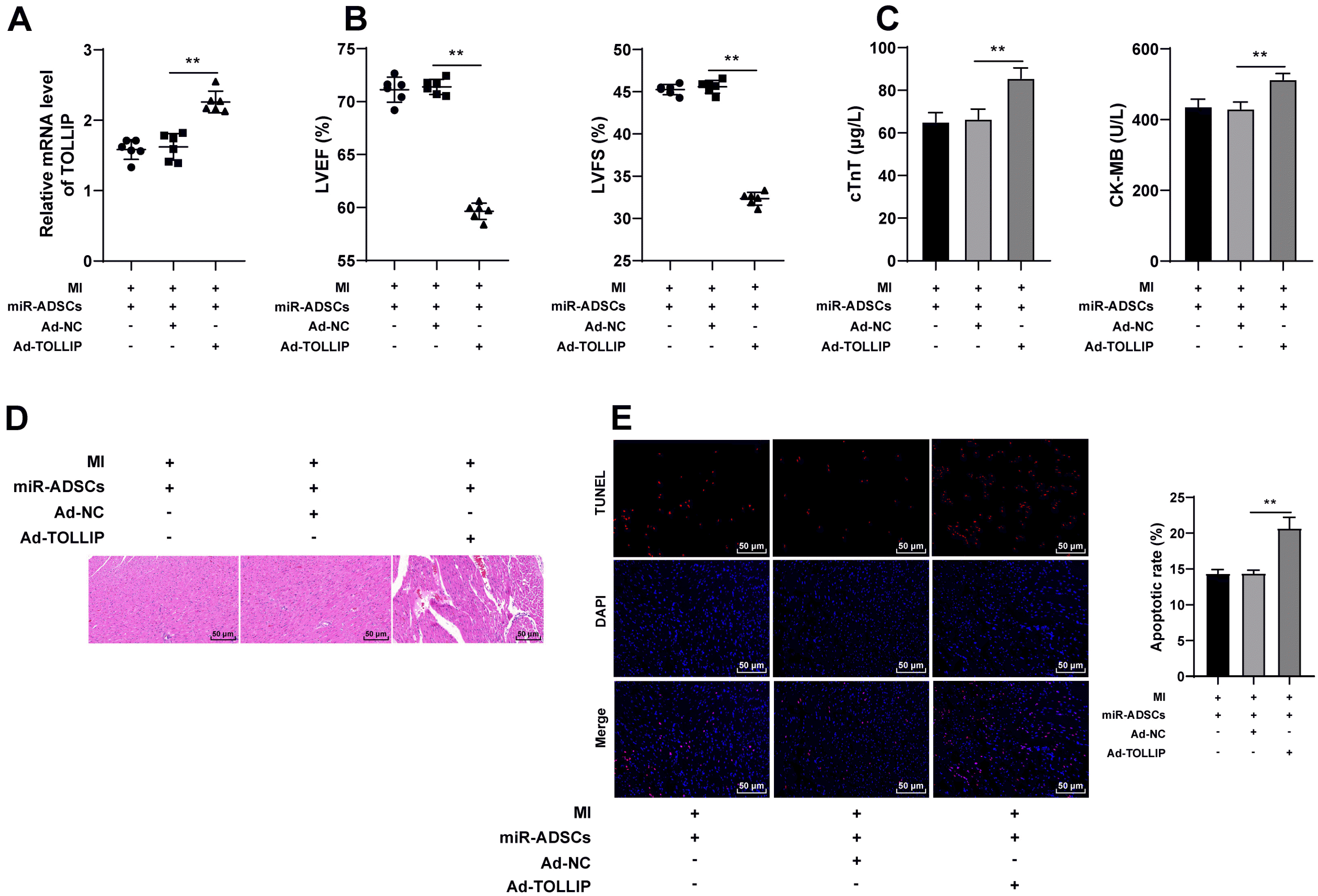
TOLLIP overexpression spoils the promotive role of miR-324-5p-modified ADSCs in post-MI repair in rats
To confirm that miR-324-5p-modified ADSCs promoted myocardial repair by targeting TOLLIP in myocardial tissues, functional rescue assay was conducted as TOLLIP expression was successfully upregulated in myocardial tissues via the injection of Ad-TOLLIP in rats from the miR-ADSCs group (p<0.01, Fig. 5A), and it was subsequently uncovered that the myocardial function of MI rats was attenuated (p<0.01, Fig. 5B), and the levels of cTnT and CK-MB in serum were elevated (p<0.01, Fig. 5C). Additionally, after overexpressing TOLLIP, both the degree of MI and apoptotic cells were greatly increased in the myocardial tissues of MI rats (p<0.01, Fig. 5D and 5E). These results illustrated that TOLLIP overexpression spoiled the promotive role of miR-324-5p-modified ADSCs in post-MI repair in rats.
Discussion
MI referred to a concept in which plaques accumulated at interior wall of arteries to block blood flow toward heart and lead to oxidative stress-induced myocardial injuries, eventually causing cardiac disease (2). For the properties of availability and renewability, ADSCs emerged as a popular choice for target treatment of post-MI repair (35). Accumulating reports suggested that miRs-modified ADSC therapy helped to reduce fibrosis, inflammation and cell death in MI when compared with ADSC therapy alone (17). Furthermore, miR dysregulation was critical player in myocardial hypertrophy and cardiac fatigue of MI (36). Woods and his colleagues uncovered that miR-324-5p could positively regulate MSC osteogenic development to maintain homeostasis and embryogenesis in patients with osteoarthritis (37). Therefore, we were inspired to explore the possible role of miR-324-5p-modified ADSCs in post-MI repair.
Firstly, in our experiments, the fact that miR-324-5p promoted ADSC proliferation and migration was noticed. It was recently unveiled that different miRs were related to the biological processes of SCs including dissemination, differentiation and apoptosis (38, 39). Interestingly, miR-324-3p propelled keratinocyte expansion, differentiation and self-renewal (40). On the other hand, inhibition of miR-324-5p impeded synapse construction of astrocyte neurons, which might incur senescence, inflammatory response and neurodegenerative disorders (41). Furthermore, miR-324-3p was activated in adipose tissues at multiple organs of obese mice, and downregulation of miR-324-3p resulted in loss of fat and body-weight, suggesting the positive correlation of miR-324-3p and ADSCs (42). Meanwhile, the interaction between miRs and ADSCs has gained a lot of success. For instance, encouraged by miR-375, ADSCs could better strengthen bone regeneration (43). Besides, miR-181-5p-modified ADSCs restored injured liver cells and interrupt liver fibrosis (44). From these progresses mentioned above, the positive crosstalk of miR-324-3p and ADSCs could be a contributory factor in post-MI repair. Consequently, we found that miR-324-5p-modified ADSCs enhanced post-MI repair in rats with the involvement of decreased levels of cTnT and CK-MB and compromised apoptosis. Apoptosis brought about retarded heart function and severe heart damage (45). CK-MB and cTnT, as common targets in heart diseases, were correlated with many cardiac specific miRs to regulate acute coronary syndrome (46). miR-324-5p overexpression could efficiently alleviate angiogenesis and pernicious cell expansion, eventually decelerating pulmonary arterial hypertension (47). On the other hand, miR-324 relieved cardiomyocyte damage and oxidative stress of myocardial injury triggered by hypoxia/reoxygenation (13). Surprisingly, Huang and his colleagues (48) reported that miR-324-5p activation attenuated apoptosis and ventricular cardiomyocyte injury exacerbated by myocardial ischemia-reperfusion to relieve MI. Moreover, miR-modified MSCs maintained ventri-cular construction and expedited post-MI repair through depleting cell death and apoptosis (49). As a class of MSCs, ADSC transplantation was extensively applied in alleviation and repair of diseases like diabetes, facial restoration, osteoarthritis and cardiac disorders (50). The accumulation of ADSCs after MI unleashed the release of vascular endothelial growth factor and led to angiogenesis (51). Moreover, ADSCs were able to remodel blood vessels when they differentiated to vascular net (52). Significantly, it was previously unmasked that miR-342-5p modified ADSCs potentiated endothelial cells to defense atherosclerosis (53). In addition to this, ADSCs limited the level of cTnT (54). Altogether, it might be quite certain that miR-342-5p modified ADSCs served as contributor to post-MI repair.
Last but not least, we found that miR-324-5p targeted TOLLIP in myocardial tissues of MI rats. As a predominant cytokine in immunological response guided by Toll-like receptor, TOLLIP proactively participated autophagy and inflammation in tumors and human disorders related to liver, neurodegeneration and angiocarpy (55, 56). According to Ryan et al. (57) TOLLIP upregulation was coupled with growing expression of Parkin and it represented a manifestation of mitochondrial injury, which finally resulted in deteriorative Parkinson’s disease. A recent finding revealed that TOLLIP exhaustion plainly degraded liver damage inflicted by ischemia-reperfusion (58). That’s to say, TOLLIP imposed adverse effect on diseases related to inflammatory reaction and immune system. Consistently, our experiment uncovered that TOLLIP overexpression spoiled the promotive role of miR-324-5p-modified ADSCs in post-MI repair in rats as presented by elevated levels of levels of cTnT and CK-MB and improved apoptosis. As a pivotal diagnosis biomarker in MI, cTnT upregulation could greatly destroy heart function (59). TOLLIP depletion was associated with stable plaques of atherosclerosis areas, attenuated inflammatory response, growing circulating lipids and impaired lipophagy in atherosclerosis (60). Besides, as an acknowledged cardiovascular modulator, TOLLIP aggravated neurocyte apoptosis and inflammatory reaction in individuals injured by ischemia-reperfusion (61). All in all, miR-342-5p modified ADSCs could work as a promising inhibitor of MI by targeting TOLLIP.
In summary, our findings revealed that miR-342-5p modified ADSCs promoted post-MI repair by targeting TOLLIP. These results originally discovered an approach for MI treatment. Still, some limitations are to be solved. For example, this study merely uncovered the modifying effect of miR-324-5p on ADSCs, without clarifying it there is any effect on SCs. In addition, whether there are other miRNAs involved is yet to know. Furthermore, other downstream target genes of miR-324-5p need to be investigated. The purpose of this study is to investigate the role of miRNA modified ADSCs in myocardial repair after MI. We did not detect ADSCs differentiation in rat hearts, but some studies showed that the adipose stem cells of heart transplantation were differentiated into cardiomyocytes as a whole (62, 63), which we will take as our next research direction. The “paracrine effect” has been proposed as a promising strategy to boost endogenous reparative and regenerative mechanisms within the cardiac tissue (64). Stem cells are known to secrete various factors that have paracrine activities; they home to hypoxic and/or inflamed areas, and release trophic factors that hasten endogenous repair and the secreted bioactive factors limit local immune system, enhance angiogenesis, inhibit fibrosis and apoptosis, and stimulate differentiation of tissue-residing stem cells (65). Once transplanted into a damaged organ, MSCs release paracrine factors that nurture the injured area, prevent further adverse cardiac remodeling, and mediate tissue repair along with vasculature (66-68). We will focus on whether miR-324-5p is carried into cardiomyocytes by ADSCs through paracrine effect in the next research.
 XML Download
XML Download