

 PDF
PDF Citation
Citation Print
Print


Introduction
Acute lung injury (ALI) is patho-physiologically characterized by alveolar epithelial cell destruction-caused increase in blood-air barrier permeability and non-cardiogenic pulmonary edema (1). Although improvements are seen in the pharmacological and supportive therapies for ALI, the disease still has a high mortality rate of more than 32% (2). Recent studies have reported that exogenous bone marrow-derived mesenchymal stem cells (MSCs) have a broad biodistribution in different organs including ALI lung tissues and can repair damage alveolar epithelium (3, 4). Because of their modulating effect on endogenous tissues and cells, MSCs with multipotent differentiation property have been applied in the treatment of multiple clinical disorders such as myocardial infarction, diabetes, and acute renal failure (5). Existing studies have shown that MSCs-based therapy is effective in alleviating human ALI (6). However, the low engraftment and differentiation rates of BMSCs limited their therapeutic effect in clinic (7). Therefore, elucidating the mechanism of promoting the alveolar cell-oriented differentiation may enhance the therapeutic effects of BMSCs implantation on ALI.
MicroRNAs (miRNAs) are a class of single-stranded, endogenous, and non-coding small RNAs that consist of 21-23 nucleotides (8). The function of miRNAs is to degrade mRNAs or inhibit mRNA translation by directly targeting their 3’-untranslated regions (3’-UTRs), which can further affect the proliferation, migration and differentiation of MSCs (8, 9), and regulate inflammatory responses (10). More importantly, previous studies have shown that miRNAs like miR-214, miR-16, and miR-23a are dysregulated in LPS (lipopolysaccharide)-induced ALI mouse model (11). MiR-23b-3p is an inflammation-related miRNA and was previously detected to be downregulated in LPS-induced male reproductive tract injury models (12). Moreover, miR-23b-3p upregulation facilitates the hepatic trans-differentiation of MSCs (13). However, the performance of miR-23b-3p in ALI and how miR-23b-3p mediates MSCs-based therapeutic effect on ALI remain obscure.
Fibroblast growth factor 2 (FGF2), which belongs to the family of transcriptional regulator fibroblast growth factors, plays crucial roles in regulating pathological conditions-mediated cell death and inflammatory responses (14). FGF2 can modulate the NF-κB pathway, which is a typical transcriptional factor that can orchestrate and sustain inflammatory responses by inducing the release of pro-inflammatory mediators and cause tissue damage (15). Cheng et al. (16) discovered that FGF2 regulates inflammation in LPS-exposed alveolar epithelial cells. These lines of evidence suggest that FGF2 inhibition may be a promising strategy to reduce ALI.
In this study, we established LPS-induced ALI models in mice to investigate the relation between miR-23b-3p and FGF2 and strove to propose a miR-23b-3p-related approach to improve the effect of MSCs-based therapy on ALI.
Go to : 
Materials and Methods
Ethics statement
All animal experiments were performed in Zhejiang Chinese Medical University in accordance with the guidelines of the China Council on Animal Care and Use. This study was approved by the Committee of Experimental Animals of Zhejiang Chinese Medical University (approval number: ICU201906026). Anesthetics and instant kill were made to minimize the pain and discomfort caused to the animals.
Cell cultures
Bone marrow-derived mesenchymal stem cells (BMSCs) from C57BL/6 mice were purchased from Cyagen Biosciences (MUBMX-01001, Santa Clara, CA, USA). The BMSCs were thawed and cultured in OriCell Mouse Mesenchymal Stem Cell Growth Medium (OCMM, MUXMX-90011, Cyagen Biosciences, USA) at 37℃ in 5% CO2. After infection with lentivirus carrying miR-27b-3p mimic or FGF2 overexpression plasmid, the BMSCs were passaged in OCMM at 37℃ in 5% CO2. Briefly, when the BMSCs grew to a monolayer of 80∼90% confluence, the media were aspirated. The BMSCs were rinsed with phosphate saline buffer (PBS, P5493, Sigma-Aldrich, St. Louis, MO, USA) for two or three times and then detached by 0.25% Trypsin-0.04% EDTA (T4049 Sigma-Aldrich, USA). The detached BMSCs were resuspended by OCMM and transferred to new flasks. The 6th or 7th passage of BMSCs was used for the following experiments except lentivirus infection.
293 T cells were purchased from Procell (CL-0005, Wuhan, China) and cultured in Dulbecco’s Modified Eagle Media (A4192101, ThermoFisher, Waltham, MA, USA) supplemented with 10% fetal bovine serum (FBS, F8687, Sigma-Aldrich, USA), 1% penicillin/streptomycin (V900929, Sigma-Aldrich, USA) and 1% Glutamine (G7513, Sigma-Aldrich, USA).
Lentiviral vector construction, packaging, and infection
Primers for miR-23b-3p overexpression (forward: 5’-CC TCGAGGTACCGTCGTATCCAGTGCAA-3’, reverse: 5’-GGGTACCCCGTATCCAGTGCGTGTCGT-3’), miR-23b-3p underexpression (forward: 5’-GGGACCGGGGTCCCTAATGGCAGCATAG-3’, reverse: 5’-GGAATTCCATAGGTCACGCACAGCACCT-3’), and FGF2 overexpression (for-ward: 5’-CCTCGAGGGCGACCCACACGTCAAACTA-3’, reverse: 5’-GGGTACCCTCCCTTGATAGACACAACTCCTC-3’) were used to produce the corresponding coding sequences. The coding sequence of miR-23b-3p overex-pression or FGF2 overexpression was cloned into pLVX-IRES-ZsGreen1 (Hedgehogbio, Shanghai, China) which had been digested by Xho1 and Kpn1 endonucleases (FD0694 and ER0521, ThermoFisher, USA). The coding sequence of miR-23b-3p underexpression was cloned into GV280 (Genechem, Shanghai, China) which had been digested by AgeI and EcoRI endonucleases (ER1461 and FD0274, ThermoFisher, USA). Empty lentiviral vectors were used as negative control. 293 T cells were seeded at 70% confluence in 12-well plates. Then, the 293 T cells were co-transfected either with the recombinant pLVX-IRES-ZsGreen1 vector, plp1, plp2 and plp/vsvg or with the recombinant GV280 vector, pGC-LV, Helper1.0 and Helper2.0 using Lipofectamine 3000 Transfection Reagent (L3000015, ThermoFisher, USA). Forty-eight hours after transfection, the culture media were collected. Viral particles were obtained via centrifugal ultrafiltration. Then, the viral particles were diluted and used to infect BMSCs.
Preparation of animals and establishment of ALI models
Fifty-four 6- to 8-week-old male C57BL/6 mice (20∼25g) were purchased from ALF Biotechnology (Jiangsu, China). All the mice were maintained at 20∼24℃ and 50% humidity on a 12 h/12 h light/dark cycle, and were given free access to food and water. Prior to experimen-tation, the mice were acclimated for 3 days.
All the mice were anesthetized via intraperitoneal injection of 2% pentobarbital sodium (P-010, Sigma-Aldrich, USA). For establishment of ALI models, the mice were intratracheally injected with a single dose of 100 μg lipopolysaccharide (LPS, L2630, Sigma-Aldrich, USA) which was previously dissolved in 50 μl normal saline (NS). The mice were allowed to recover until fully awake. The lentivirus-infected BMSCs (5×104 cells) were resuspended in 30 μl of PBS for implantation therapy.
Animal grouping
All the mice were randomly allocated into nine groups (n=6) and received different treatments as follows. Control group: mice received no further treatment; LPS group: 4 hours after modeling, ALI mice underwent intratracheal installation of 30 μl of PBS; LPS+NC group: ALI mice were intratracheally injected with BMSCs infected with lentivirus carrying negative control (5×104 cells in 30 μl of PBS; MOI=10); LPS+M group: ALI mice were intratracheally injected with BMSCs infected with lentivirus carrying miR-23b-3p mimic (5×104 cells in 30 μl of PBS); LPS+I group: ALI mice were intratracheally injected with BMSCs infected with lentivirus carrying miR-23b-3p inhibitor (5×104 cells in 30 μl of PBS); NC+FGF2-NC group: ALI mice were intratra-cheally injected with BMSCs infected with lentivirus carrying mimic control and FGF2-negative control; M+FGF2-NC group: ALI mice were intratracheally injected with BMSCs infected with lentivirus carrying miR-23b-3p mimic and FGF2-negative control; NC+FGF2 group: ALI mice were intratracheally injected with BMSCs infected with lentivirus carrying mimic control and FGF2 overexpression plasmid; M+FGF2 group: ALI mice were intratracheally injected with BMSCs infected with lentivirus carrying miR-23b-3p mimic and FGF2 overexpression plasmid. Lastly, all the mice were sacrificed via spinal dislocation, and their lung tissues were collected and stored at −80℃ until use in the following experiments.
Dual-luciferase reporter assay
The putative binding sites of miR-23b-3p on FGF2 transcripts were provided by Targetscan7.2 (http://www.targetscan.org/vert_72/), an online prediction tool. A reporter plasmid containing the 3’ UTR of FGF2-Wild-Type (5’-TGGTGTTATTTAGTTAATGTGAT-3’) or FGF2-Mutant-Type (5’-TGGTGTTATTTAGTTGGCTCACG-3’) was con-structed using pmirGLO vectors (E1330, Promega, Madison, WI, USA). BMSCs were seeded in 12-well plates (140656, ThermoFisher, USA) at 37℃ with 5% CO2. When reaching 70% confluence, the BMSCs were co-transfected with the reporter plasmid and either miR-23b-3p mimic (3’-CCAUUAGGGACCGUUACACUA-5’, miR10000891-1-5, RIBOBIO, China) or mimic control (3’-CACACACACA CAUGUGUGUGU-5’, miR10000125-1-5, RIBOBIO, China) using Lipofectamine 3000 Transfection Reagent. After transfection for 48 hours, the luciferase activity of the reporter plasmid was detected by a dual-Luciferase Reporter Assay System (E1980, Promega, USA).
Evaluation of lung edema
The ratio of lung wet weight to body weight (LWW/BW) was calculated and recorded to indicate the severity of lung edema. Briefly, the collected mouse lung tissues were trimmed for removal of extrapulmonary tissues, and then lung wet weight and body weight were measured. The results were presented as milligram per gram.
Quantitative reverse transcription polymerase chain reaction (qRT-PCR)
Total RNA and miRNA were extracted from lung tissues of ALI mice using Trizol reagent (15596026, ThermoFisher, USA) and PureLink miRNA Isolation Kits (K157001, ThermoFisher, USA), respectively. The total RNA and total miRNA were purified by successive treatments with chloroform (151858, Sigma-Aldrich, USA), isopropanol (I9516, Sigma-Aldrich, USA), and 75% ethanol (E7023, Sigma-Aldrich, USA). The purified total mRNA and miRNA underwent reverse transcription reaction with SuperScript IV reverse transcriptase (18090010, ThermoFisher, USA) for cDNA synthesis. A Real-Time PCR detection System (CFX Connect, Bio-Rad, Philadelphia, PA, USA) and PowerUp SYBR Green Master Mix (A25742, ThermoFisher, USA) were used for product amplification. The primers used for the amplification were listed in Table 1. The reaction conditions were: 95℃ pre-denaturation for 10 min, 95℃ denaturation for 15 s, and 40 circles of 95℃ annealing for 15 s and 60℃ elongation for 60 s. The relative expression levels were calculated by the 2−ΔΔCt method (17).
Table 1
Primers used in quantitative reverse transcription polymerase chain reaction for the target genes
![]()
Immunoblot
Total protein lysates were prepared from lung tissues of ALI mice using RIPA Buffer (89900, ThermoFisher, USA) and quantitated using a BCA protein assay kit (A53227, ThermoFisher, USA). Forty microliter of protein and 5 μl of marker (PR1910, Solarbio, China) were separately loaded on 12% SDS-PAGE gel (P0053A, Beyotime, Shanghai, China) and subjected to electrophoresis. Afterwards, the protein was transferred onto PVDF membranes (P2438, Sigma-Aldrich, USA), and the membranes were blocked by Tris Buffered Saline and 1% Tween 20 (TBST, TA-125-TT, ThermoFisher, USA) containing 5% skimmed milk. Then, the membranes were incubated at 4℃ overnight with primary antibodies against Occludin (SAB35 00301, 57 kDa, 1:1,000, Sigma-Aldrich, USA), SPC (ab2 11326, 21 kDa, 1:1,000, Abcam, Cambridge, MA, USA), FGF2 (SAB2108135, 31 kDa, 1:1,000, Sigma-Aldrich, USA), and GAPDH (ab181602, 36 kDa, 1:10,000, Abcam, USA). Then, the membranes were incubated with Goat Anti-Rabbit IgG (A32733, 1:20,000, ThermoFisher, USA) at room temperature for 1 hour. The immunoreactive bands were detected on an imaging system (iBright CL1500, A44240, ThermoFisher, USA) using an enhanced chemiluminescence reagent kit (WP20005, ThermoFisher, USA). The density of the bands was quantified by ImageJ software (version 1.52s, National Institutes of Health, Bethesda, MD, USA).
Histopathology
Lung tissues of ALI mice were fixed in 10% paraformaldehyde (16005, Sigma-Aldrich, USA) for 24 h, dehydrated by gradient alcohol, transparentized by xylene (95682, Sigma-Aldrich, USA), embedded in paraffin (1496904, Sigma-Aldrich, USA) and cut into 5-μm thick sections. Then, after being dewaxed by xylene and rehydrated by gradient alcohol, the sections were stained with Hematoxylin (H3136, Sigma-Aldrich, USA) for 12 min and eosin for 5 min. After staining, the sections were sealed with neutral balsam (N861409, Macklin, Shanghai, China) and dried at 37℃ for 4 hours. The histopathological changes of the lung tissues were observed by an optical microscope (13132071, fisherscientific, Massachusetts, USA) under 100× magnification.
Enzyme-linked immunosorbent assay (ELISA)
A tracheal cannula was passed through the nose into the lungs of ALI mice. Then, the tracheal cannula was withdrawn and flushed by pre-cooled PBS thrice. The flushing fluid was centrifuged at 1,000×g at 4℃ to obtain bronchoalveolar lavage fluid (BALF). The levels of interleukin (IL)-1β, IL-6, IL-4 and IL-8 in the BALF were measured by their corresponding ELISA kits (KGEMC001b, KGE MC004, KGEMC003, KGEMC104, keygentec, Jiangsu, China) according to the instruction of the manufacturer. Briefly, the BALF samples were diluted by normal saline, added into ELISA plates (100 μl/well) and incubated at 36℃ for 90 min. After being washed by PBS, the ELISA plates were added with biotinylated antibodies (100 μl/well) and incubated at 36℃ for 60 min. After being washed by PBS again, the ELISA plates were added with enzyme conjugation working solution (100 μl/well) and incubated at 36℃ for 60 min in the darkness. Lastly, Tetramethyl-benzidine was added into the ELISA plates for color development. The reaction was terminated using 2 mol/l H2SO4, and optical density was read under a microplate reader (ELx808, BioTek, Vermont, USA) at a wavelength of 450 nm.
Statistical analysis
All data were obtained from at least three independent experiments and are expressed as the mean±standard deviation. Statistical analyses were performed using SPSS 22.0 software (IBM, Armonk, NY, USA). Comparisons among multiple groups and between two groups were performed using one-way analysis of variance and student t-test respectively, and both were followed by Bonferroni’s post-hoc test. p<0.05 was considered statistically signifi-cant.
Go to :
Results
MiR-23b-3p was low-expressed in the lung tissues of ALI mice
To investigate the impact of implantation of BMSCs with manipulated miR-23b-3p expression on ALI, LPS-induced mice models of ALI were injected with BMSCs infected with lentivirus carrying miR-23b-3p mimic or inhibitor. qRT-PCR analysis showed that compared to the Control group, miR-23b-3p expression was decreased after LPS injection (p<0.001), while implantation of BMSCs overexpressing miR-23b-3p increased miR-23b-3p expression in ALI lung tissues, compared to the ALI mice without implantation (p<0.001, Fig. 1A). However, implantation of BMSCs under-expressing miR-23b-3p exerted no obvious effects on miR-23b-3p expression in ALI lung tissues.
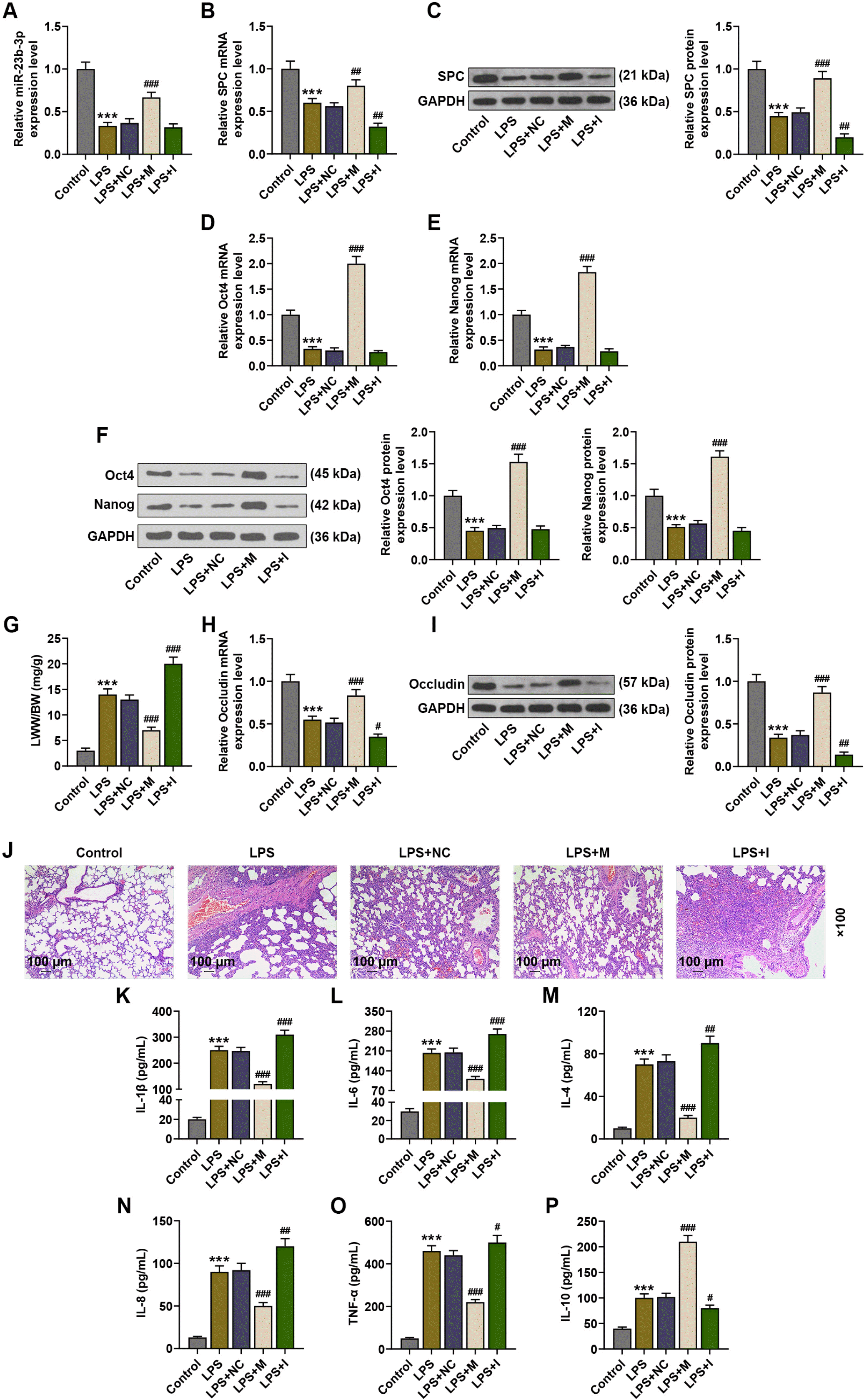 | Fig. 1MiR-23b-3p-overexpressing BMSCs promoted the differentiation of BMSCs into TypeII pneumocytes and alleviated injuries in ALI lung tissues. (A, B, D, E, H) The expressions of miR-23b-3p, SPC, Oct4, Nanog and Occludin in the lung tissues of LPS-induced ALI mice were analyzed by qRT-PCR, after injection of BMSCs with no altered gene expression or BMSCs with miR-27a-3p overexpression or underexpression. (C, F, I) The expressions of SPC, Oct4, Nanog and Occludin in the lung tissues of LPS-induced ALI mice were analyzed by immunoblot, with GAPDH serving as a reference gene, after injection of BMSCs with no altered gene expression or BMSCs with miR-27a-3p overexpression or underexpression. (G) The ratio of LWW to BW of LPS-induced ALI mice was calculated, after injection of BMSCs with no altered gene expression or BMSCs with miR-27a-3p overexpression or underexpression. (J) Histopathological changes in the lung tissues of LPS-induced ALI mice were observed via hematoxylin-eosin staining, after injection of BMSCs with no altered gene expression or BMSCs with miR-27a-3p overexpression or underexpression (scale: 100 μm; magnification: ×100). (K∼P) The levels of IL-1β, IL-6, IL-4, IL-8, TNF-α and IL-10 in the bronchoalveolar lavage fluid of LPS-induced ALI mice were assessed by ELISA, after injection of BMSCs with no altered gene expression or BMSCs with miR-27a-3p overexpression or underexpression. #p<0.05; ##p<0.01; ***p or ###p<0.001; * vs. Control ; # vs. LPS+NC (LPS: lipopolysaccharide, SPC: surfactant protein C, NC: negative control, M: miR-23b-3p mimic, I: miR-23b-3p inhibitor, qRT-PCR: Quantitative reverse transcription polymerase chain reaction, ELISA: Enzyme-linked immunosorbent assay, ALI: acute lung injury, LWW/BW: lung wet weight/body weight, BMSCs: bone marrow-derived mesenchymal stem cells).
|
MiR-23b-3p-overexpressing BMSCs promoted the differentiation of BMSCs, increased lung epithelium permeability, and improved lung edema
Subsequently, the expressions of SPC (a marker of TypeII pneumocytes), Oct4, Nanog (stemness marker) and Occludin (a marker of lung permeability) in ALI lung tissues were analyzed, and the LWW/BW ratio was calcu-lated. Occludin expression is associated with the status of the tight junctions of the lung epithelium (18). qRT-PCR and immunoblot analyses and lung edema evaluation revealed that LPS injection downregulated the mRNA and protein expression levels of SPC, Oct4, Nanog and Occludin and increased the LWW/BW ratio, compared to the Control group (p<0.001), while compared to the ALI mice without implantation, implantation of BMSCs overexpressing miR-23b-3p increased the expression levels of SPC, Oct4, Nanog and Occludin and decreased the LWW/BW ratio, and implantation of BMSCs under-expressing miR-23b-3p decreased the expression levels of SPC, Oct4, Nanog and Occludin and increased LWW/BW in ALI lung tissues (p<0.05, p<0.01, p<0.001, Fig. 1B∼I).
MiR-23b-3p-overexpressing BMSCs inhibited inflammatory factor release in ALI lung tissues
Later, changes in histopathology and inflammatory factor levels were detected in ALI lung tissues by Hema-toxylin-eosin staining and ELISA. As presented in Fig. 1J∼P, LPS injection caused inflammatory infiltration, structural damage, and increased levels of IL-1β, IL-6, IL-4, IL-8, TNF-α and IL-10, compared to the control group (p<0.001). Implantation of BMSCs overexpressing miR-23b-3p alleviated inflammatory infiltration and structural damage and lowered the levels of IL-1β, IL-6, IL-4, IL-8 and TNF-α, while increased the level of IL-10, compared to the ALI mice without implantation (p<0.001), while implantation of BMSCs under-expressing miR-23b-3p aggravated inflammatory infiltration and structural damage and elevated the levels of these inflammatory factors (Fig. 1J∼P).
MiR-23b-3p directly targeted FGF2
Bioinformatics analysis conducted on TargetScan V7.2 predicted that there existed highly conserved putative binding sites of miR-23b-3p on the 3’-UTR of FGF2-wild type (Fig. 2A). To validate the binding relation between miR-23b-3p and FGF2, we performed dual-luciferase reporter assay. The results showed that BMSCs co-transfected with the 3’UTR of FGF2-wild-type and miR-23b-3p mimic displayed a decreased luciferase activity, compared to those co-transfected with the 3’UTR of FGF2-wild-type and NC (p<0.001, Fig. 2B). However, transfection of miR-23b-3p mimic into the BMSCs loaded with FGF2-mutant type did not produce obvious changes in luciferase activity (Fig. 2B). These data suggested that miR-23b-3p could interact with FGF2 by directly binding to the 3’-UTR of FGF2.
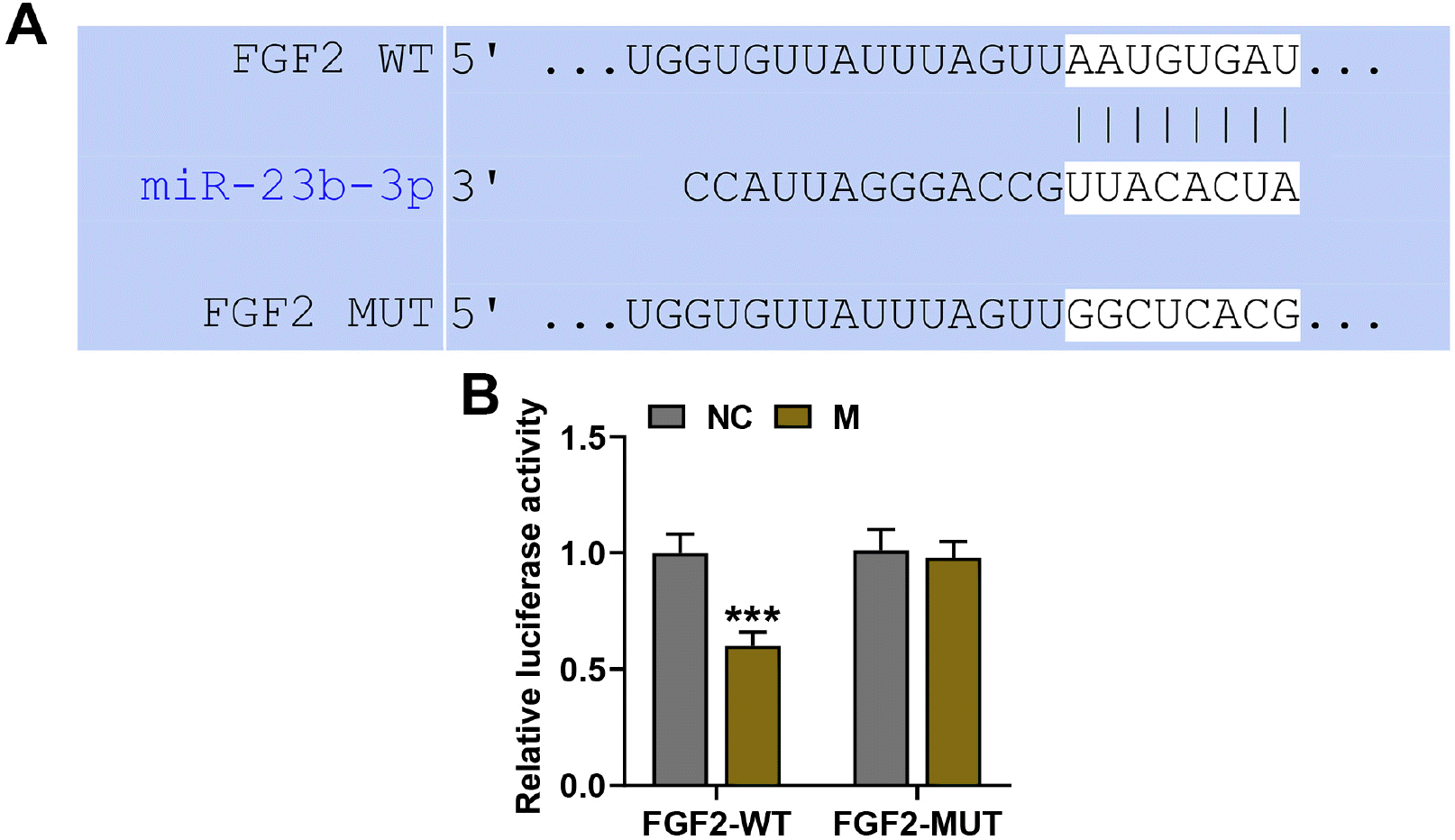 | Fig. 2MiR-23b-3p directly targeted FGF2. (A) The putative binding sites of miR-23b-3p on FGF2 were predicted by TargetScan V7.2. (B) The interaction between miR-23b-3p and FGF2 was validated by dual-luciferase reporter assay. ***p<0.001; * vs. NC (NC: negative control, M: miR-23b-3p mimic, WT: wild type, MUT: mutant type, FGF2: fibroblast growth factor 2).
|
FGF2 expression was inhibited by miR-23b-3p overexpressing BMSCs, and it inhibited the differentiation of BMSCs into TypeII pneumocytes
The role of the interaction between FGF2 and miR-23b-3p in ALI was examined via implantation of BMSCs infected with lentivirus carrying both miR-23b-3p mimic and FGF2 overexpression plasmid into ALI mice. As revealed by qRT-PCR and immunoblot analyses, miR-23b-3p-overexpressing BMSCs inhibited the mRNA and protein levels of FGF2, while implantation of BMSCs overexpressing FGF2 promoted these levels, compared to the NC+FGF2-NC group (p<0.001, Fig. 3A and 3B). Moreover, FGF2 mRNA and protein expressions under the implantation of BMSCs co-overexpressing miR-23b-3p and FGF2 were lower than those under the implantation of BMSCs overexpressing FGF2 alone (p<0.001), but higher than those under the implantation of BMSCs overexpressing miR-23b-3p alone (p<0.01, p<0.001, Fig. 3A and 3B). Furthermore, SPC mRNA and protein expressions were increased after implantation of BMSCs overexpressing miR-23b-3p, while they were decreased after implantation of BMSCs overexpressing FGF2, compared to the NC+FGF2-NC group (p<0.01, p<0.001, Fig. 3C and 3D). Moreover, SPC mRNA and protein expressions under the implantation of BMSCs co-overexpressing miR-23b-3p and FGF2 were higher than those under the implantation of BMSCs overexpressing FGF2 alone (p<0.001), but lower than those under the implan-tation of BMSCs overexpressing miR-23b-3p alone (p< 0.05, p<0.01, Fig. 3C and 3D).
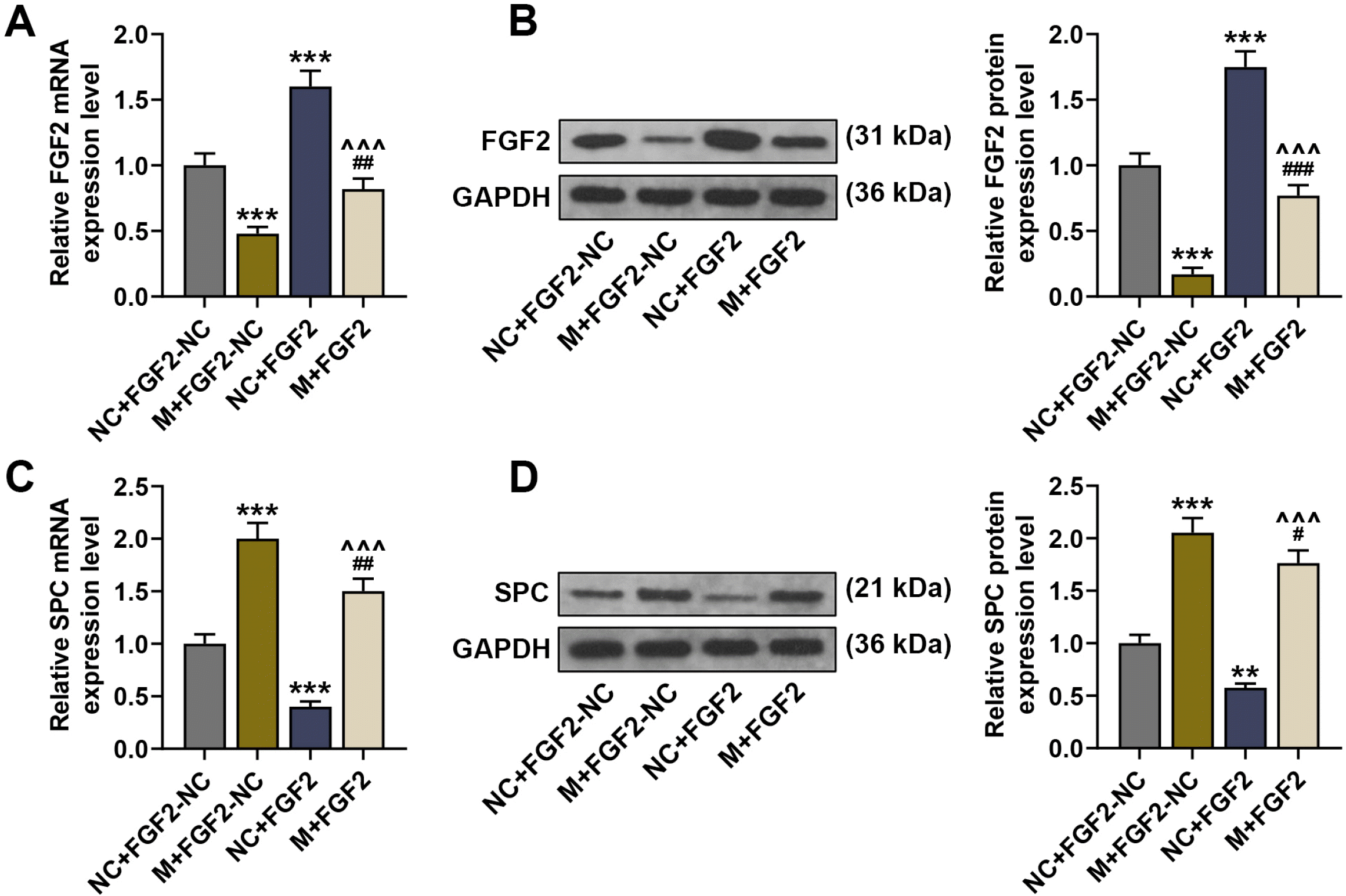 | Fig. 3FGF2 expression was inhibited by miR-23b-3p overexpressing BMSCs and it inhibited the differentiation of BMSCs into TypeII pneumocytes. (A, C) The expressions of FGF2 and SPC in the lung tissues of LPS-induced ALI mice were analyzed by qRT-PCR, after injection of BMSCs with no altered gene expression, BMSCs with overexpression of miR-27a-3p or FGF2 alone, or BMSCs with co-overexpression of miR-27a-3p and FGF2. (B, D) The expressions of FGF2 and SPC in the lung tissues of LPS-induced ALI mice were analyzed by immunoblot, with GAPDH serving as a reference gene, after injection of BMSCs with no altered gene expression, BMSCs with overexpression of miR-27a-3p or FGF2 alone, or BMSCs with co-overexpression of miR-27a-3p and FGF2. #p<0.05; **p or ##p<0.01; ***p or ###p or ^^^p<0.001; * vs. NC+FGF2-NC ; # vs. M+FGF2-NC; ^ vs. NC+FGF2 (LPS: lipopolysaccharide, SPC: surfactant protein C, FGF2: fibroblast growth factor 2, NC: negative control, M: miR-23b-3p mimic, qRT-PCR: Quantitative reverse transcription polymerase chain reaction, BMSCs: bone marrow-derived mesenchymal stem cells).
|
FGF2 reversed the effect of the implantation of BMSCs overexpressing miR-23b-3p on ALI lung tissues
Then, the relation between miR-23b-3p-mediated changes in FGF2 expression and ALI progression was further explored. As compared to the NC+FGF2-NC group, implantation of BMSCs overexpressing FGF2 alone increased the LWW/BW ratio and downregulated the mRNA and protein levels of Occludin (Fig. 4A∼C), and it reversed the effect of miR-23b-3p upregulation on lowering the LWW/BW ratio and promoting Occludin expression. Meanwhile, implantation of BMSCs co-overexpressing miR-23b-3p and FGF2 reversed the inhibitory effect of miR-23b-3p upregulation on the LWW/BW ratio and the inhibitory effect of FGF2 upregulation on Occludin expression (Fig. 4A∼C). Furthermore, implan-tation of BMSCs overexpressing FGF2 alone caused inflammatory infiltration and structural damage and elevated the levels of IL-1β, IL-6, IL-4, IL-8 and TNF-α, while decreased IL-10 level, compared to the NC+FGF2-NC group (p<0.05, p<0.01, p<0.001), and it reversed the inhibitory effect of miR-23b-3p upregulation on histopathological abnormalities and inflammatory factor release (Fig. 4D∼J). Meanwhile, implantation of BMSCs overexpressing miR-23b-3p reversed the promotive effect of FGF2 upregulation on histopathological abnormalities and inflammatory factor release (Fig. 4D∼J).
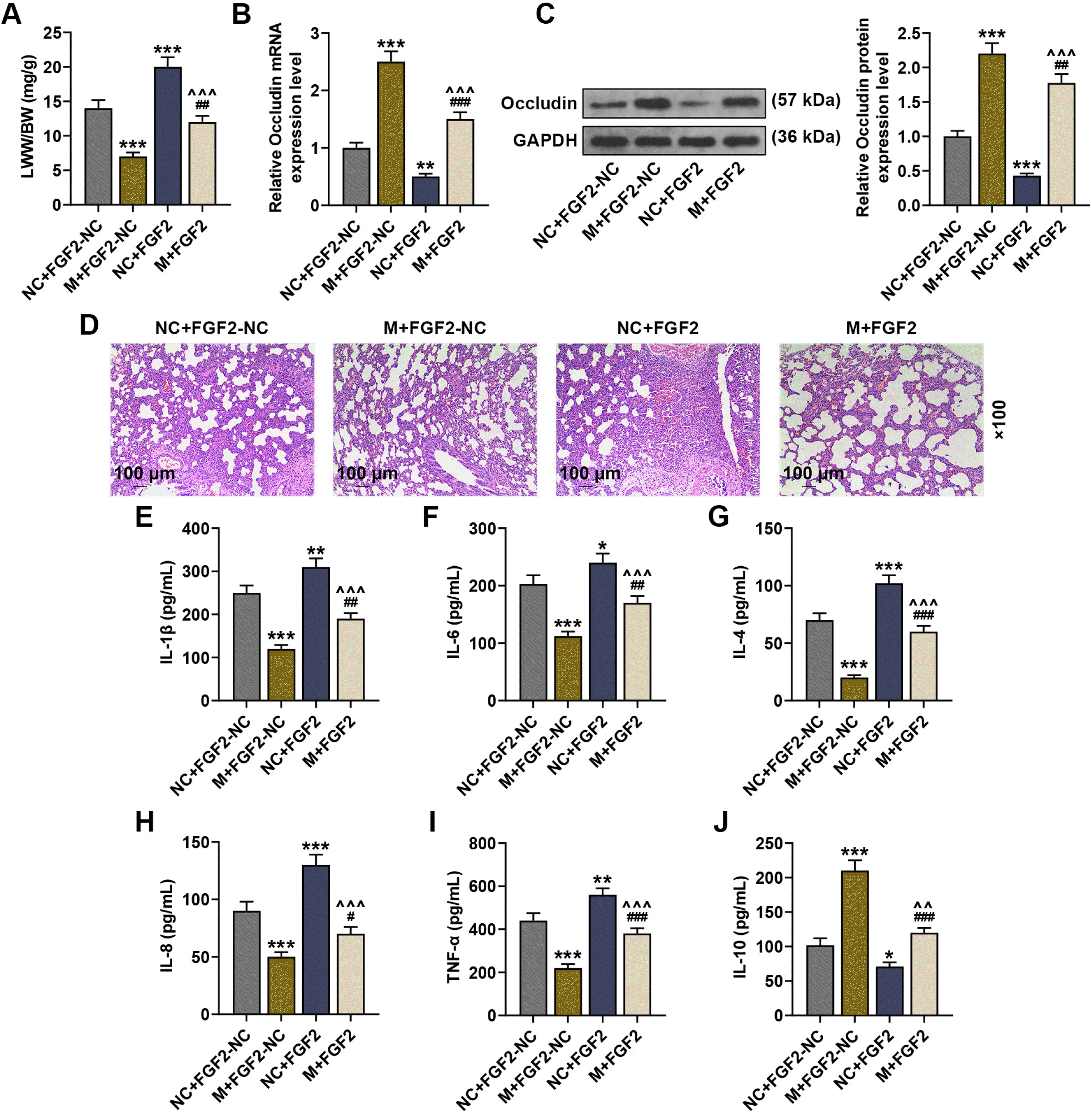 | Fig. 4FGF2 reversed the effect of the implantation of BMSCs overexpressing miR-23b-3p on ALI lung tissues. (A) The ratio of LWW to BW of LPS-induced ALI mice was calculated, after injection of BMSCs with no altered gene expression, BMSCs with overexpression of miR-27a-3p or FGF2 alone, or BMSCs with co-overexpression of miR-27a-3p and FGF2. (B) The expression of Occludin in the lung tissues was analyzed by qRT-PCR, after injection of BMSCs with no altered gene expression, BMSCs with overexpression of miR-27a-3p or FGF2 alone, or BMSCs with co-overexpression of miR-27a-3p and FGF2. (C) The expression of Occludin in the lung tissues of LPS-induced ALI mice was analyzed by immunoblot, with GAPDH serving as a reference gene, after injection of BMSCs with no altered gene expression, BMSCs with overexpression of miR-27a-3p or FGF2 alone, or BMSCs with co-overexpression of miR-27a-3p and FGF2. (D) Histopathological changes in the lung tissues of LPS-induced ALI mice were observed via hematoxylin-eosin staining, after injection of BMSCs with no altered gene expression, BMSCs with overexpression of miR-27a-3p or FGF2 alone, or BMSCs with co-overexpression of miR-27a-3p and FGF2 (scale: 100 μm; magnification: ×100). (E∼J) The levels of IL-1β, IL-6, IL-4, IL-8, TNF-α and IL-10 in the bronchoalveolar lavage fluid of LPS-induced ALI mice were assessed by ELISA, after injection of BMSCs with no altered gene expression, BMSCs with overexpression of miR-27a-3p or FGF2 alone, or BMSCs with co-overexpression of miR-27a-3p and FGF2. *p or #p<0.05; **p or ##p or ^^p<0.01; ***p or ###p or ^^^p<0.001; * vs. NC+FGF2-NC ; # vs. mimic+FGF2-NC; ^ vs. NC+FGF2 (LPS: lipopolysaccharide, FGF2: fibroblast growth factor 2, NC: negative control, M: miR-23b-3p mimic, qRT-PCR: Quantitative reverse transcription polymerase chain reaction, ELISA: Enzyme-linked immunosorbent assay, ALI: acute lung injury, LWW/BW: lung wet weight/body weight, BMSCs: bone marrow-derived mesenchymal stem cells).
|
Go to :
Discussion
BMSCs-based therapy is currently deemed as a feasible strategy for the treatment of ALI, during which BMSCs migrate to injured lungs to further differentiate into alveolar epithelial cells and facilitate the retention of the cells in the lung (3, 4). Recent studies have demonstrated that regulating gene expression in MSCs that are used in the implantation therapy for ALI can inhibit cell apoptosis, oxidative stress and inflammatory response and promote the migration of MSCs and their differentiation into TypeII pneumocytes in injured lung tissues, suggesting that MSCs may enhance BMSCs-mediated protection of lung tissues (19, 20). However, the low engraftment and differentiation rates limit the therapeutic potential of MSCs-based therapy (21). Therefore, finding a method to promote the migration and differentiation of MSCs is in urgent need. In this study, we revealed that miR-23b-3p mediated inhibition of FGF2 enhanced alveolar cell-oriented differentiation and inhibited the release of inflammatory factors, thus contributing to lung tissue repair in ALI.
MiRNAs are present in the circulatory system and changes in miRNA expression may herald the development of a variety of diseases including ALI (22, 23). Cardinal-Fernández et al. (24) have proposed that miRNAs are suitable biomarkers and therapeutic targets for ALI. As reported by Suo et al. (25), miR-1246 expression is significantly increased in ALI mice, whereas knockdown of miR-1246 inhibited cell apoptosis and inflammation in an in vitro ALI model. In contrast to our findings, Zhang et al. (26) discovered that miR-23b was downregulated in LPS-induced injury models and overexpressing miR-23b alleviated LPS-induced inflammatory responses. In addition, miR-23a-3p downregulation was previously found to be associated with LPS-induced rat testis (12), while miR-23a-3p upregulation was detected in acute laminitis horses (27). Our study demonstrated that LPS-induced ALI markedly downregulated miR-23a-3p expression, and similar to Dong’s findings (28), it also increased LWW/BW, promoted inflammatory infiltration and enhanced inflammatory factor release in vivo.
Meanwhile, Wang’s study demonstrated that the alleviation of LPS-induced ALI caused by lung-resident MSCs displayed an increased expression of SPC and a reduced number of inflammatory cytokines (29). SPC, an essential component of pulmonary surfactant, is primarily expressed in TypeII pneumocytes and has an anti-inflammation property, and SPC deficiency prompts the fast development of lung injury (30). Occludin plays a crucial role in assembling or maintaining epithelial tight junctions and regulating the barrier function of tight junctions (31). Deterioration of Occluding resulted in impaired function of alveolar epithelial barrier, which predisposes people to ALI (32). We also detected downregulated expressions of SPC and Occludin after LPS injection.
As emphasized above, lung tissue repair requires the retention of exogenous BMSCs in the damaged sites on tissues (33). We investigated BMSCs implantation-delivered therapeutic effect of miR-23a-3p on tissue damage and inflammatory responses in ALI. Our study discovered that injection of miR-23a-3p-overexpressing BMSCs into ALI mice not only restored miR-23a-3p expression, but also decreased the LWW/BW ratio, upregulated the expressions of SPC and Occludin, alleviated inflammatory infiltration, and repressed inflammatory factor release in the lung tissues. Previous study showed that miR-155-5p-mediated inhibition of differentiation of BMSCs into TypeII pneumocytes promoted the progression of acute respiratory distress syndrome (22). Raut and Khanna (13) reported that miR-23a-3p contributed to the promotion of hepatic trans-differentiation of MSCs. Therefore, our study inferred that upregulated miR-23a-3p in BMSCs might promoted the migration of BMSCs and their differentiation into TypeII pneumocytes to alleviate histopathological abnormalities and inflammatory responses, thus facilitating ALI repair.
MiRNAs can inhibit target mRNA translation or promote target mRNA degradation to modulate various biological functions (34). Our study showed that miR-23a-3p exerted an anti-inflammatory effect in ALI mice. Since FGF2 is a mediator of inflammatory responses (14), we selected FGF2 as a potential downstream mechanism of miR-23a-3p-mediated anti-inflammatory reaction. In our study, bioinformatic and experimental analyses confirmed that miR-23a-3p could directly target FGF2, and we found that BMSCs implantation-induced upregulation of miR-23a-3p decreased FGF2 expression in ALI mice, which implied that FGF2 might be involved in miR-23a-3p-mediated BMSCs-based lung tissue repair. FGF2 is widely implicated in various biological processes such as cell differentiation, proliferation, and angiogenesis (14). Besides, FGF2 participates in the regulation of autoimmune inflammation (35). Cheng’s study discovered that FGF2 overexpression significantly enhanced the release of IL-6 and IL-8 in LPS-stimulated alveolar epithelial cells by inducing the activation of the pro-inflammatory NF-κB pathway (16). Our study demonstrated that implantation of FGF2-overexpressing BMSCs downregulated the expressions of SPC and Occludin, while increasing the LWW/BW ratio and promoting the release of IL-1β, IL-6, IL-4 and IL-8, and it also suppressed the protective effect of miR-23a-3p on the lung tissues. However, the surrounding pericapillary repairs such as pericye activation, as progenitors of MSCs, angintensin II levels, or angiotensin converting enzymes 1-2 was need to further analyzed.
In conclusion, this study revealed that upregulation of miR-23a-3p in BMSCs used for implantation therapy alleviated lung edema and inflammatory infiltration and enhanced lung epithelial permeability through increasing the levels of SPC and Occludin and decreasing the level of inflammatory factors, and thereby promoted the repair of ALI lung tissues. Moreover, inhibiting FGF2 expression in BMSCs used for implantation therapy is the underlying mechanism of miR-23a-3p-mediated BMSCs implantation. However, whether miR-23a-3p directly promoted the differentiation of BMSCs into TypeII pneumocytes should be further validated.
Go to :
 XML Download
XML Download