

 PDF
PDF Citation
Citation Print
Print


Introduction
Lumbar fusion surgery is mainly used for the treatment of lumbar disc herniation, lumbar spinal stenosis and lumbar spondylolisthesis (1). The principle of its fusion is to take the diseased vertebra as the center, and to fuse the diseased upper vertebra with the next vertebra as a unit. At present, the bone materials mainly include autogenous, allograft, xenograft, and alloplast (2). Autogenous bone graft is regarded as the main bone graft material (3), but the mechanism of its fusion remains unclear.
Adult bones undergo a process of continuous remodeling, beginning with the resorption of the mineralised bone matrix by osteoclasts, followed by the modification of the skeleton by osteoblasts, filling the space between the graft and the diseased upper and lower vertebrae for fusion (4, 5). However, osteoblasts are differentiated short-lived cells, therefore bone growth and maintenance require continuous replacement of new osteoblasts produced by bone marrow mesenchymal stem cells (BMSCs) (6). BMSCs are a kind of primitive cells with self-renewal and multidirectional differentiation potential, and are the main source of osteoblasts in vivo (7). Their proliferation and osteogenic differentiation are of great significance for maintaining the number of osteoblasts (7).
MicroRNAs (miRNAs) are a class of endogenous, non-coding small RNAs (18-24 nucleotides) that inhibit the initiation of translation or induce cleavage of mRNAs by specific pairing with the 3’ Untranslated Regions (UTR) of the target mRNAs (8). MiRNAs are important bio-regulatory factors that can affect cell differentiation, proliferation, and apoptosis (9). Current researches demonstrated that miRNAs play critical roles in osteogenic differentiation of BMSCs (10, 11). Tamura et al. (12) found that miR-34 expression is upregulated during osteogenic differentiation. Furthermore, Liu et al. (13) showed that miR-34c could elevate osteogenic differentiation of mesenchymal stem cells (MSCs). Besides, miR-34c could also enhance the activity of osteoblasts (14). However, the specific mechanism of its regulation of osteogenic differentiation and proliferation was still unclear. Accumulating evidence has shown that miR-34c-5p could negatively regulate Bcl-2 (15), which is an anti-apoptotic protein (16). Recombinant overexpression of Bcl2 promotes the proliferation but inhibits the osteogenic/odontogenic potential of dental pulp stem cells (17). In addition, TargetScan 7.2 predicted that miR-34C-5p could target BCL2. Based on these studies, we hypothesized that miR-34c-5p could affect osteogenic differentiation and proliferation of BMSCs through regulating Bcl-2.
In this study, the expression of miR-34c-5p in the osteogenic differentiation induction process of BMSCs was detected. Besides, the effect of miR-34c-5p on the proliferation, osteoblastic differentiation, calcium deposition, and alkaline phosphatase (ALP) expression of BMSCs were also investigated. This study investigated the mechanism of autologous bone grafting in lumbar fusion surgery based on BMSCs.
Go to : 
Materials and Methods
Ethics statement
The animal experiment of this study was carried out in strict accordance with the guidelines of China Animal Care and Use Committee, and the discomfort and pain of experimental animals were minimized during the processes. The experiment was conducted in Nanfang Hospital, and the study was approved by the Experimental Animal Committee of Nanfang Hospital (NF201811273).
Establishment rabbit model of lumbar fusion
Twenty-four healthy adult male New Zealand white rabbits (2.5∼2.7 kg, 2∼3 months old) were randomly divided into two groups. In the modeling group (12 rabbits), bilateral autogenous bone marrow intertransverse bone graft posterolateral fusion was performed at L5-L6 segment, and another 12 rabbits at the same weight in the control group were performed with L5-L6 segment posterolateral fusion. The rabbits were sacrificed at 4 weeks, 8 weeks and 12 weeks after the surgery, respectively. The accuracy of fusion was determined by comparison of X-ray and CT scan. Tissues surrounding the bone graft were collected for qRT-PCR detection.
Bioinformatics analysis
Datasets GSE115197 and GSE19232 on osteogenic differentiation of MSCs were obtained from Gene Expression Omnibus (GEO) database (https://medworm.com/journal/geo-gene-expression-omnibus/). Differential expression of miRNA was analyzed by limma packet (R language). TargetScan 7.2 (http://www.targetscan.org/vert_72/) was used to predict the targeted binding sites of miR-34c-5p and Bcl-2.
Cell culture and differentiation
Human bone marrow-derived mesenchymal stem cells (BMSCs) were purchased from ATCC (ATCCⓇ PCS-500-012TM, Manasas, VA, USA) and maintained in DMEM/F12 medium (C11330500BT, Gibco,Grand Island, NY, USA ) containing 10% FBS (10099-141,Gibco) in an incubator (37℃, 5% CO2).
For osteogenic differentiation of BMSCs, briefly, the cells were maintained in a specific medium for 14 days (d), and the medium was changed every 3 d. Specific culture medium was prepared (6) by supplementing DMEM/F12 with 1% HyClone serum (16050-122, Gibco), 200 μM L-glutamine (25030-081, Gibco), 100 nM dexamethasone (D4902, purity≥97%, Sigma aldrich, St. Louis, MO, USA), 10 mM β-glycerolphosphate (50020, purity≥98%, Sigma aldrich), 10 nM dihydroxyvitamin D3 (50020, purity≥99%, Sigma aldrich) and 80 μg/ml L-Ascorbic acid (A7506, Sigma aldrich).
Quantitative reverse transcription polymerase chain reaction (qRT-PCR)
Total RNA was extracted from the differentiated BMSC cells at 0, 3, 7 and 14 d using the mirVana kit (AM1556, Invitrogen, Carlsbad, CA, USA). Then 1 μg of total RNA was reverse-transcribed into cDNA using a reverse transcription kit (RR037A/D350A, TaKaRa, Kyoto, Japan). Then the reaction system of 20 μl consisted of 2 μl cDNA, 10 μl SYBR Mix, 0.8 μl primers, and 6.4 μl distilled water was prepared according to the kit instructions (RR820A/DRR081, TaKaRa, Kyoto, Japan). The primers for this experiment were provided by Sangon Biotech (Shanghai, China) and Ribobio (Guangzhou, China), and the primer sequences were shown in Table 1. The target gene was then amplified in the ABI 7500 reaction system (Biosystems, Foster City, USA) according to the reaction conditions (at 95℃ for 30 seconds (s) and 40 cycles at 95℃, for 5 s, then at 60℃ for 30 s). The CT value obtained from ABI 7500 was calculated by 2−ΔΔCT method (18). U6 was the internal reference of miRNAs and GAPDH was the internal reference of RUNX2, OCN and Bcl-2.
Table 1
Specific primer sequences for quantitative reverse transcription polymerase chain reaction
![]()
Enzyme linked immuno sorbent assay (ELISA)
The ELISA kit for ALP was obtained from Nanjing SenBeiJia Biological Technology Co., Ltd. (H0318, Nanjing, China, http://www.senbeijia.com/). To be brief, gradient standard substance (concentrations at 90 U/l, 60 U/l, 30 U/l, 15 U/l and 7.5 U/l) was prepared according to the instructions and added into the substrate plate at 50 μl/well.
Then, blank hole and sample hole were set, without liquid in the blank hole, and 40 μl sample diluent and 10 μl sample were added to the sample hole. After the liquid was added, the plate was sealed with a sealing film and incubated for 30 min (at 37℃). After the liquid was removed, the plate was dried. The plate was washed by adding sashing liquid for 5 times (30 s for each time) and dried. Subsequently, 50 μl of enzymatic reagent was added to each well (except for blank holes). Incubation and washing were the same as above. The reagent A of 50 μl was added into each hole, which was then added with 50 μl reagent B. After gent shaking and mixing, the plate was incubated in the dark for 15 min (at 37℃). Finally, 50 μl termination solution was added into each well to terminate the reaction. After the reaction was terminated for 15 min, the absorbance (OD value) at 450 nm wavelength was measured using a microplate reader (Molecular Devices, Sunnyvale, CA, USA).
Alizarin red staining
BMSCs at 0, 3, 7, and 14 d of differentiation were used for alizarin red staining. After discarding the medium in the 6-well plate, the plate was rinsed in PBS for 3 times (3 min for each time). Then, 4% paraformaldehyde (P0099, Beyotime, Shanghai, China) was used to immobilize the cells at room temperature for 10 min. Afterwards, alizarin red solution (20 mM, ST1078, Beyotime) was used to treat each well and stain the cells for 5 min at room tem-perature. Finally, the solution in the cells was removed and rinsed again in PBS for 3 times (3 min for each time). A small amount of water was retained in the six-well plate, the cells were observed under an inverted microscope (POMEAS, Guangzhou, China) and photographed (magni-fication at ×200).
Cell transfection
Lentivirus used for transfection (including miR-34c-5p mimic, miR-34c-5p inhibitor, Bcl-2 overexpression, siBcl-2 and its control) in this study were designed and constructed by Genepharm (Shanghai, China). For the transfection of lentivirus into BMSC cells, the cells were planted into a 24-well plate at 1×105 cells/well. After 24 h, the original culture medium was replaced with 2 ml fresh medium containing 6 μg/ml polybrene (H8761, Solarbio, Beijing, China) and 4 μl lentivirus suspension. The cell culture continued for another 24 h, and then the medium containing the virus was replaced with a fresh medium.
Dual luciferase reporter gene assay
The 3’UTR sequence of Bcl-2 was acquired from NCBI (https://www.ncbi.nlm.nih.gov/). Based on the predicted result of TargetScan database, we designed the mutation sequence of Bcl-2’s 3’ UTR sequence. The cloned Bcl-2-WT and Bcl-2-MUT sequences were inserted into pmiRGLO dual luciferase reporter vector (E1303, Promega, Madison, WI, USA) to obtain Bcl-2-WT and Bcl-2-MUT plasmid.
The BMSC cells were planted in 6-well plates at 2×105 cells/well. Co-transfection was conducted after the degree of cell fusion reached 70∼80%. The co-transfection groups were specifically as follows: Bcl-2-WT+miR-34c-5p mimic, Bcl-2-WT+mimic control, Bcl-2-MUT+miR-34c-5p mimic, Bcl-2-MUT+mimic control, Bcl-2-WT+miR-34c-5p inhibitor, Bcl-2-WT+inhibitor control, Bcl-2-MUT+miR-34c-5p inhibitor and Bcl-2-MUT+inhibitor control. The transfection was conducted the same as above. The cells were lysed after transfection for 30 h, and fluorescence intensity was measured with a dual luciferase assay kit (E1910, Promega).
Cell counting kit (CCK)-8
After transfection with lentivirus, the cells began to differentiate. The cells were planted in a 96-well plate at 37℃. Cell proliferation was determined by CCK-8 kit (C0038, Beyotime) at 0, 3, 7 and 14 d of the induction of differentiation. For CCK-8 assay, the CCK-8 reagent of 10 μl was added to target cells and cultured together for 3 h at 37℃ (5% CO2). Then, the culture plate was taken out, and the OD value at 450 nm was measured using a microplate reader.
Western blot
The treated or untreated cells were collected, and the total protein was extracted by RIPA reagent (P0013B, Beyotime) with 1% protease inhibitors (P1030, Beyotime). After that, the concentration of the protein was determined by BCA kit (P0012, Beyotime). Then, cellular protein of 30 μg was separated by SDS-PAGE gel (P0012A, Beyotime). After electrophoresis for 90 min, the proteins were transferred on PVDF membranes (ISEQ00010/IPVH00010, MILLIPORE, MA, USA). Then, the membranes were blocked by 5% skimmed milk (D8340, Solarbio) for 1 h at room temperature, followed by incubation in primary antibodies (RUNX2, ab23981, 1:1,000, 60 kDa, Abcam, Cambridge, MA, USA; OCN, ab93876, 1:500, 11 kDa, Abcam; BCL2, ab182858, 1:2,000, 26 kDa, Abcam; GAPDH, ab181602, 1:10,000, 36 kDa, Abcam) at 4℃ overnight. Next day, the primary antibodies were recycled, and TBST (ST-673, Beyotime) was used to wash the membranes for 4 times (5 min for each time). Afterwards, secondary antibody (Goat Anti-Rabbit IgG H&L (HRP): ab6721, 1:10,000, Abcam) was incubated the membranes for 1 h (room temperature), followed by washing with TBST for 6 times (5 min for each time). Finally, ECL solution (WBKLS0500, MILLIPORE, Billerica, MA, USA) was dripped onto the membrane, and a specific imaging system (Bio-Rad, CA, USA) was used to visualize the band.
Statistical analysis
All data were expressed as the mean±standard deviation. Comparison among groups was performed by evaluating one-way ANOVA, followed by Turkey’s test. Paired-sample t-test was used to analyze paired differences. p<0.05 were considered as statistically significant. All the data were analyzed using SPSS (IBM, NY, USA).
Go to :
Results
MiR-34c-5p was positively correlated with osteogenic differentiation
Cross-analysis of two mesenchymal stem cell osteogenic differentiation datasets (GSR115197 and GSE19232) screened five candidate miRNAs (miR-483-3p, miR-34c-5p, miR-199a-5p, miR-324-5p, and miR-361-5p) (Fig. 1A). However, as only miR-34c-5p, miR-199a-5p, miR-324-5p and miR-361-5p were conserved in human and rabbit, we subsequently detected the expressions of miR-34c-5p, miR-199a-5p, miR-324-5p and miR-361-5p in the tissues from rabbit lumbar fusion model. The results of qRT-PCR demonstrated that compared with the control group, the expressions of miR-483-3p, miR-199a-5p, miR-324-5p and miR-361-5p showed no significant difference, while the expression of miR-34c-5p was obviously higher in rabbit lumbar fusion model group than that in the control group (Fig. 1B; p<0.001). Thus, miR-34c-5p was determined as the target in this study. Then we verified the role of miR-34c-5p in osteogenic differentiation of BMSCs at a cellular level. According to our detection, the expression of miRNA-34c-5p was gradually upregulated during the process of osteogenic differentiation (Fig. 1C; p<0.001). Besides, the content of ALP and calcium deposition (calcium in the alizarin red staining tissues, the red part in the figure shows calcium deposition) was also gradually increased with the osteogenic differentiation of BMSCs (Fig. 1D and 1E; p<0.001). This suggests that miR-34c-5p was positively correlated with osteogenic differentiation of BMSCs.
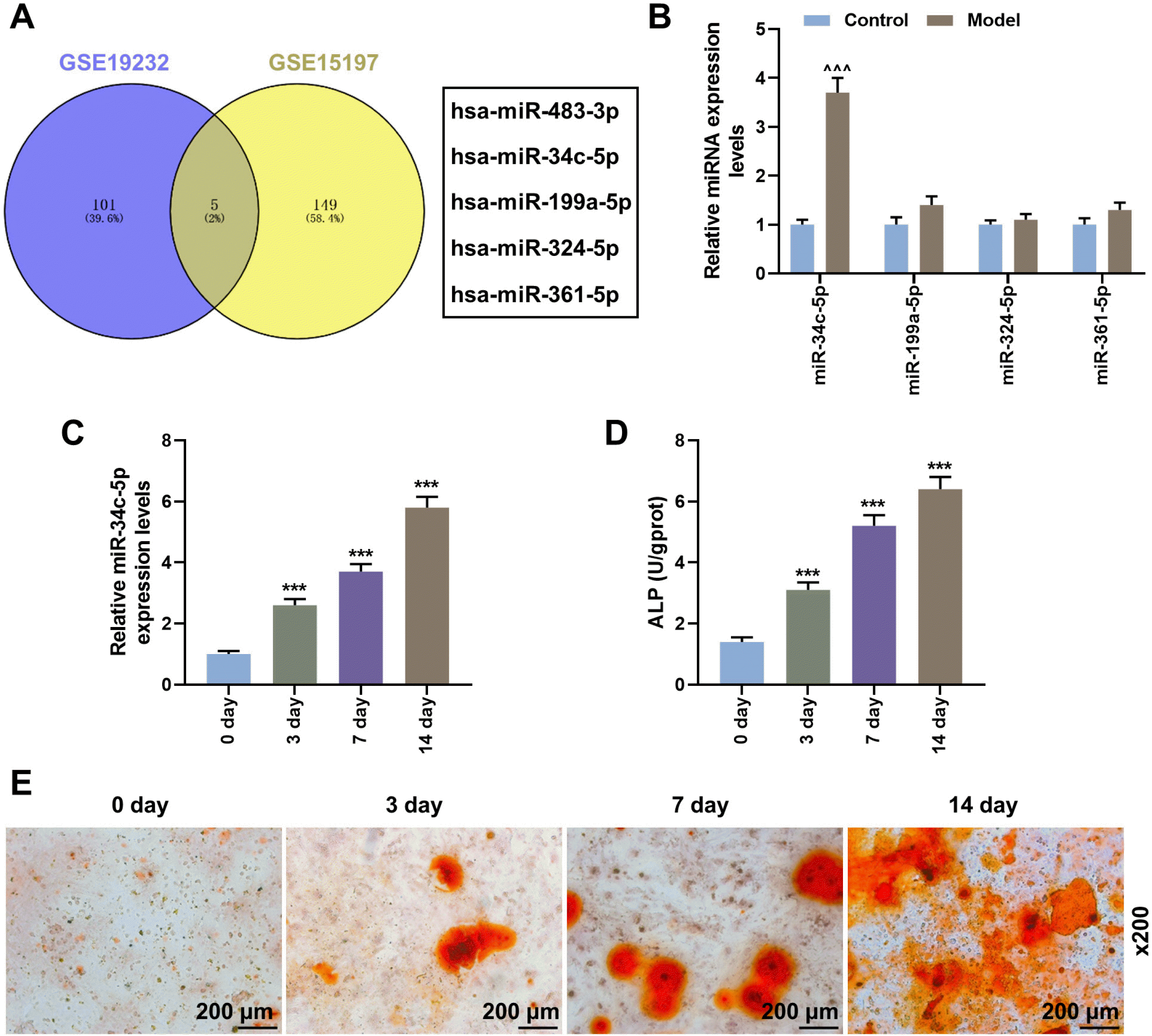 | Fig. 1During osteogenic differentiation of BMSCs, miR-34c-5p expression, ALP content and calcium deposition were promoted. (A) Abnormal expressions of miRNAs during osteogenic differentiation of mesenchymal stem cells obtained from the GEO database (GSE115197 and GSE19232). (B) The expressions of miR-34c-5p, miR-199a-5p, miR-324-5p and miR-361-5p in rabbit lumbar fusion model rabbit or normal rabbit were determined by qRT-PCR. (C) qRT-PCR was used to detect the expression of miR-34c-5p in BMSCs with different differentiation times (0, 3, 7 and 14 day). (D) ELISA was used to measure the content of ALP in BMSCs at different differentiation times (0, 3, 7 and 14 day). (E) Alizarin red staining of BMSCs at different differentiation times (0, 3, 7 and 14 day). Scales: 200 μm; magnifications: ×200. ^vs. Control, *vs. 0 days. ^^^ or ***p<0.001. BMSCs, bone marrow mesenchymal stem cells; ALP, alkaline phosphatase; GEO, gene expression omnibus; qRT-PCR, quantitative reverse transcription polymerase chain reaction; ELISA, enzyme linked immune sorbent assay. The experiment was independently repeated three times.
|
MiR-34c-5p promoted osteogenic differentiation of BMSCs
To further understand the relationship between miR-34c-5p and osteogenic differentiation of BMSCs, miR-34c-5p was up-regulated or down-regulated in BMSCs. Transfection efficiency of miR-34c-5p mimic and miR-34c-5p inhibitor were shown in Fig. 2A and 2B. MiR-34c-5p mimic obviously upregulated the expression of miR-34c-5p, and miR-34c-5p inhibitor significantly inhibited the expression of miR-34c-5p as compared with the control group (p<0.01, p<0.001). Afterwards, the results of CCK-8 proved that miR-34c-5p mimic could inhibit the proliferation of BMSCs compared with the control group, while miR-34c-5p inhibitor had the opposite effect (Fig. 2C and 2D; p<0.05). Subsequent experimental results showed that miR-34c-5p mimic increased the content of ALP and calcium deposition compared with the control group, and miR-34c-5p inhibitor had an opposite effect (Fig. 2E∼H; p<0.01, p<0.001). The detection results of osteogenic differentiation-related genes were also consistent with the above results. According to Fig. 3A∼F, miR-34c-5p mimic upregulated the expressions of RUNX2 and OCN (either protein level or mRNA level) compared with the control group (p<0.01, p<0.001). However, inhibition of miR-34c-5p was the opposite.
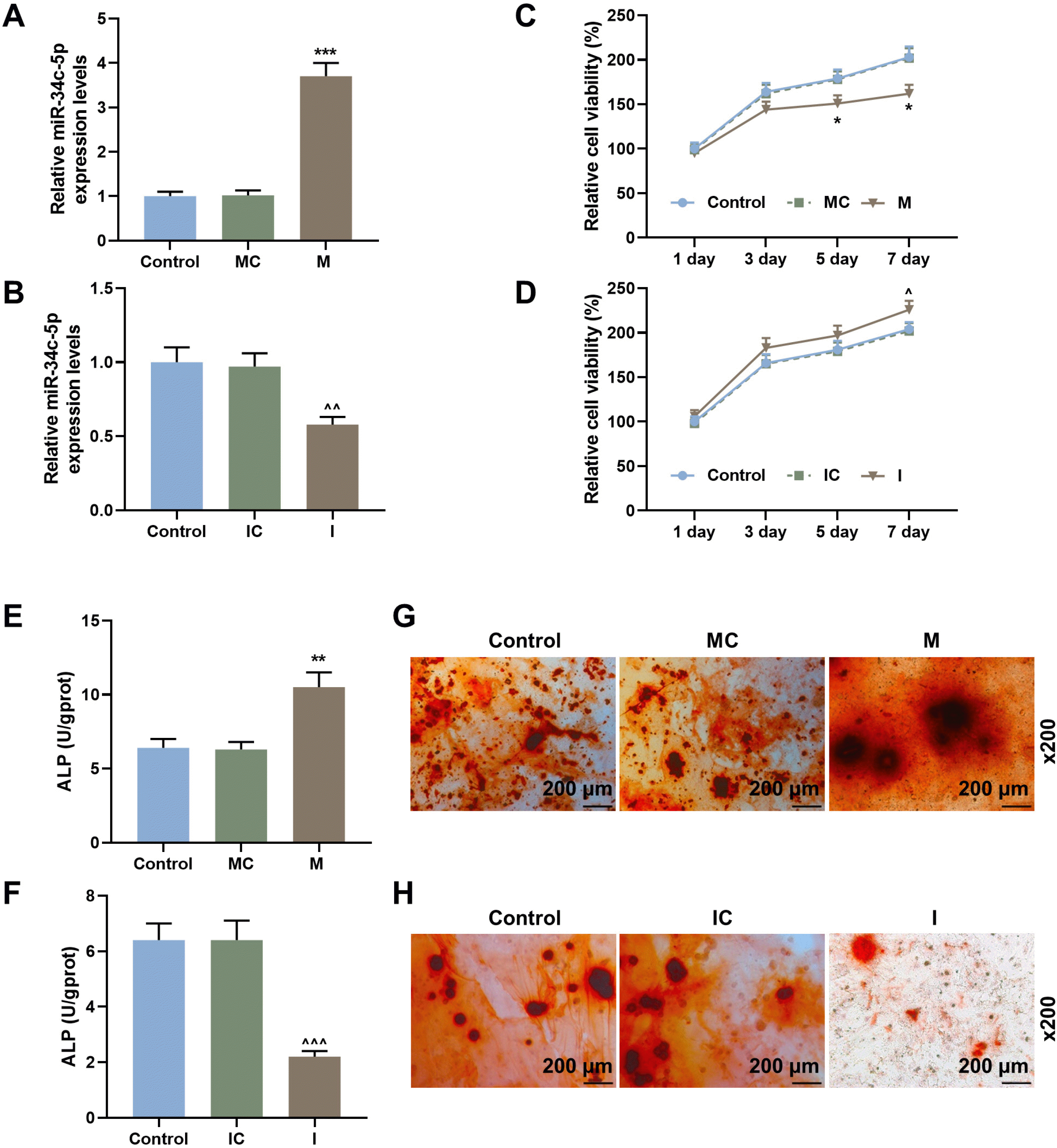 | Fig. 2miR-34c-5p decreased cell proliferation and increased ALP content and calcium deposits of BMSCs. (A) The expression of miR-34c-5p in BMSCs transfected with miR-34c-5p mimic or mimic control was detected by qRT-PCR. (B) The expression of miR-34c-5p in BMSCs transfected with miR-34c-5p inhibitor or inhibitor control was detected by qRT-PCR. (C) The cell proliferation of BMSCs transfected with miR-34c-5p mimic or mimic control was identified by CCK-8 at different times (1, 3, 5 and 7 day). (D) The cell proliferation of BMSCs transfected with miR-34c-5p inhibitor or inhibitor control was identified by CCK-8 at different times (1, 3, 5 and 7 day). (E) The ALP content of BMSCs transfected with miR-34c-5p mimic or mimic control was detected by ELISA. (F) The ALP content of BMSCs transfected with miR-34c-5p inhibitor or inhibitor control was detected by ELISA. (G) Alizarin red staining of BMSCs transfected with miR-34c-5p mimic or mimic control at 14 days. Scales: 200 μm; magnifications: ×200. (H) Alizarin red staining of BMSCs transfected with miR-34c-5p inhibitor or inhibitor control at 14 days. Scales: 200 μm; magnifications: ×200. *vs. MC; ^vs. IC. * or ^p<0.05; ** or ^^p<0.01; *** or ^^^p<0.001. BMSCs, bone marrow mesenchymal stem cells; qRT-PCR, quantitative reverse transcription polymerase chain reaction; CCK-8, cell counting 8; ALP, alkaline phosphatase; ELISA, enzyme linked immune sorbent assay; I, miR-34c-5p inhibitor; IC, inhibitor control; M, miR-34c-5p mimic; MC, mimic control. The experiment was independently repeated three times.
|
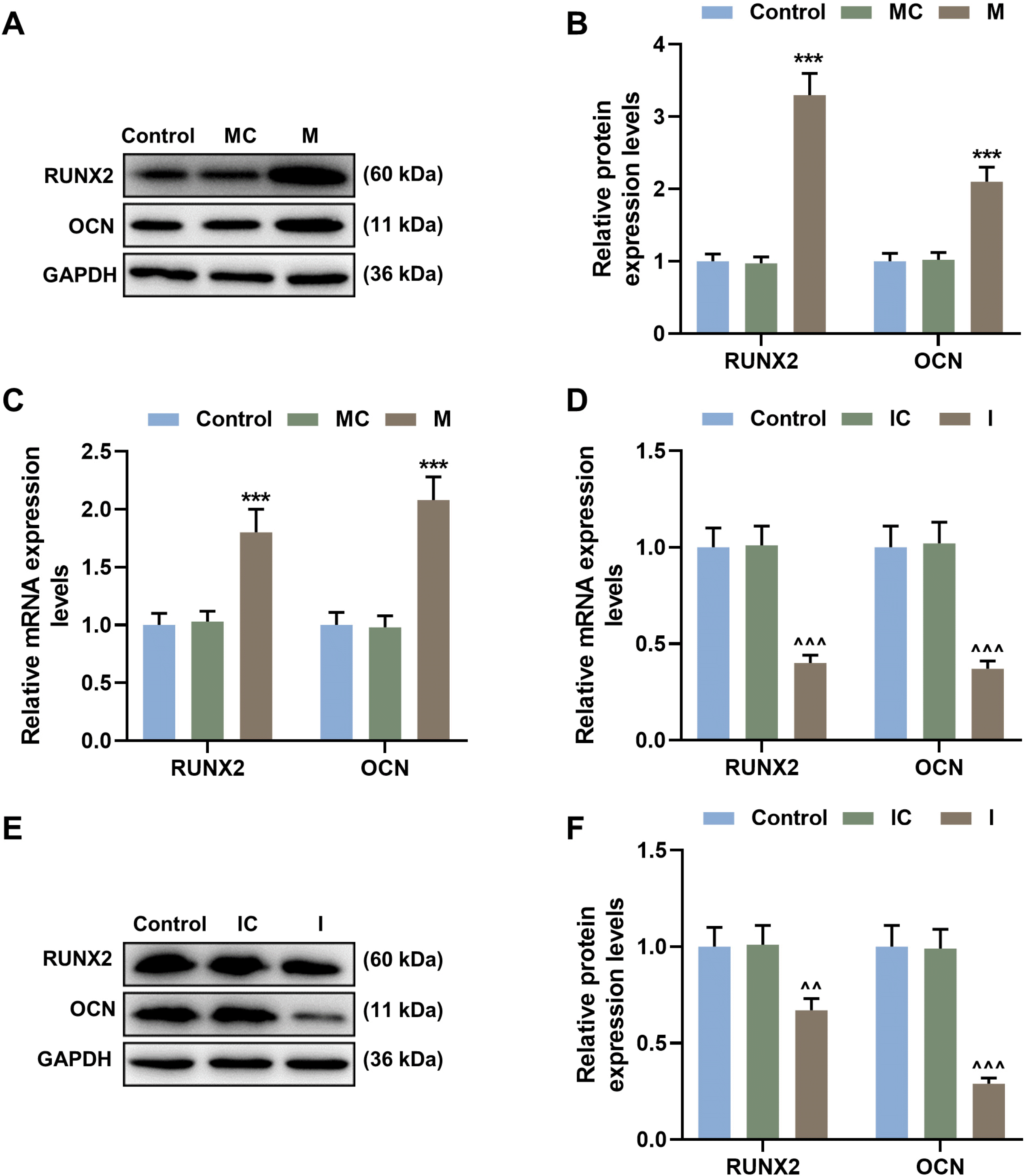 | Fig. 3MiR-34c-5p promoted the expression of RUNX2 and OCN of BMSCs. (A, B) The expression of RUNX2 and OCN of BMSCs transfected with miR-34c-5p mimic or mimic control was detected by Western blot. (C) The expressions of RUNX2 and OCN of BMSCs transfected with miR-34c-5p mimic or mimic control were detected by qRT-PCR. (D, E) The expressions of RUNX2 and OCN of BMSCs transfected with miR-34c-5p inhibitor or inhibitor control were detected by Western blot. (F) The expressions of RUNX2 and OCN of BMSCs transfected with miR-34c-5p inhibitor or inhibitor control were detected by qRT-PCR. *vs. MC; ^vs. IC. ^^p<0.01; *** or ^^^p<0.001. BMSCs, bone marrow mesenchymal stem cells; RUNX2, Runt-related transcription factor 2; OCN, osteocalcin; qRT-PCR, quantitative reverse transcription polymerase chain reaction; I, miR-34c-5p inhibitor; IC, inhibitor control; M, miR-34c-5p mimic; MC, mimic control. The experiment was independently repeated three times.
|
MiR-34c-5p promoted osteogenic differentiation of BMSCs by inhibiting the expression of Bcl-2
TargetScan database is widely used to predict the downstream target genes and targeted binding sites of miRNA (19). In this study, as shown in Fig. 4A, TargetScan predicted the targeted binding sites of miR-34c-5p and Bcl-2. The following dual-fluorescent reporter gene assay results demonstrated that after mutating the targeted binding sites of miR-34c-5p and Bcl-2, up-regulation and down-regulation of miR-34c-5p expression did not affect the fluorescence intensity compared with the control group (Fig. 4B and 4C). However, in the wild-type group, overexpression of miR-34c-5p reduced the fluorescence intensity compared with the control group, while down-regulation of miR-34c-5p showed opposite effects (Fig. 4B and 4C; p<0.05, p<0.001). This suggests that miR-34c-5p could target Bcl-2. CCK-8 assay showed that overexpression of miR-34c-5p decreased the proliferation of BMSCs compared with the control group (Fig. 4D, p<0.05, p<0.01). However, upregulation of Bcl-2 reversed the effect of miR-34c-5p mimics (Fig. 4D; p<0.05, p<0.01, p<0.001). In contrast, miR-34c-5p inhibitor promoted the proliferation BMSCs compared with control group, and siBcl-2 reversed the effect of miR-34c-5p inhibitor (Fig. 4E, p<0.05, p<0.01). Moreover, miR-34c-5p mimic increased the content of ALP and calcium deposition compared with the control group, and overexpression of Bcl-2 reversed the effect of miR-34c-5p mimic (Fig. 4F, p< 0.001). Similarly, the ALP content and calcium deposition decreased by miR-34c-5p inhibitor but were increased by siBcl-2 (Fig. 4G, Fig. 5A and 5B; p<0.001). Furthermore, the expression of RNUX2 and OCN also verified the above results. As shown in the Fig. 5C∼H, miR-34c-5p mimic downregulated the expression of Bcl-2 and upregulated the expressions of RUNX2 and OCN compared with the control group. Meanwhile, overexpression of Bcl-2 reversed the effects of miR-34c-5p mimic. The effects of miR-34c-5p inhibitor were opposite to those of miR-34c-5p mimic, and inhibiting the expression of Bcl-2 could also partially reverse the effects of miR-34c-5p inhibitor.
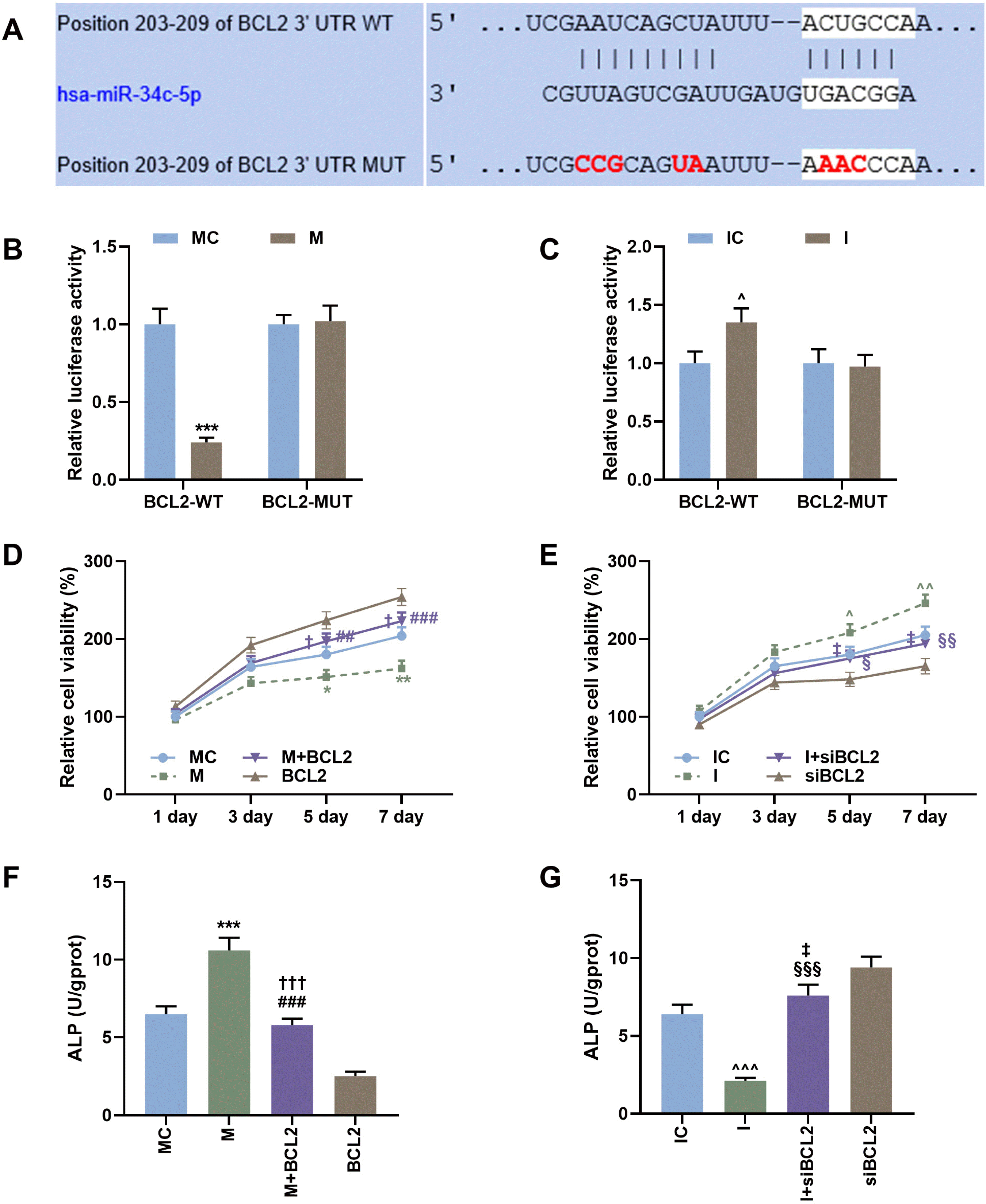 | Fig. 4MiR-34c-5p decreased cell activity and increased ALP content of BMSCs by inhibiting Bcl-2 expression. (A) The binding site of miR-34c-5p and Bcl-2 was predicted by TargetScan. (B, C) Dual-luciferase reporter gene assay was used to verify the targeted binding relationship between miR-34c-5p and Bcl-2. (D) The cell proliferation of BMSCs transfected with or untransfected miR-34c-5p mimic and/or Bcl-2 was detected by CCK-8 at 1, 3, 5 and 7 day. (E) The cell proliferation of BMSCs treated or untreated with miR-34c-5p inhibitor and/or siBcl-2 was detected by CCK-8 at 1, 3, 5 and 7 day. (F) The ALP content of BMSCs treated or untreated with miR-34c-5p mimic and/or Bcl-2 was detected by ELISA. (G) The ALP content of BMSCs treated or untreated with miR-34c-5p inhibitor and/or siBcl-2 was detected by ELISA. *vs. MC, #vs. M, †vs. BCL2, ^vs. IC, §vs. I, ‡vs. siBCL2. † or ^ or ‡ or §p<0.05; ## or ^^ or §§p<0.01; *** or ^^^ or ††† or ###p<0.001. BMSCs, bone marrow mesenchymal stem cells; CCK-8, cell counting 8; ALP, alkaline phosphatase; ELISA, enzyme linked immune sorbent assay; Bcl-2, B cell lymphoma/lewkmia-2; I, miR-34c-5p inhibitor; IC, inhibitor control; M, miR-34c-5p mimic; MC, mimic control. The experiment was independently repeated three times.
|
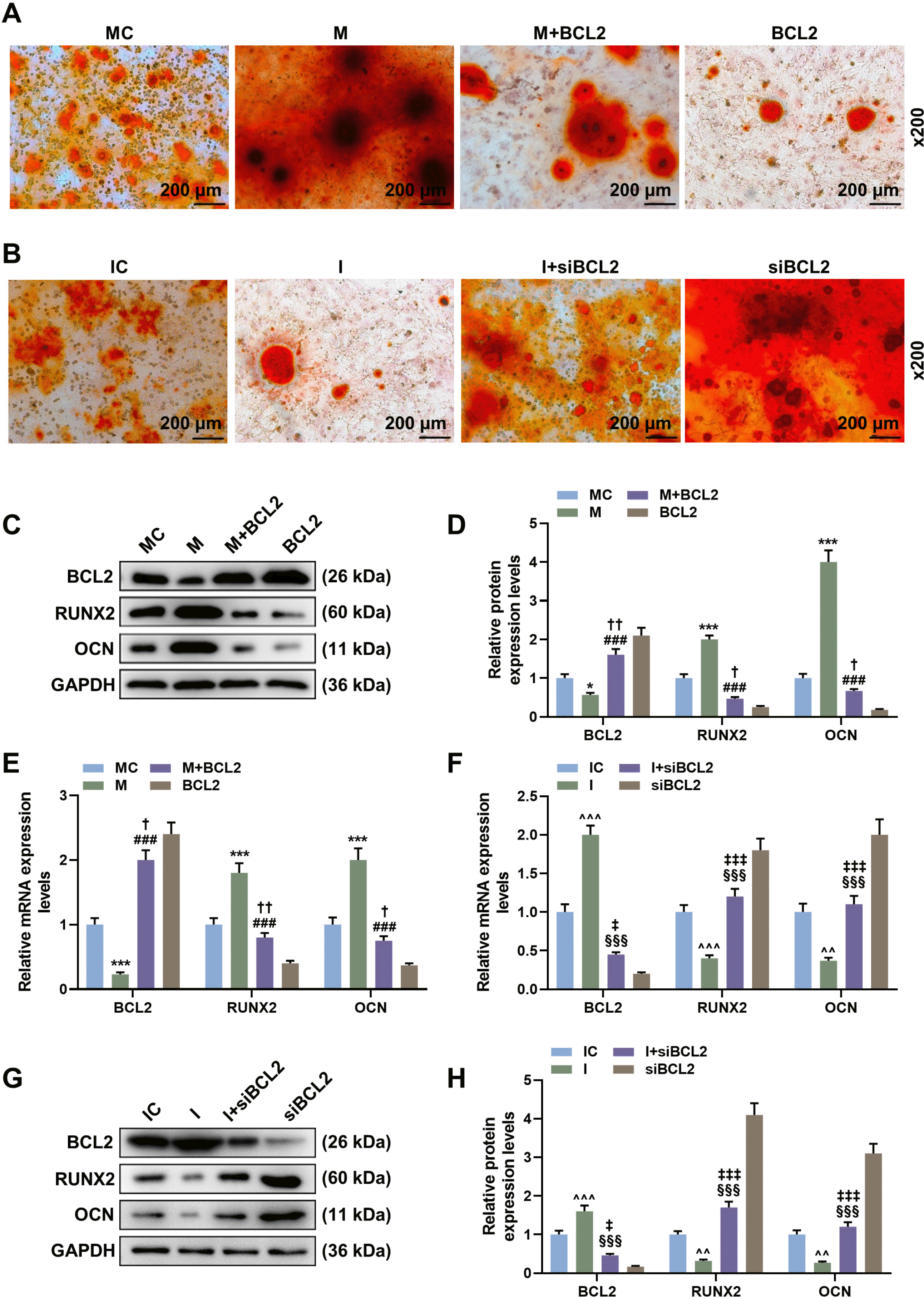 | Fig. 5MiR-34c-5p promoted calcium deposits and the expression of RUNX2 and OCN of BMSCs by inhibiting Bcl-2 expression. (A) Alizarin red staining of BMSCs treated or untreated with miR-34c-5p mimic and/or Bcl-2 at 14 days. Scales: 200 μm; magnifications: ×200. (B) Alizarin red staining of BMSCs treated or untreated with miR-34c-5p inhibitor and/or siBcl-2 at 14 days. Scales: 200 μm; magnifications: ×200. (C, D) The expression of Bcl-2, RUNX2 and OCN of BMSCs treated or untreated with miR-34c-5p mimic and/or Bcl-2 were detected by Western blot. (E) The expressions of Bcl-2, RUNX2 and OCN of BMSCs treated or untreated with miR-34c-5p mimic and/or Bcl-2 were detected by qRT-PCR. (F) The expressions of Bcl-2, RUNX2 and OCN of BMSCs treated or untreated with miR-34c-5p inhibitor and/or siBcl-2 were detected by qRT-PCR. (G, H) The expressions of Bcl-2, RUNX2 and OCN of BMSCs treated or untreated with miR-34c-5p inhibitor and/or siBcl-2 were detected by Western blot. *vs. MC, #vs. M, †vs. BCL2, ^vs. IC, §vs. I, ‡vs. siBCL2. † or * or ‡p<0.05; ^^ or ††p<0.01; *** or ^^^ or ‡‡‡ or ### or §§§p<0.001. BMSCs, bone marrow mesenchymal stem cells; CCK-8, cell counting 8; ALP, alkaline phosphatase; ELISA, enzyme linked immune sorbent assay; Bcl-2, B cell lymphoma/lewkmia-2; I, miR-34c-5p inhibitor; IC, inhibitor control; M, miR-34c-5p mimic; MC, mimic control. The experiment was independently repeated three times.
|
Go to :
Discussion
MiRNAs regulate the expressions of various molecules by inhibiting protein translation and promoting the degradation of mRNAs (20). At present, their important roles in cell cycle regulation, proliferation, differentiation, and apoptosis have attracted much research attention (21). Previous studies showed that multiple miRNAs are regulators of osteogenic differentiation of MSCs (22-24). Therefore, we analyzed the database related to osteogenic differentiation of MSCs through bioinformatics methods, and the obtained data were verified by qRT-PCR to finally obtain the miRNA (miR-34c-5p) with the greatest expression difference between normal rabbit and lumbar fusion model rabbit. Liu et al. (13) found that miR-34c is involved in osteogenic differentiation of BMSCs. In the following experiments, we also observed that the expression of miR-34c-5p was gradually upregulated during the osteogenic differentiation of BMSCs. In addition, our test results proved that ALP content increased gradually during osteogenic differentiation of BMSCs. ALP is a marker enzyme indicative of the level of catabolism in bone tissues, and plays a key role in calcification. Under the action of ALP, calcium ions are deposited on collagen to complete the matrix mineralization process. Bone tissues are formed through calcification of bone matrix, which is synthesized and secreted by osteoblasts (25). The ALP content of osteoblasts, which will reach the highest at the beginning of calcification and the lowest near the end of calcification, could reflect the differentiation degree and functional state of osteoblasts to some extent (26). Further test results showed that calcium deposition gradually increased with the advancement of osteogenic differentiation of BMSCs cells. Notably, calcium deposition eventually induces calcification. It can be concluded that miR-34c-5p is positively correlated with osteogenic differentiation of BMSCs.
In order to further understand whether miR-34c-5p is involved in osteogenic differentiation of BMSCs, miR-34c-5p in BMSCs cells was overexpressed or subtracted. The experimental results showed that miR-34c-5p overexpression alleviated the proliferation of BMSCs, and promoted the ALP content and calcium deposition. Upregulation of miR-34c-5p would increase the expression of RUNX2 and OCN, but silencing miR-34c-5p results in the opposite effect. Notably, RUNX2 is the most important transcription factor for inducing MSCs to osteoblasts (27). The mechanisms of osteoblast differentiation regulated by RUNX2 are complex and involve many types of post-translational modifications (28). It has reported that miR-34c inhibits osteoblast differentiation of MC3T3-E1 premature osteoblast by directly targeting Runx2 (29, 30). Runx2 also interacts with other factors, such as alkaline phosphatase (ALP), osterix (OSX), and osteocalcin (OCN), to activate genes related to osteoblast differentiation (31). OCN is an osteoblast-specific protein that promotes calcium and phosphorus deposition (32). The above results indicated that miR-34c-5p is involved in the osteogenic differentiation process of BMSCs, and provided evidence that miR-34c-5p affects the proliferation of osteoblasts.
In this study, TargetScan predicted that miR-34c-5p could bind to Bcl-2 gene. The regulatory relationship between miR-34c and Bcl-2 has been previously detected in many diseases. For example, miR-34c increases the sensitivity of osteosarcoma cells to cisplatin by inhibiting Bcl-2 (33); injection of Bcl-2 protein into mouse fertilized oocytes enhances the effect of miR-34c inhibitor (34). However, the regulatory effect between miR-3c-5p and Bcl-2 has not been verified. Bcl-2 gene is a widely recognized apoptosis-related protein and is often considered as an oncogene (35-37). Endothelial progenitor cell–derived microvesicles promoted proliferation and migration of MC3T3-E1 cell and reduced apoptosis through Erk1/2-Bcl-2 pathway (38), indicating that upregulated Bcl-2 could inhibit apoptosis and promote proliferation. In addition, Bcl-2, which is expressed in osteoblasts, osteoclasts, chondrocytes, and osteocytes (39), plays an important role in maintaining bone integrity (40). Bone-targeted overexpression of Bcl-2 increases osteoblast adhesion and differentiation and inhibits mineralization in vitro (41). However, the role of Bcl-2 in affecting the osteogenic differentiation of BMSCs should be further explored. Interestingly, recent studies indicated that Bcl-2 is associated with calcium deposition (42). Bcl-2 promotes the differentiation, activity, and survival of both osteoblasts and osteoclasts (43), but study also reported that overexpression of Bcl2 inhibited the osteogenic/odontogenic potential of DPSCs, as indicated by lower levels of ALP activity and mineralized calcium deposition (17). Our study found that Bcl-2 promoted the proliferation of BMSCs, reduced ALP content and calcium deposition, and suppressed RUNX2 and OCN expression. Bcl-2 overexpression could partially reverse the regulatory effects of miR-34c-5p on ALP content, calcium deposition and RUNX2 and OCN expression. The above results indicated that Bcl-2 could affect the proliferation and osteogenic differentiation of BMSCs, and these effects are regulated by miR-34c-5p.
This study revealed that miR-34c-5p promotes the osteogenic differentiation and inhibits the proliferation of BMSCs by inhibiting the expression of Bcl-2. As human BMSCs are an important source of osteoblasts, we speculate that the expression of miR-34c-5p might be an important mechanism underlying the success of lumbar fusion surgery with autogenous bone graft.
Go to :
 XML Download
XML Download