

 PDF
PDF Citation
Citation Print
Print


Introduction
Hepatocellular carcinoma (HCC), a major form of liver cancer, ranks the second leading cause of cancer related death globally (1). Nowadays, surgical resection combined with radiotherapy, chemotherapy and biotherapy have maintained the mainstream for HCC therapy strategy (2). However, the majority of HCC patients are diagnosed with advanced liver cancer due to an asymptomatic early stage, which results in the unavoidable systemic metastasis when surgery option is no longer available (3). Additionally, resistances to radiotherapy and chemotherapy always occur in HCC patients (4).
Cancer stem cells (CSCs) are the specific subpopula-tions of cells with stem cell-like characteristics in tumor tissues (5). CSCs have the potentials of self-renewal and differentiation, which may be responsible for the recurrence and metastasis of HCC (6). Bone mesenchymal stem cells (BMSCs) are cells derived from pluripotent bone marrow, characterized by easy obtaining and culture, and vigorous proliferation ability (7). Emerging evidence demonstrate the implication of BMSCs in the development and progression of various cancers via interacting with tumor cells (8, 9), but how BMSCs works in tumor microenvironment remains much to be determined.
MicroRNAs (miRNAs) belong to the small non-coding RNAs with 20∼25 nucleotides in length. MiRNAs can degrade or inhibit the translation of the targeted gene by binding to the 3’ untranslated region (UTR) of mRNA in various conditions (10). Among which, microRNA 140 (miR-140) has been documented as a tumor suppressor in HCC (11-13). The association of miR-140 with BMSCs was proved in chondrocyte cells (14), but whether this association still stands in HCC remains unclear.
Traditional Chinese Medicine (TCM) has been used in the adjuvant treatment of HCC with satisfactory therapeutic efficiency (15, 16). Bie Jia Jian pill (BJJP) is a kind of TCM from “the Synopsis of Prescription of the Golden Chamber”. BJJP has been used for the therapy of liver fibrosis or hepatitis B cirrhosis (17). A previous studies highlighted the importance of BJJP on suppression HCC progression, whose mechanism involves MAPK signal pathway (18). According to collected data, 80∼90% of HCC patient develops from liver fibrosis or cirrhosis (19). However, the effects of BJJP on HCC are unknown. In this present study, BJJP and BMSCs were used to treat CSCs enriched from Huh7 cells to investigate their effects on CSCs. This study identified that both BJJP and BMSCs can suppress the CSCs viability. In addition, BJJP and BMSCs together could further exert their protective effect against CSCs through regulating miR-140 and Wnt/β-catenin signal pathway.
Go to : 
Materials and Methods
Animals
A total of 26 male BALB/c mice (18±2 g) provided by the Hubei Provincial Laboratory Animal Public Service Center were housed in the specific pathogen free (SPF) animal room under the 12 h light/dark cycles at 24∼26℃ with free access to food and water. All experiments were conducted in accordance with the Guidelines for the Institutional Animal Care and Use Committee (IACUC) and approved by the Committee of The First Affiliated Hospital of Guangxi University of Chinese Medicine.
Enrichment of stem cell-like Huh7 spheres
Human HCC cells (Huh7) were purchased from iCell (Shanghai, China). First, Huh7 cells were cultured in high glucose DMEM medium (iCell, Shanghai, China) supplemented with 10% fetal bovine serum (FBS) at 37℃ in a humidified incubator with 5% (v/v) CO2. After digestion and centrifugation, Huh7 cells were resuspended with stem cell conditioned culture medium [low glucose DMEM-F12 medium containing B27 (1:50), 0.4% BSA, EGF (20 ng/ml) and bFGF (20 ng/ml)]. Then, the cells were plated into 24 well ultra-low attachment plates with serum-free stem cell conditioned culture medium to form the spheres. The spheres were collected after 8 days. Sphere-like cells were trypsinized into single cell suspension for follow-up experiments.
Isolation and culture of BMSCs
Male BALB/c mice (n=6) were sacrificed by cervical dislocation. Then mice were then soaked in 75% (v/v) ethanol for 10 min. Under aseptic condition, the femur and tibia were removed. After the epiphysis at both ends was removed, BMSCs in the bone marrow cavity were washed out with low-glucose DMEM. The culture medium containing cell suspension was centrifuged at low speed for 5 min and the supernatant was discarded, and then resuspended with low-glucose DMEM complete culture medium. The collected cells were cultured at 37℃ in a humidified incubator with 5% (v/v) CO2. After incubation for 48 h, the non-adherent cells were removed. When confluence was 80%∼90%, the adherent cells were harvested by trypsin digestion and designated BMSCs.
Cell transection
miR-140 overexpression cells were established by transfecting miR-140 mimic into CSCs using the lipofectamine 3,000 reagent (Invitrogen, USA) according to the manufacturer’s instructions. MiR-140 knockdown cells were established by transfecting miR-140 inhibitor into CSCs using the lipofectamine 3,000 reagent (Invitrogen, USA) according to the manufacturer’s instructions. Sequences for miRNAs transfection were shown as follows: miR-140 mimic NC: TTCTCCGAACGTGTCACGT; miR-140 mimic: CAGTGGTTTTACCCTATGGTAG; miR-140 inhibitor NC: CAGTACTTTTGTGTAGTACAA; miR-140 inhibitor: CTACCATAGGGTAAAACCACTG.
Preparation of drug-containing serum
Male BALB/c mice (n=20) received intragastric administration of saline (0.9%) once a day for continuous 15 days. The other 10 BALB/c mice received intragastric administration of BJJP (1.1 g/kg, Sinopharm Zhonglian, Wuhan, China) once a day for continuous 15 days. After the mice were anesthetized with pentobarbital sodium, the blood was collected from the carotid artery and centrifuged at 4℃ for 10 min (3,000 r/min). The obtained serum was inactivated at 56℃ for 20 minutes, filtered and sterilized at 0.22 μm, and stored at 4℃.
Cell culture and treatments
Using the transwell chamber as the co-culture system, BMSCs and CSCs were directly co-cultured in a non-touch manner. Briefly, CSCs with different treatments were inoculated in the lower chamber with or without serum containing BJJP. BMSCs were inoculated in the upper chamber. After cultured in a humidified incubator with 5% (v/v) CO2 at 37℃ for 7 days, CSCs in the lower chamber were used for the subsequent experiments.
Measurement on surface antigen
Flow cytometry analysis was conducted to evaluate the cell surface marker expressions of the CSCs. CSCs with various treatments were harvested and trypsinized into a single cell suspension. Subsequently, cells were incubated with fluorochrome-conjugated antibodies, including FITC-conjugated anti-human cluster of differentiation (CD) 24 (eBioscience, CA, USA), APC-conjugated anti-human CD133 (Biolegend, CA, USA) at 4℃ in the dark. The appropria-ted isotype-matched antibodies were used as negative controls. After 30 min incubation, the samples were centrifuged and washed with PBS for flow cytometry analysis. CSCs with positive expressions of both CD24 and CD133 were selected for further experiments. Similarly, the cell surface marker expressions of the BMSCs were also evaluated by flow cytometry. The conjugated-specific antibodies used were as follows: PE-conjugated anti-mouse CD14 (Biolegend, CA, USA), APC-conjugated anti-mouse CD34 (Biolegend, CA, USA), APC-conjugated anti-mouse CD45 (Biolegend, CA, USA), PE-conjugated anti-mouse CD44 (Biolegend, CA, USA), PE-conjugated anti-mouse CD73 (Biolegend, CA, USA), APC-conjugated anti-mouse CD105 (Biolegend, CA, USA), FITC-conjugated anti-mouse CD166 (abcam, CA, USA), FITC-conjugated anti-mouse CD29 (Biolegend, CA, USA). Cell analysis was performed using a Flow cytometer (BD Biosciences, CA, USA) and FolwJo software version 7.6 (FlowJo LLC, OR, USA).
Differentiation ability of BMSCs
Adipogenic differentiation by oil red O staining: BMSCs (2×104 cells/cm2) were inoculated into the 6 well plates for cell fusion. Adipogenic differentiation was induced with adipogenic differentiation basal medium (MUCMX-90031, Cyagen, Guangzhou, China) when cell fusion reaches 100%. BMSCs were added with 2 ml of culture medium for OriCell BALB/C BMSCs culture medium (liquid A) after the original culture medium was removed. After adipogenic differentiation for 3 day, the liquid A in the 6 well plates was removed and replaced with 2 ml of culture medium for OriCell BALB/C BMSCs culture medium (liquid B) for cell culture for 24 h. Then the liquid B was absorbed and replaced with liquid A. The inter-replacement of liquid A and liquid B was performed for 3∼5 times (12∼20 days), the BMSCs were cultured with liquid B for 4∼7 day (refresh every 2∼3 days) until the lipid droplets are big enough. The 6 well plates were washed with 1×PBS for 1∼2 times and fixed with 2 ml 4% neutral formalin solution per well for 30 min. Then the formalin solution was removed and the 6 well plates were washed with 1×PBS for twice. After PBS wash, each well was added with 1 ml of oil red O dye liquor for 30 min (dye liquor formulate: oil red O:distill water=3:2, filtered through the neutral filter paper). After the oil red O dye liquor was removed, the 6 well plates were washed in 1×PBS for 2∼3 times and observed under a micro-scope.
Osteogenic differentiation and alizarin red staining: BMSCs of 2×104 cell/cm2 were seeded into the gelatin coated 6 well plates. Osteogenic differentiation was induced with Osteogenic differentiation culture medium (MUCMX-90031, Cyagen, Guangzhou, China) when cell fusion reaches 60∼70%. After the culture medium was removed, the 6 well plates were added with culture medium for 2 ml OriCell BALB/C BMSCs (refresh every 3 days) for 2∼4 weeks. Alizarin red was used for cell staining. After staining, the culture medium was removed and the 6 well plates were washed with 1×PBS for 1∼2 times, followed by fixation with 2 ml 4% neutral formalin solution per well for 30 min. Then the formalin solution was removed and the 6 well plates were washed with 1×PBS for twice. After PBS wash, each well was added with 1 ml of alizarin red. About 30 min later, the 6 well plates was washed with 1×PBS for 2∼3 times and then observed under a microscope.
CCK8 assay
Cell counting kit-8 (CCK-8) assay was conducted to evaluate the cell viability of CSCs. A total of 10 μl of CCK-8 reagent (Beyotime, Shanghai, China) was added into each well and then incubated for another 2 h. The absorbance of each well at 450 nm was assessed using a microplate reader (Diatek, Wuxi, China).
Clone formation assay
Cells in logarithmic phase were digested with 0.25% trypsin and made into single cells for cell suspension in culture medium containing 10% fetal calf serum. Cell suspensions were seed into pre-warmed culture disk which contains 10 ml of culture medium at the density of 50, 100 and 200 cells per disk. Cells were gently shaken and then maintained at 37℃ environment with 5% saturated humidity for 2 to 3 weeks. Culture was terminated once cell clones were visible by naked eyes. The cell suspension was removed and the cells were washed in PBS for twice and fixed in 5 ml of acetic acid/methanol (1:3) for 15 min. After that, cell clones were stained with Giemsa staining for 10 to 30 min and washed in running water. The air dryed culture disks were inversed and counted for cell clones. Cell clones with more than 10 cells were counted to calculate the cell clone rate.
Cell apoptosis assay
Cell apoptosis of CSCs was determined using Annexin V cell apoptosis kit (Sungene Biotech, Tianjin, China) following the manufacturer’s protocol. The harvested CSCs were washed with PBS and resuspended in the Annexin V binding buffer. Subsequently, cells were incubated with 5 μl of Annexin V-PE and 5 μl of 7-AAD solution at room temperature for 15 min in the dark and analyzed by flow cytometry (Becton Dickinson Co., CA, USA).
Fluorescent quantitative polymerase chain reaction
Total RNA was extracted from the harvested CSCs using TRIpure Total RNA Extraction Reagent (ELK Biote-chnology, Wuhan, China) following the manufacturer’s protocol. Reverse-transcription was conducted using EntiLinkTM 1st Strand cDNA Synthesis Kit (ELK Biotechnology, Wuhan, China). Next, the synthesized cDNA was amplified using the EnTurboTM SYBR Green PCR SuperMix Kit (ELK Biotechnology, Wuhan, China). Relative expression levels of the targeted gene were calculated using the 2−ΔΔCt method. For CD24, CD133 and epithelial cell adhesion molecule (EpCAM), GAPDH served as the internal control. For miR-140, U6 served as the internal control. The sequences of PCR primers are listed in Table 1.
Table 1
The primer sequences used in this study
![]()
Western blot
The harvested CSCs were lysed with RIPA Lysis Buffer (Aspen, Canada, USA) containing the protease inhibitor cocktail (Roche, Basel, Switzerland), followed by centrifugation at 12000 rpm at 4℃ for 5 min. Protein concentration was determined by BCA protein kit (Aspen, Canada, USA). Total protein was electrophoresed on 10% SDS-PAGE and then transferred to PVDF membrane. Subsequently, the membrane was incubated with the primary antibodies overnight at 4℃, followed by the corresponding horseradish peroxidase conjugated secondary antibodies at room temperature for 30 min. GAPDH served as the internal control. Protein bands were captured by enhanced chemiluminescence (ECL) substrate solution kit (Aspen, Canada, USA) for 1 min and analyzed with AlphaEaseFC software. Primary antibodies used in this study are listed in Table 2.
Table 2
The primary antibodies used for western blot analyses in this study
![]()
TOP-Flash/FOP-Flash luciferase assay
Cells in logarithmic phase were seeded in 24-well plates at the density of 1×105/ml for cell culture for 24 h. The mixture of TOP/FOP Flash and pRL-TK plasmid (50:1) was transfected into cells for cell culture of 48 h before cell lysis at room temperature of 15 min. The FLuc and RLuc solution was prepared based on the instruction of kit (GeneCopoeia, Guangzhou, China). The luciferase activity was measured at the ultra-sensitive tubular chemiluminescence apparatus (Berthold, Stuttgart, Germany). The ratio of TOP Flash/FOP Flash should be calculated to reflect the luciferase activity.
Statistical analysis
All experiments were repeated three times and all data were presented as mean±standard deviation (SD). Com-parisons between multiple groups were analyzed by One-way ANOVA analysis using the GraphPad Prism 7.00 software (GraphPad Software, Inc., CA, USA). Data between two groups were analyzed using Student’s t test. p<0.05 was considered statistically significant.
Go to :
Results
Identification of CSCs and BMSCs
Huh7 cells were cultured with stem cell conditioned medium and formed large spheres after 8 days (Fig. 1A). Flow cytometry showed that both spheres and Huh7 cells had positive expression of specific stem cell markers CD24 and CD133. The expression rate of CD24 and CD133 in spheres was 40.3% (Fig. 1B), while that in Huh7 cells was less than 5% (Fig. 1C). Above results suggest the successful isolation of CSCs. Flow cytometry showed that the isolated BMSCs were positive for surface antigens CD44, CD73, CD105, CD166 and CD29, while negative for antigens CD14, CD34 and CD45, indicating the successful isolation of BMSCs (Fig. 1D). Staining by oil red O staining and alizarin red staining in Fig. 1E and 1F supported the potential of osteogenic differentiation and adipogenic differentiation ability of BMSCs. Collec-tively, isolated CSCs and BMSCs are qualified for further experiments.
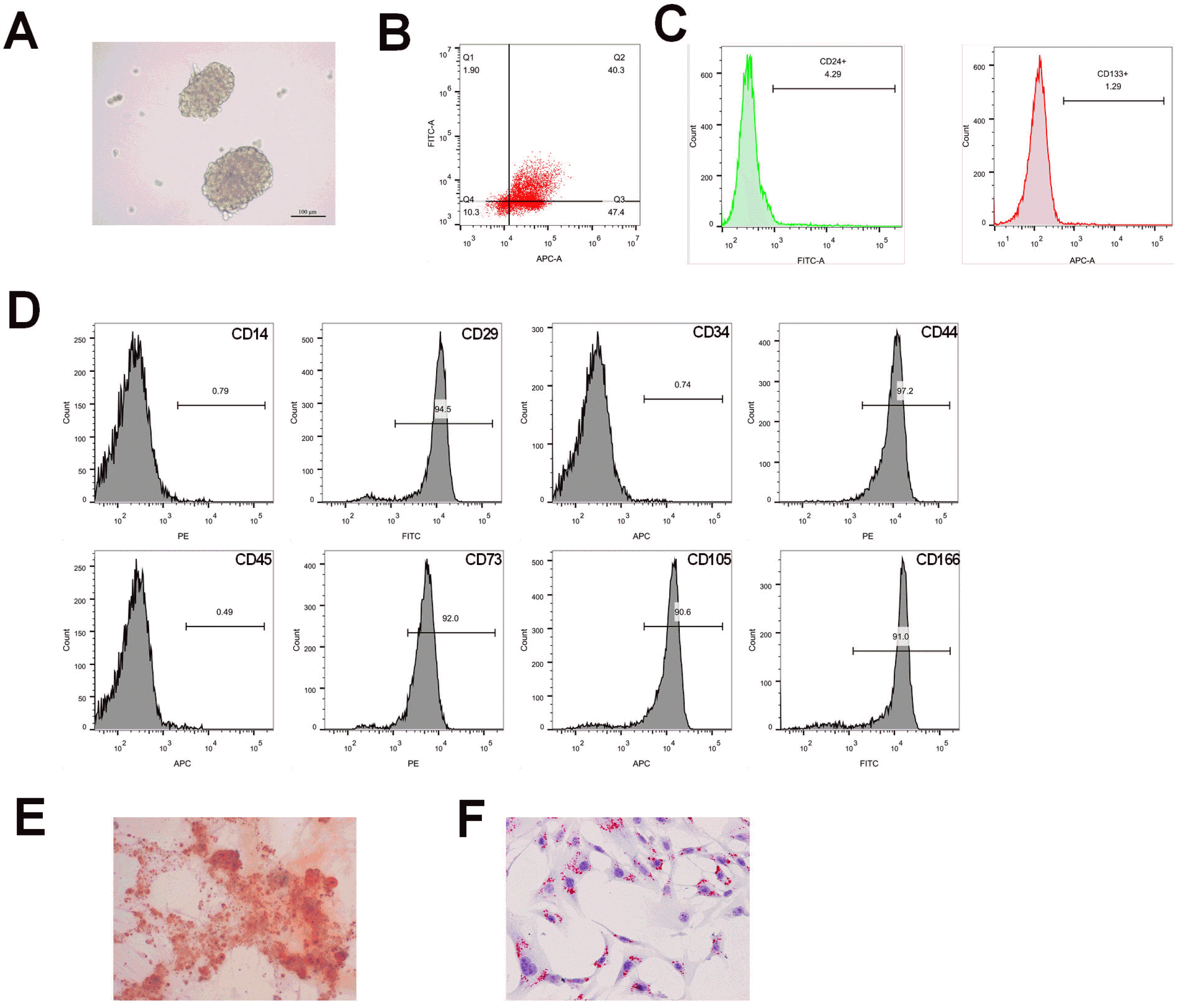 | Fig. 1Identification of isolated CSCs and BMSCs and their potential of osteogenic differentiation and adipogenic differentiation ability. (A) CSCs sphere; (B, C) measurement on CSCs biomarker, CD24 and CD133 by Flow cytometry in spheres and in Huh 7 cells; (D) surface antigens of BMSCs, CD44, CD73, CD105, CD166, CD29, CD14, CD34 and CD45 were measured by Flow cytometry; (E) alizarin red staining for determination of osteogenic differentiation; (F) oil red O staining for determination of adipogenic differentiation. CSCs, cancer stem cells; BMSCs, bone mesenchymal stem cells; BJJP, Bie Jia Jian pill.
|
BJJP+BMSCs inhibits the viability of CSCs and promotes cell apoptosis
Previous study supported the implication of BMSCs in tumor development (8, 20). Therefore in this study we further explore the possible effect of BJJP or/and BMSCs in CSCs. To this end, we used BJJP or BMSCs to treat CSCs before cell proliferation and apoptosis rate were calculated. The comparison showed that compared with control group, CSCs biomarker CD24, CD133 and EpCAM were suppressed (Fig. 2A and 2B, p<0.05) in BJJP and BMSCs groups. Meanwhile, we also found suppressed cell viability and proliferation (Fig. 2C and 2D, p<0.05) as well as increased cell apoptosis rate (Fig. 2E, p<0.05) in BJJP and BMSCs groups in contrast to control group. Collectively, both BJJP and BMSCs can suppress the viability and enhance cell apoptosis of CSCs.
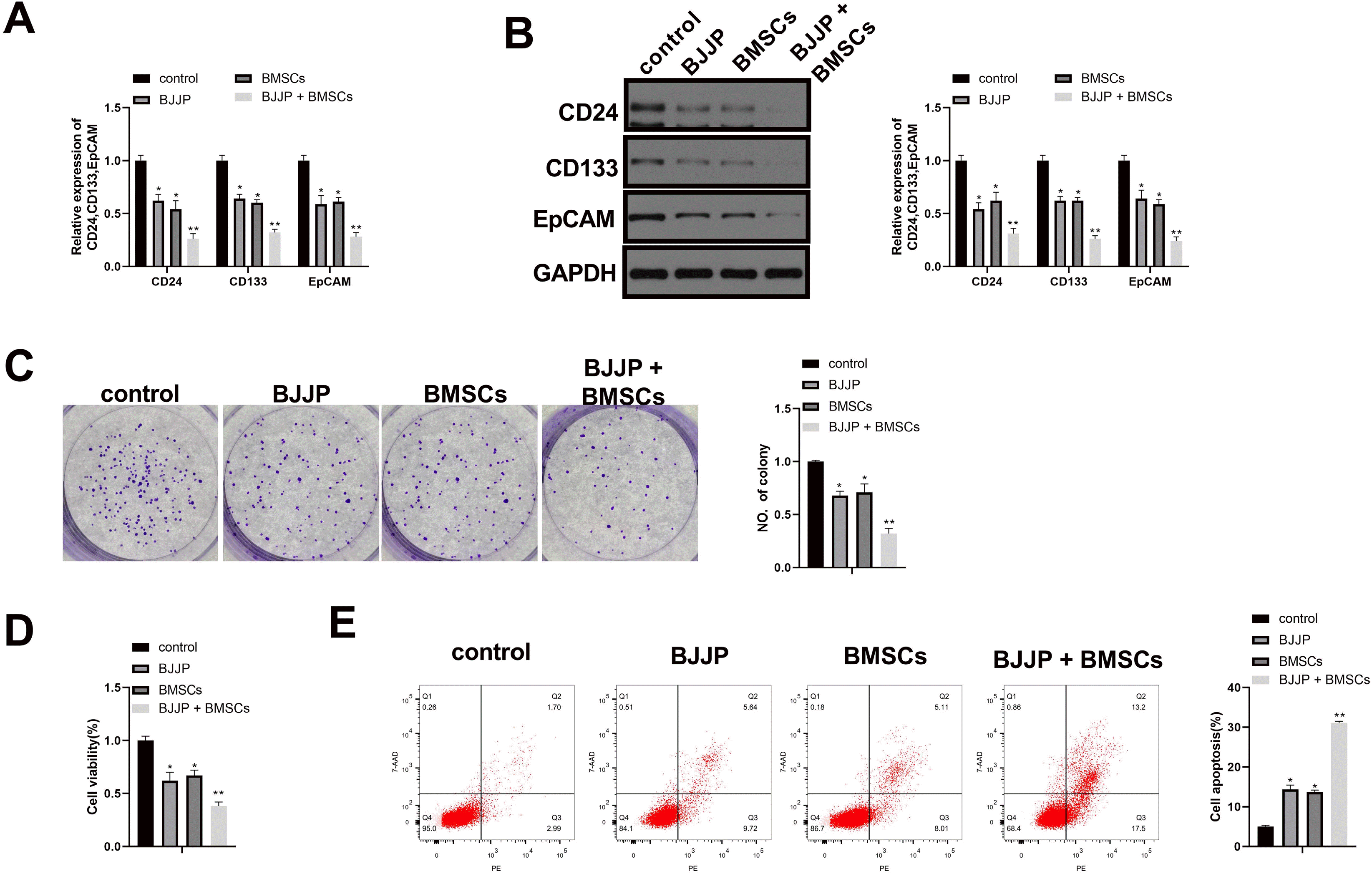 | Fig. 2Combination use of BJJP and BMSCs further suppress the cell growth of CSCs in addition to enhancing cell apoptosis. (A, B) the expressions of CSCs biomarkers CD24, CD133 and EpCAM were measured with RT-qPCR and western blot; (C) cell proliferation ability was assessed by cell clone formation assay; (D) cell viability was determined by CCK-8 assay; (E) cell apoptosis rate was assessed by Flow cytometry. N=3, *p<0.05, **p<0.01; EpCAM, epithelial cell adhesion molecule; CSCs, cancer stem cells; BMSCs, bone mesenchymal stem cells; BJJP, Bie Jia Jian pill.
|
To verify the possible interaction of BJJP and BMSCs in CSCs, CSCs were co-treatment with BMSCs and BJJP. The results showed that compared with BJJP group or BMSCs group, the CD24, CD133 and EpCAM were suppressed while cell viability and proliferation ability of CSCs were suppressed in BJJP+BMSCs group (Fig. 2A∼D, p<0.05). We also found increased cell apoptosis rate in BJJP+BMSCs group when compared with BJJP group or BMSCs group (Fig. 2E, p<0.05). Collectively, combination use of BJJP and BMSCs could further suppress the cell growth of CSCs and promote cell apoptosis.
BJJP and BMSCs promoted the expression of miR-140 in CSCs
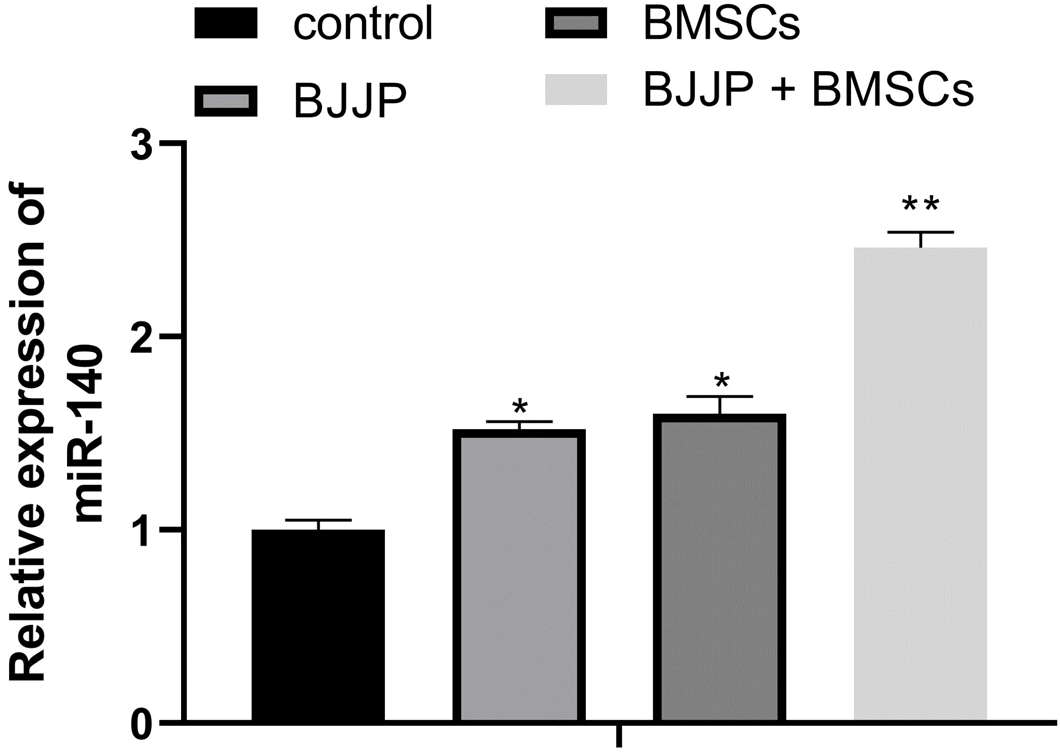
Based on previous research, miR-140 was believed to suppress the proliferation of CSCs with the potential of serving as a tumor therapeutic target (21, 22). RT-qPCR demonstrated that compared with control group, the Huh7 cells had much higher expression level of miR-140 (Fig. 3, p<0.05). We measured the expression of miR-140 in CSCs with or without BJJP/BMSCs treatment. The analysis on miR-140 expression showed that compared with control group, BJJP or BMSCs treatment alone could increase the expression of miR-140, while the expression of miR-140 was further elevated in BJJP+BMSCs group when compared with BJJP or BMSC group (Fig. 3, p<0.05). Collectively, both BJJP and BMSCs could upregulate miR-140 in CSCs.
BJJP and BMSCs suppress CSCs through up-regulating miR-140
miR-140 inhibitor was transfected into CSCs to verify whether miR-140 was involved in the suppressive effect of BJJP and BMSCs in CSCs. The transfection efficiency by RT-qPCR showed that miR-140 was downregulated in miR-140 inhibitor group when compared with inhibitor-NC (Fig. 4A, p<0.05), suggesting the transfection efficiency was satisfactory. Detection on CSCs transfected with miR-140 inhibitor showed that inhibition on miR-140 could partially abandon the suppressive effect of BJJP or BMSCs on cell growth of CSCs (Fig. 4B∼F, p<0.05). Collectively, BJJP and BMSCs could up-regulating miR-140 expression to suppress the cell growth of CSCs.
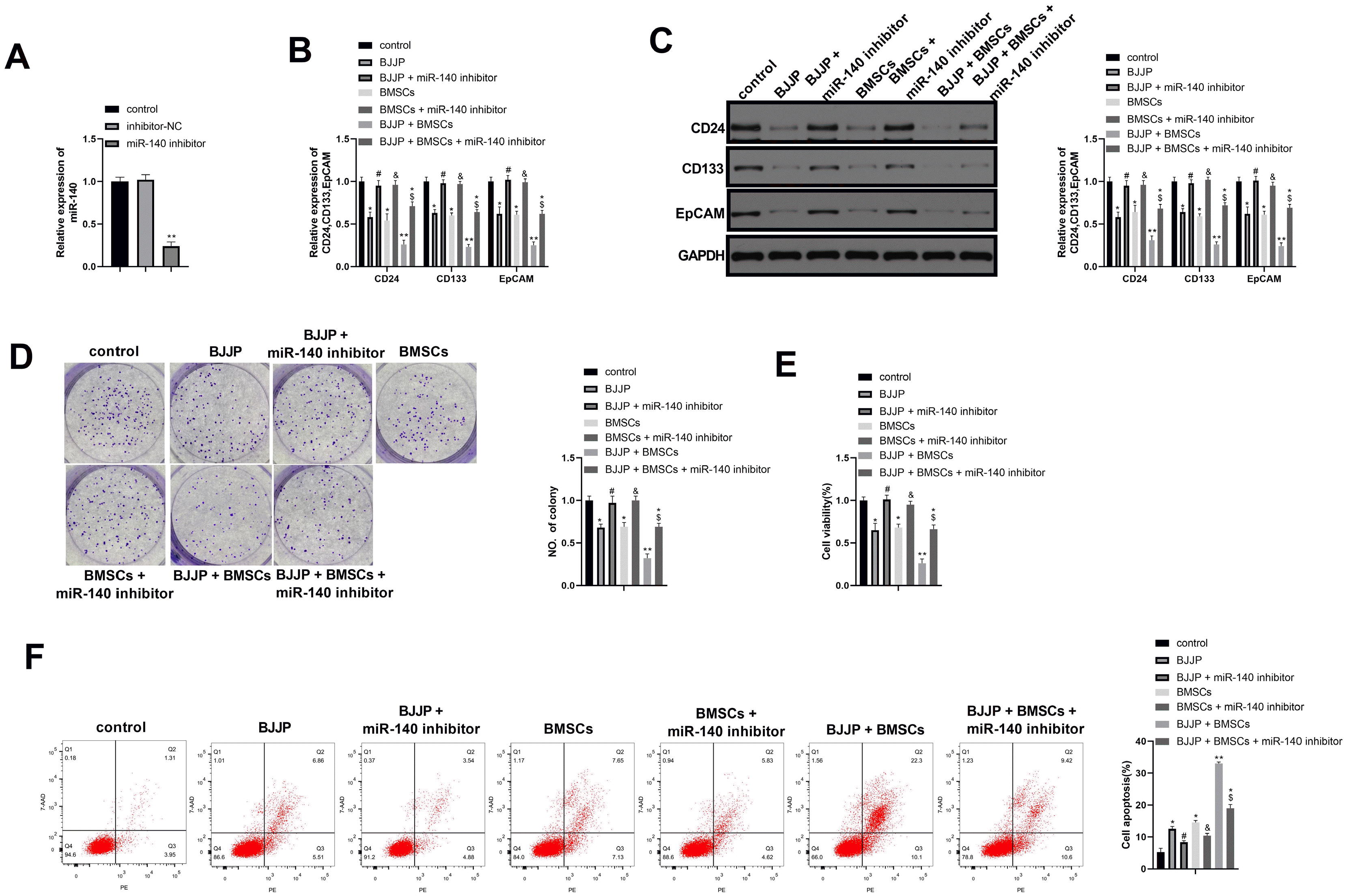 | Fig. 4BJJP and BMSCs suppress CSCs cell proliferation and enhance cell apoptosis through regulating miR-140. (A) Transfection efficiency was verified by RT-qPCR; (B, C) RT-qPCR and western blot were applied to detect the expression of CD24, CD133 and EpCAM after cell transfection; (D) cell proliferation ability was assessed by cell clone formation; (E) cell viability was measured by CCK-8 assay; (F) cell apoptosis was determined by Flow cytometry. N=3, *, vs control group, p<0.05; **, vs control group, p<0.01; #, vs BJJP group, p<0.05; *, vs BMSCs group, p<0.05; *, vs BJJP+BMSCs group, p<0.05; EpCAM, epithelial cell adhesion molecule; CSCs, cancer stem cells; BMSCs, bone mesenchymal stem cells; BJJP, Bie Jia Jian pill.
|
BJJP and BMSCs inhibits Wnt/β-catenin signal pathway in CSCs
Wnt/β-catenin signal pathway plays a vital role in regeneration and renewal of CSCs (23, 24). Western blot was applied to detect the expressions of Wnt/β-catenin signal pathway related proteins (Fig. 5A). In contrast to control group, BMSCs or BJJP treatment lead downregulated expressions of Wnt3a and β-catenin, meanwhile, BMSCs+BJJP group had further decreased Wnt3a and β-catenin expression when compared with BMSCs or BJJP treatment alone (p<0.05). However, knockdown of miR-140 could rescue the expressions of Wnt3a and β-catenin (p<0.05). Luciferase assay in Fig. 5B on activity of β-catenin/TCF was consistent with that of β-catenin. Collectively, BJJP and BMSCs could upregulate miR-140 and suppress Wnt/β-catenin signal pathway to suppress the cell growth of CSCs.
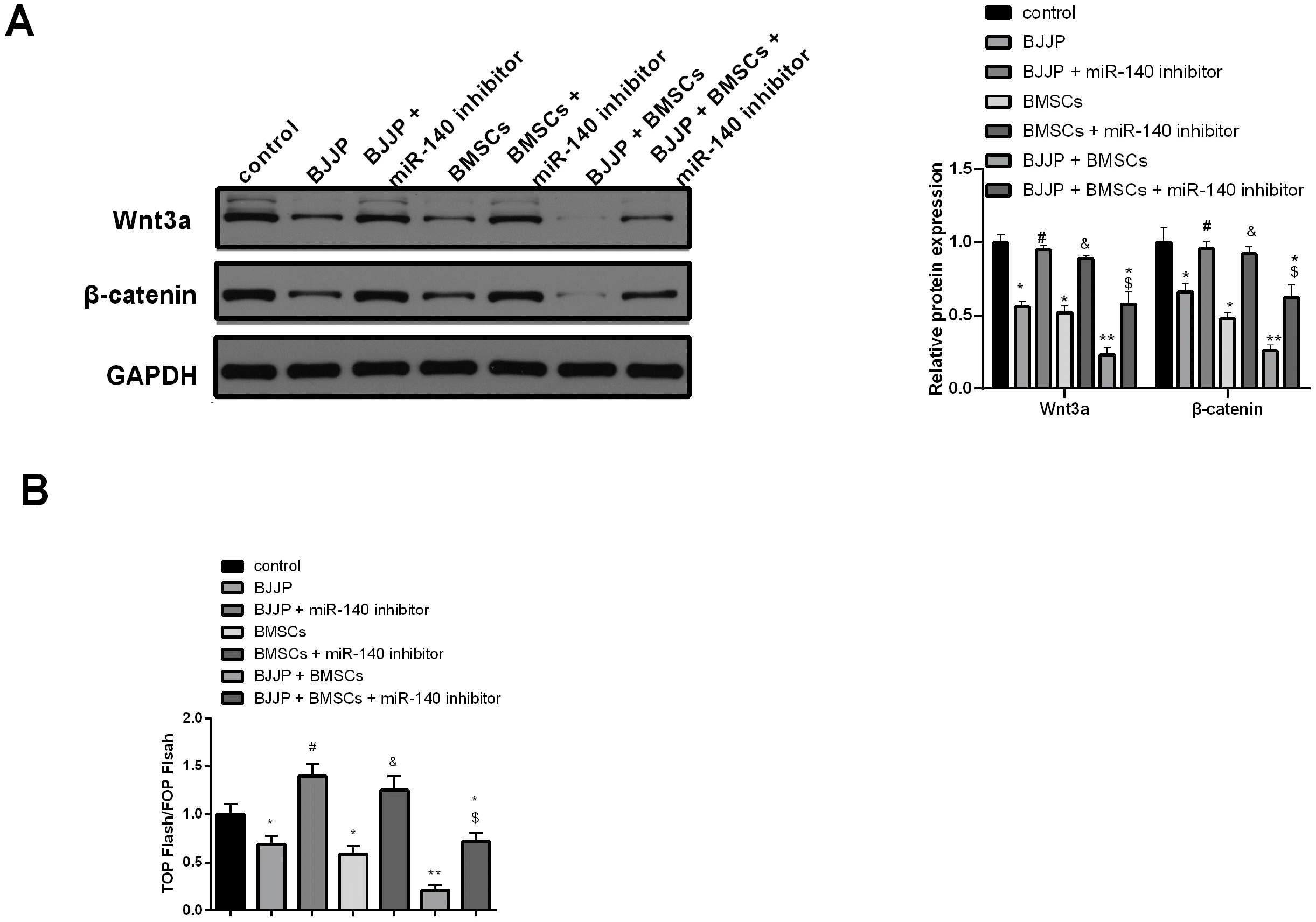 | Fig. 5BJJP and BMSCs could suppress the activation of Wnt/β-ca-tenin signal pathway through regulating miR-140. (A) Western blot was applied to measure the proteins related to Wnt/β-catenin signal path-way. (B) The activity of β-catenin was measured by TOP-Flash/FOP-Flash luciferase assay. N=3, * vs control group, p<0.05; ** vs control group, p<0.01; # vs BJJP group, p<0.05; * vs BMSCs group, p<0.05; *, vs BJJP+BMSCs group, p<0.05. BMSCs, bone mesenchymal stem cells; BJJP, Bie Jia Jian pill.
|
Go to :
Discussion
HCC is a kind of cancer with high mortality and high recurrence rate due to the advanced diagnosis, drug resistance and other factors. In this study, we initially explored the possible role of BJJP in the progression of HCC. The collected evidence in this study showed that both BJJP and BMSCs can suppress the viability of CSCs and enhance cell apoptosis in HCC cell models. Furthermore, we also found the combination use of BJJP and BMSCs could enhance the suppressive effect on CSCs proliferation, among which the mechanism involves the upregulation of miR-140 and suppression of Wnt/β-catenin signal pathway.
Mounting evidence showed that CSCs are associated with cancer recurrence, metastasis, therapy resistance and poor prognosis (25, 26). Lin et al. (27) have found that liver CSCs with tumorigenic potential shall be responsible for relapse and therapy resistance in HCC. CD24, CD133 and EpCAM are three markers of CSCs, which are usually related to the poor prognosis of various cancers (5). EpCAM, a hepatic CSCs biomarker, has been demonstrated to be implicated in poor clinical outcomes of HCC (28). In this study, the expressions of CD 24, CD133 and EpCAM were detected in CSCs enriched from Huh7 cells, which indicated that these cells possessed the potential of stem cells. We found that BJJP markedly reduced the levels of CD 24, CD133 and EpCAM in CSCs, suggesting the inhibitory effect of BMSCs on stem cell-like capacities. Moreover, the inhibitory effect of BJJP on cell viability and the promoting effect of BJJP on cell apoptosis were also demonstrated. Based on the obtained results, we suppose that BJJP inhibit the tumorigenic potential of CSCs and would be beneficial for the treatment of HCC.
BMSCs are cells with muti-differentiate potentials, which can differentiate into osteoblasts, nerve cells, chondrocytes and adipocytes in different conditions. BMSCs play important role in the resistance of liver fibrosis and the protection of liver cells (29). It has been documented that multipotent adult progenitor cells (MAPCs) from BMSCs could further differentiate into cells with the functional capacity of hepatocytes in vitro (30). In vitro studies have found that MSCs derived from fetal skin inhibit the growth of human liver cancer cells, manifested by the reduced cell proliferation, colony formation and oncogene expressions (31). As we previously reported, the combination of human MSCs and Rougan Huaqian granules attenuate the liver fibrosis associated with liver cancer (32). In this study, BMSCs were extracted from BALB/c mice. The identification of BMSCs cell phenotype was firstly conducted, evidenced by the positive expressions of CD29, CD44, CD73, CD105 and CD166, and the negative expressions of CD14, CD34 and CD45, which is consistent with the previous studies (33). These results confirmed the successful isolation of BMSCs form BALB/c mice. Subsequently, we co-cultured BMSCs with CSCs, and observed the effects of BMSCs on the functions of CSCs. We discovered that BMSCs significantly suppressed the expressions of three markers of CSCs (CD24, CD133 and EpCAM). Furthermore, BMSCs significantly inhibited the cell viability of CSCs and promoted the apoptosis of CSCs. These results demonstrate that BMSCs affect the functions of CSCs and may be beneficial for HCC treatment. Collected evidence in this study highlighted that the combined use of BJJP and BMSCs achieved better therapeutic effect on HCC progression. Data in a previous study exploring the effect of BJJP drug serum on facilitating the transformation of hepatocyte to BMSCs, therefore we speculated that BJJP may able to enhance the suppressive effect of BMSCs on HCC cells by enhancing the transformation of hepatocyte to BMSCs (34). Aberrant expression of miR-140 has been reported in liver cancer and demonstrated as a liver tumor suppressor (12). Yang et al. (11) have found that miR-140 inhibits the proliferation, invasion and metastasis of HCC cells by down-regulating TGF-bR1 and FGF9. Lu et al. (13) have reported the promoting role of the lncRNAFLJ33360/miRNA-140/MMP regulatory loop in the metastasis of HCC. In our study, we discovered that BMSCs alone, BJJP alone or BMSCs combined with BJJP treatment of CSCs markedly promoted the expression of the anti-tumor gene miR-140 in CSCs. Importantly, knockdown of miR-140 in CSCs increased the levels of three markers of CSCs, cell viability and reduced cell apoptosis of CSCs administrated with BMSCs or BMSCs combined with BJJP. These results confirm that miR-140 is a tumor suppressor gene in HCC and is beneficial for inhibiting the tumorigenic potential of CSCs. Additionally, BJJP and BMSCs inhibit the tumorigenic potential of CSCs through regulating miR-140. A study in esophagus cancer showed that miR-140 could suppress ZEB2 to regulate Wnt/β-catenin signal pathway (35) and in melanoma, miR-140 was reported to target SOX4 so as to regulate Wnt/β-catenin signal pathway (36). Further exploration on downstream target of miR-14 in CSCs showed that miR-140 knockdown could increase the expressions of Wnt3a and β-catenin. But our study did not further explore the mechanism of miR-140 regulation on Wnt/β-catenin signal pathway. Western blot also showed that the combination use of BJJP with BMSCs could further suppress the expressions of Wnt/β-catenin signal pathway related proteins. Taken together, BJJP and BMSCs may be able to suppress HCC progression via upregulating miR-140 to suppress the activation of Wnt/β-catenin signal pathway.
In summary, we demonstrated that BJJP combined with BMSCs exert satisfactory regulation on the functions of CSCs through regulating the expression of anti-tumor gene miR-140 and Wnt/β-catenin signal pathway. There are several limitations of this study that should be addressed. Although our study highlighted the therapeutic effect of combination of BMSCs with BJJW on HCC, the explanation of how BJJP could have combination effect with BMSC was not mentioned in this study. Meanwhile, in vivo researches are required to investigate the potential mechanism of BJJP and BMSCs in HCC, which is the future direction for our study.
Go to :
 XML Download
XML Download