

 PDF
PDF Citation
Citation Print
Print


Introduction
The NKX3.1 homeobox gene is located on human chromosome 8 and encodes a 234 amino acid protein whose functions are critical for embryogenesis as well as oncogenesis (1). Nkx3.1 is a putative prostate tumor suppressor and the earliest known marker of prostate formation during murine development, and its homolog in Drosophila is bagpipe (2). Nkx3.1 is expressed during all stages of prostate differentiation and is expressed largely in a prostrate- and androgen-specific manner, restricted to the luminal cells of the prostate gland (3). Loss of Nkx3.1 expression leads to defects in prostatic protein secretion and duct morphogenesis, and contributes to prostate carcino-genesis. Nkx3.1 expression is commonly lost in prostate carcinomas and prostatic intraepithelial neoplasia due to deletion of alleles, promoter methylation, or post-transcriptional silencing (1, 4).
The prostrate serves as the first line of defence against foreign pathogens originating from the male reproductive system. Prostatic inflammation acts as a marker for a clinical condition called prostatitis as well as prostate cancer (PCa) (5, 6). Prostatitis can be triggered by several factors including pathogens, chemical or mechanical trauma and diet (6). Prostatic inflammation is often accompanied by infiltration of specific immune cells into the prostrate, and their presence is associated with increased cancer risk and poor prognosis (7-9). Maintenance of the prostate and other male reproductive organs is the function of androgens such as testosterone and 5α-dihydrotestosterone (DHT) (10). The key effects of androgens, in addition to promoting normal physiological processes, include the initiation, development, and growth of prostate cancer (PCa) (11). Nkx3.1 plays a role in regulating transcription of the androgen receptor (AR) and thus favouring PCa cell survival (12). It is also active in non-prostate tissues and in regulating stemness. It has been identified in a rare population of luminal epithelial cells called CARNs (Castration-resistant Nkx3.1-expressing cells) that possess stem cell properties in prostate regeneration. It has also been shown to act as a transcription factor regulating Oct4 during reprogramming. Oct4 functions as a master regulator maintaining pluripotency and in the generation of induced pluripotent stem cells (iPSCs) (13). The substitution of the oncogenic Oct4 by a tumor suppressor has opened up multiple avenues of research that could be beneficial in the clinical application of these iPSCs.
This review therefore focuses on the role of Nkx3.1 as a transcription factor in various cellular functions including prostate development and cancers. The unconventional role of Nkx3.1 in regulating stemness and its novel function during the reprogramming of iPSCs is also considered in this review.
Go to : 
Structural Features of Nkx3.1
Nkx3.1 is a 234 amino acid protein that contains structural motifs mediating its interactions with other proteins and DNA. These motifs influence the function and stability of the Nkx3.1 protein. The three major regions of Nkx3.1 are the N-terminal region, corresponding to amino acids 1-123; the homeodomain region, corresponding to amino acids 124-183; and the C-terminal region, corresponding to amino acids 184-234 as depicted in Fig. 1A (14). Many of its amino acid are subject to post-translational modification and play unique roles in the functionality of Nkx3.1. Some of the features of this gene are discussed here.
 | Fig. 1Structural features of the homeobox protein Nkx3.1. (A) Nkx3.1 is a 234 amino acid protein that contains three major structural motifs crucial for its interactions with other proteins and DNA. Amino acids 1-123 correspond to the N terminal region and are depicted in purple. The homeodomain region corresponds to amino acids 124-183 and contains three helices, Helix I depicted in blue, Helix II in green and Helix III in yellow. The C terminal region corresponds to amino acids 184-234, depicted in brown. (B) Nkx3.1 protein structure. The structural feature of Nkx3.1 protein have been identified by nuclear magnetic resonance spectroscopy (22, 23). The N terminal is depicted in purple, Helix I in blue, Helix II in green, Helix III in yellow and the C terminal in brown.
|
Nkx3.1 has been reported to be regulated at the transcriptional level by several factors. Regulation by androgens is mediated by two androgen response elements (AREs) in the 3’ UTR region (15).
ETS1 and Sp1 have also been reported to regulate the expression of this gene by binding to the 5’ promoter (16, 17). Its transcriptional activity is also regulated by Wnt signalling through a positive regulatory loop that plays an important role in prostate bud growth and luminal epithelial differentiation (18). Selective CpG methylation at the Nkx3.1 promoter has been linked to diminished expression of Nkx3.1 in PCa, and evidence has been found suggesting an inverse correlation between MYC and Nkx3.1 levels during PCa progression (19, 20).
Divergent amino acid residues around the Nkx homeodomain contribute to the differential DNA binding specificity of members of the Nkx homeobox family, which characteristically contain a tyrosine residue at position 54 of the homeodomain. The Nkx3.1 homeodomain forms three α-helices. The first two generate the parallel scaffold, which stabilizes the third helix, perpendicular to them, which contacts the major groove of DNA (21). The structural features of the Nkx3.1 homeobox protein are depicted in Fig. 1 (22, 23). Nkx3.1 has been shown to have a binding preference for the consensus sequence 5’-TAAGTA-3’ over 5’-TAAGTG-3’ (24).
Go to :
Post-Translational Regulation of Nkx3.1
Nkx3.1 is regulated at the gene level as well as at post-translational levels. The stability of the NKX3.1 gene product is influenced by modification of the homeodomain and flanking regions. Mutations of threonine to alanine at positions 89 and 93 in the N-terminal flanking region reduce its half-life by 50% (25). Protein kinase CK2 is responsible for phosphorylating these threonine residues and prolonging half-life (25). In contrast, at the C-terminal, phosphorylation of serine 185 controls the ubiquitination and degradation of the protein, and mutation of serine 185 to alanine prolongs half-life (26). Inflammatory cytokines such as TNF-α induce phosphorylation of serines 195 and 196 (27). Nkx3.1 expression is reduced in inflammatory cell-containing tissues and this effect appears to be related to the accelerated ubiquitination and degradation of Nkx3.1 due to TNF-α and IL-1β observed in vitro (26, 27). Serine-48-mediated phosphorylation is also critical in Nkx3.1 and has been shown to be affected by the status of the arginine at residue 52, which is a common target for genetic polymorphism. C145T polymorphism causes a non-conservative R52C amino acid replacement that decreases phosphorylation at serine 48 and impairs DNA binding in vitro (28).
Nkx3.1 also undergoes ubiquitination by TOPORS, a RING-finger E3 ligase (29). Topoisomerase I and TOPORS are together responsible for the DNA-unwinding activity of Nkx3; both are expressed in the prostate, making TOPORS a highly efficient E3 ubiquitin ligase for Nkx3.1 (29-31). In vitro and in vivo experiments have revealed TOPORS-mediated ubiquitination of Nkx3.1. Overex-pression of TOPORS enhanced proteasomal degradation of Nkx3.1 in PCa cells, whereas its knockdown increased Nkx3.1 half-life and steady state level (29). TOPORS-mediated ubiquitination is largely targeted via the Nkx3.1 homeodomain, which contains nine lysine residues that can potentially be ubiquitinated, and the N-terminal domain also has some activity (29). The exact site responsible for TOPORS-mediated ubiquitination is yet to be established. The homeodomain Lys182 residue plays an important role in Nkx3.1 turnover via PIM1 inhibition; however, the role of TOPORS in this process has not been elucidated (32).
Go to :
Functions of Nkx3.1
Although the role of Nkx3.1 dysregulation in PCs has been well documented over the years, much remains to be understood about the functions of this transcription factor.
Nkx3.1 in prostate development and cancer
The prostate is a male accessory sex gland that supports and promotes male fertility and insemination. It is composed of small exocrine glands that produce a thin, slightly alkaline fluid that is rich in proteins and compounds like citric acid, acid phosphatases, cholesterol, zinc, calcium, and prostate specific antigen (PSA) (33). The human prostate gland is located below the urinary bladder surrounding the prostatic urethra, residing in front of the rectum. It is covered by three separate, distinct fascial layers-prostatic, endopelvic, and denovilliers-and a band of fibromuscular stroma (34, 35). The unilobular prostate has been classified into four globular zones, designated transitional, peripheral, fibromuscular, and central (36, 37). In contrast, the murine prostate is composed of multiple lobes and arranged at the base of the bladder around the urethra. Based on their anatomical positions, these lobes have been classified as anterior prostate (AP), ventral prostate (VP), dorsal prostate (DP), and lateral prostate (LP), each having a unique secretion pattern, morphology, and histology (38). Even though the murine prostate lobular structure and human prostate zonal architecture share no overall homology, the dorsolateral and anterior lobes in the mouse are similar to the peripheral zone of the human prostate (39).
Formation of the prostate in mice as well as in humans occurs through epithelial budding from the urogenital sinus (UGS), which is a derivative of the hindgut endoderm arising mid-gestation during mammalian development. The UGS also develops into the bulbourethral glands (BUGs) and prostatic urethra (40). The significance of Nkx3.1 in prostate development is highlighted by developmental defects in mutant mice leading to downregu-lation of genes essential for prostate differentiation, reduced ductal branching, decreased secretory protein production, and epithelial hyperplasia and dysplasia in the prostate, accompanied by changes in cellular differen-tiation and decreased secretory proteins in BUGs (41-43). In mice, expression of Nkx3.1 RNA is first observed at approximately 15.5 days post-coitum (dpc) in the lateral urogenital sinus epithelium (UGE), which precedes formation of the prostate and BUG (41). At the end of gestation, approximately 17.5 dpc, prostatic buds emerge at the rostral end of the UGE, and Nkx3.1 expression has been localized to the leading edge of outgrowing prostatic ducts (41). Roles of Wnt and Fgf10 signalling in activating Nkx3.1 expression during prostate organogenesis have been demonstrated by studies using urogenital sinus explant cultures (44-46). Expression of Wnt ligands has been detected in the urogenital mesenchyme and epithelium prior to and during prostate development (47-49). Deletion of β-catenin resulted in loss or reduction of prostate bud formation, further suggesting a role of Wnt signalling in prostate formation (46, 50). β-Catenin is required during embryonic prostate growth and branching but is dispensable in normal adult organs. β-Catenin regulates progenitors in the epithelial buds as well as a discrete network of genes that include c-Myc and Nkx3.1 during development (46).
Nkx3.1 expression is restricted to the luminal epithelium of mature prostate post-ductal canalization and to the UGE caudal ends where the epithelial buds of BUGs develop (41). Expression of Nkx3.1 continues through BUG morphogenesis to the appearance of the mature epithelium (41). Although the initial expression of Nkx3.1 precedes that of androgen receptors (ARs) in the prostatic epithelium, subsequent expression appears to be dependent on androgen signalling (41). Although Nkx3.1 is positively regulated by androgens, it inhibits the expression of the AR, forming an important loop regulating PI3K-AKT signalling pathway activation via an AR/PI3K-dependent mechanism (51). Castration-mediated androgen deprivation in mice leads to regression of the prostate and apoptosis of the majority of its luminal cells, as well as loss of Nkx3.1 in the cells that survive (3, 52, 53). Nkx3.1 null mutant mice display defects in epithelial differentiation, as indicated by altered prostate secretory protein expression, epithelial hyperplasia, and expression of seminal vesicle markers, underlining the importance of this gene during prostate epithelial specification (41, 43).
Nkx3.1 is recognized as a tumor suppressor because its expression decreases with progression of PCa, an effect corroborated by immunohistological analysis of human neoplastic prostate epithelium (54). PCa, like other epithelial cancers, arises from precursor lesions called prostatic intraepithelial neoplasia (PIN), which are induced by inflammatory mitogenic factors and ultimately develop into locally invasive disease and metastasis (55, 56). PCa initiation has been attributed to a reciprocal relationship between inflammation and the status of the NKX3.1 gene (57). Studies of Nkx3.1 mutant mice have shown that cancer initiation correlates with enrichment of specific immune populations and increased immune-regulatory gene expression together with the finding that human prostate tumors show low Nkx3.1 expression (57). Loss of Nkx3.1 expression reportedly leads to aberrant overexpression of a cohort of cancer-associated genes that include clusterin and quiescin Q6 (58-60). Nkx3.1 further exacerbates inflammation-induced initiation of PCa, as evidenced by enhanced epithelial plasticity and defects in cellular differentiation (57). A gradual decrease of Nkx3.1 expression has been reported in benign prostatic hyperplasia, high-grade prostatic intraepithelial neoplasia (HGPIN), metastases, carcinomas, and pre-invasive and invasive cancer cells (19, 54). Although the exact mechanisms underlying complete loss of Nkx3.1 have not been elucidated, there are clearly selective pressures leading to its loss during PCa development.
Numerous genetic, epigenetic, and chromosomal mutations have been linked to specific stages of PCa progression (61). Human NKX3.1 maps to chromosome 8p21 and is reported to frequently undergo loss of heterozygosity (LOH) in cases of PIN and PCa (62). About 21 germ-line mutations, which correlate with hereditary disease, have been described at this locus while somatic mutations have not been reported. One germ-line mutation is the T164A polymorphism in the NKX3.1 homeodomain, which reduces Nkx3.1 binding to its cognate DNA binding sequence (22). Genome-wide association studies (GWAS) have helped identify single nucleotide polymorphisms in the Nkx3.1 sequence that are markers for PCa suscepti-bility. For example, a single nucleotide polymorphism in one allele of the 5’UTR led to decreased binding affinity of Nkx3.1 for transcription factor Sp1 (63). In this variant, target gene expression was altered and there was decreased transcriptional activity at the Nkx3.1 promoter, reducing its expression (63).
Expression of the tumor suppressor PTEN (phosphatase and tensin homolog deleted on chromosome 10) is critical for maintenance of Nkx3.1 expression in the prostate and is frequently altered in human cancers. PTEN mutations have been strongly implicated in PCa development and have been seen in primary PCa as well as metastatic prostate tissues (64, 65). Murine gene targeting experiments have revealed an interaction between Nkx3.1 and PTEN that supresses PCa, and reductions in the levels of the two factors act synergistically to augment prostate neoplasia (66, 67). PTEN is a phosphatase for Nkx3.1, which acts to maintain the level of Nkx3.1 under conditions that induce the degradation of Nkx3.1 via its phosphorylation (68). Nkx3.1 also plays an important role in modulating the half-life of p53 by regulating the activity of nuclear MDM2 and HDAC1. Nkx3.1 binds to HDAC1, thereby stimulating the activity and acetylation of p53 by releasing it from p53-MDM2-HDAC1 complexes. The formation of these complexes would otherwise result in p53 degra-dation. Reduced p53 activity leads to activation of the PI3K/AKT pathway and ultimately stimulates cell proliferation, decreasing cell death and the initiation of PCa (67).
Nkx3.1 functions in non-prostate tissues
Nkx3.1 is transiently expressed during organogenesis across a wide range of tissues outside the urogenital system. Expression is most evident in the paraxial mesoderm, where it is localised to the ventral regions of the most caudal eight-to-nine somites and becomes restricted to the sclerotome (69, 70). Expression of Nkx3.1 in explant cultures of somites is dependent on the neural tube or notochord and is induced by Sonic Hedgehog (69). Despite the expression of Nkx3.1 in the newly-formed paraxial mesoderm, Nkx3.1 null mutants do not display any phenotype in sclerotomal derivatives (70, 71). Nkx3.1 expression has also been reported in the distal epithelium of the tongue, dorsal aorta, a subset of teeth, the arcuate and interlobular arteries of the kidney, and the dorsal region of Rathke’s pouch (71, 72).
Nkx3.1 null mutants display only a minor phenotype in the lobular arteries of the kidneys, seen as vacuolation of the arterial walls. No other phenotypes have been observed for other non-urogenital tissues that express Nkx3.1, including the cerebral cortex, pituitary glands, hippocampus, and hair follicles (42). The only significant non-urogenital phenotype described for Nkx3.1 null mutants occurs within the minor salivary glands of the oral cavity, where there is increased accumulation of mucous secretions accompanied by a significant decrease in ductal branching of the palatine glands (42, 71). The prognosis for oral squamous cell carcinoma (OSCC) depends significantly on the presence of cervical lymph node metastasis (LNM), and genome-wide analysis of DNA copy number aberrations (CAN) using data from OSCC patients has shown that expression of NKX3.1, along with several other genes, is significantly correlated with LNM (73). The role of Nkx3.1 in OSCC has been further validated by qRT-PCR and immunohistochemical (IHC) analyses showing reduced Nkx3.1 expression in occult LNM cases (73). Nkx3.1 expression level is a useful aid in diagnosing salivary duct carcinoma (SDC). However, SDC progression was not correlated with decreased Nkx3.1 expression (74).
Nkx3.1 is an important mediator of bladder fibrosis in bladder outlet obstruction (75). Fibrosis leads to stiffening of healthy tissues and organ dysfunction due to the irreversible remodelling caused by deposition of collagen in the extracellular matrix of various organs. Expression of Nkx3.1 is induced in the bladder following partial bladder outlet obstruction (PO), and is responsible for activation of pathways leading to tissue fibrosis (75). Even though the bladder appears to be functionally and morphologically intact, fibrosis is attenuated in the absence of Nkx3.1 (75). Nkx3.1 is also a potential therapeutic target for the treatment of nasopharyngeal carcinoma (NPC), along with cyclin B2 (CCNB2) (76). NPC tumors arise from the epithelial cells covering the surface and lining of the nasopharynx, and gene expression profiling of NPC identified NKX3.1 as a down-regulated and CCNB2 as an up-regulated gene; in agreement with this, overexpression of Nkx3.1 led to loss of NPC cell viability, and invasiveness (76).
Nkx3.1 and stemness
The activities and regulation of adult epithelial stem cells can be studied in the prostate system, which provides contexts for examining changes due to cancer development and tissue regeneration. Three differentiated cell types exist in prostate epithelium: basal cells, luminal secretory cells, and neuroendocrine cells (77). Absence of androgens results in rapid apoptosis of approximately 90% of luminal cells. However, a small, number of basal cells are stably maintained in the regressed state (78). Re-administration of androgens results in regeneration of the prostate epithelium, implying the presence of a population of stem cells (79, 80). A role of Nkx3.1 as a stemness-re-gulating factor was first suggested by Wang et al. (53), who by lineage-tracing identified a population of rare luminal cells that expressed Nkx3.1 even in the absence of testicular androgens. These bipotential, self-renewing cells, which could regenerate prostate ducts in renal grafts, were called castration-resistant Nkx3.1-expressing cells or CARNs (53). When CARNs were marked in genetically-engineered mouse lines using an inducible CreERT2 recombinase under the control of the endogenous Nkx3.1 promoter. Cre activation in castrated Nkx3.1CreERT2/+; R26R-YFP/+ male mice by tamoxifen led to yellow fluorescent protein (YFP) expression in luminal epithelial cells that corresponded to lineage-marked CARNs. All luminal cells, as well as a small percentage of basal cells, normally express Nkx3.1 in the hormonally-intact adult prostate, whereas only the rare luminal CARNs express Nkx3.1 in regressed epithelium (81). The multipotent nature of CARNs was also demonstrated by transplantation of single lineage-marked CARNs into the kidney, which resulted in the formation of prostatic ducts containing all three prostate epithelial cell types (53).
Two possible functions have been proposed for the CARNs in prostate epithelium (81). One model assumes the co-existence of basal as well as luminal stem cells within the prostate epithelium, with CARNs representing the luminal stem cells. According to the other model, CARNs are facultative stem cells that correspond to luminal progenitors, and acquire stem cell properties after androgen deprivation (81). These models imply different roles for CARNs as ‘cells of origin’ in PCa. ‘Cell of origin’ models posit the initiation of tumorigenesis from a normal tissue cell that retains the ability to self-renew and generates different cell lineages within tumors. CARNs maintained stem-like properties and expressed markers such as Sox2, Nanog, Oct4 and ALDH1A1 associated with stemness in an established human xenograft model (82). Besides Nkx3.1-expressing CARNs, other forms of cancer stem cell (CSC) have been identified that give rise to castration-resistant prostate cancers (CRPC). One such example includes cancer-associated fibroblasts (CAF) (83-85).
Another role of Nkx3.1 has been recently elucidated, namely as a replacement for Oct4 in stem cell reprogramming (86). Originally, Takahashi and Yamanaka (87) identified four factors capable of reprogramming differentiated cells back to stem cells -Oct4, Sox2, Klf4, and Myc (OSKM), collectively known as Yamanaka factors. Viral-mediated transduction of the Yamanaka factors has provided a major technique for generating induced pluripotent stem cells (iPSCs)-since that time (88). However, activation of an endogenous pluripotency program can create a barrier to reprogramming (89), and overexp-ression of oncogenic transcription factors during somatic reprogramming poses a risk of malignant transformation, which limits their use in a clinical setting (90). Various groups have attempted to replace or omit some of the reprogramming factors in order to understand the molecular role of each factor (91-94). Others have attempted to use small molecule compounds as substitutes (91, 95, 96). One of these strategies, replacement or omission of Oct4, has gained a lot of attention (86). As an alternative to omitting oncogenes during reprogramming, the use of tumor suppressor genes such as p53, p16 and Rb1 as regulators of cell reprogramming is well established (90, 97-99). The process of reprogramming has usually been associated with deletion of tumor suppressor genes and duplication of oncogenes (100), and the tumor suppressor p53 has been reported to act as a barrier to reprogramming. However, iPSCs generated by incorporating an extra copy of the p53 locus were found to retain normal pluripotency and to have reduced tumorigenic potential in mice (90, 97). Thus, increasing the dosage of tumor suppressors to generate iPSCs may limit their tumorigenicity and increase their clinical applicability (90).
A previously unknown role of the tumor suppressor Nkx3.1 was revealed by using a heterokaryon reprogramming system generated by fusing human fibroblasts and mouse embryonic stem cells (ESCs) (86). This system was used to generate molecular maps of early reprogramming by RNA-seq at time points over the first 24 hours post fusion. Techniques such as assays for transposase-acces-sible chromatin by sequencing (ATAC-sequencing) and motif-enrichment analysis then demonstrated a brief period of expression of Nkx3.1 two to three hours post fusion, at the onset of reprogramming (86). Oct4 and Nanog were reported to be the first to show signs of transcriptional activation one day post-fusion (101). Since Nkx3.1 expression preceded activation of Oct4 the authors proposed that Nkx3.1 was an upstream activator of endogenous Oct4, and this was confirmed by demonstrating presence of Nkx3.1 at the Oct4 promoter. Further experiments omitting and replacing individual components from the reprogramming cocktail revealed the potential of Nkx3.1 as a replacement for exogenous Oct4 in human and mouse reprogramming (86).
Reprogramming of fibroblasts using OSKM has revealed that Nkx3.1 acts downstream of the IL6-Stat3 signalling cascade. Expression of Nkx3.1 depends upon IL-6R, which is essential for OSKM reprogramming in both human and mouse fibroblasts. Stat3 has been reported to play a role in activating Nkx3.1 as well as Oct4; as shown in Fig. 2A, co-expression of Stat3 and Nkx3.1 rescued colony formation in the absence of Il-6r. When Nkx3.1 was used to replace Oct4 in the Oct4, Sox2, Klf4 (OSK) cocktail the results showed that reprogramming of Nkx3.1, Sox2, Klf4 (NSK)-derived iPSCs were as efficient as that of OSK-derived iPSCs in mice and humans (Fig. 2B). The NSK-derived iPSCs were pluripotent and capable of producing cells of all three germ layers (86).
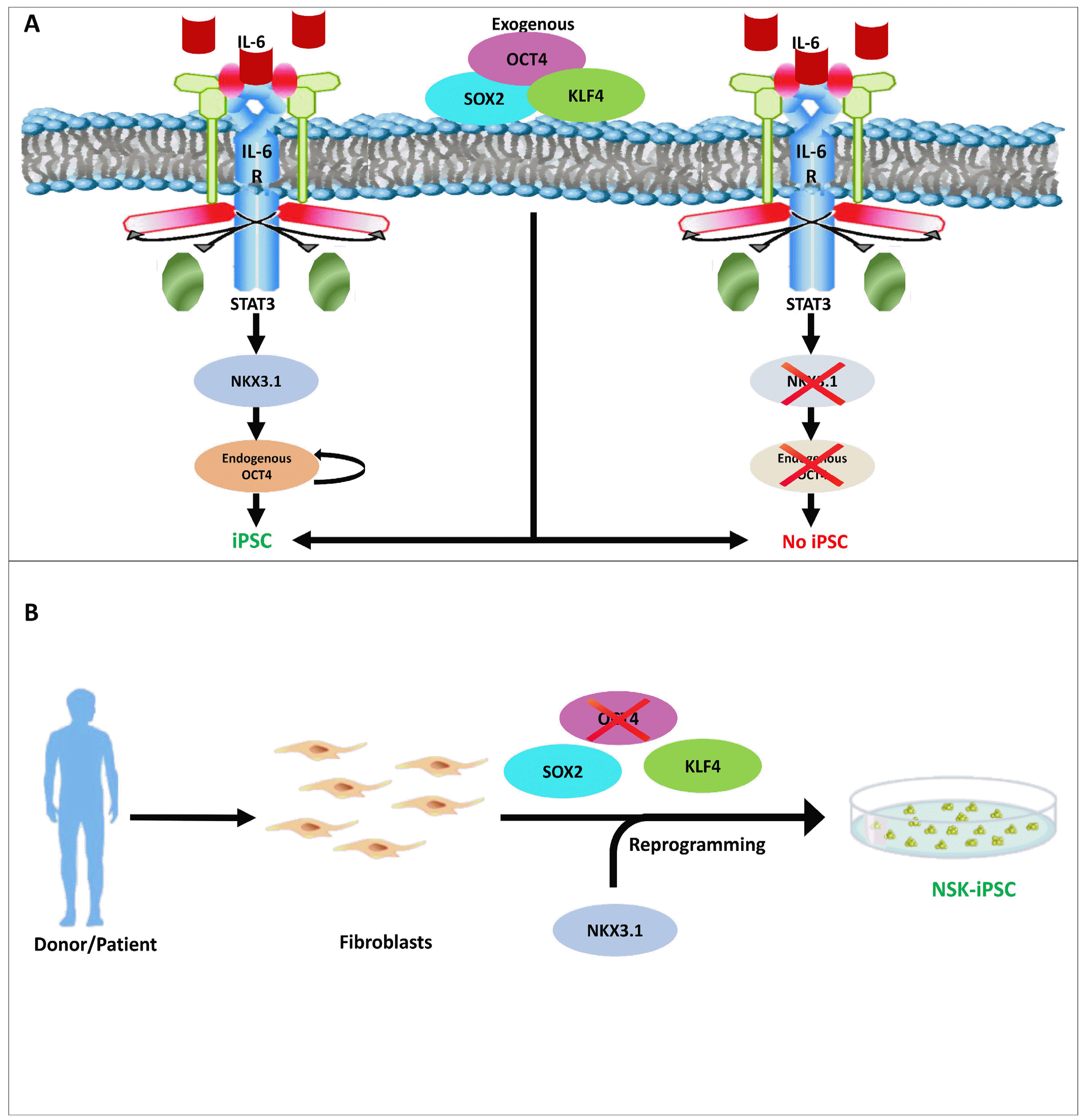 | Fig. 2The activities of Nkx3.1. (A) The role of Nkx3.1 in the IL-6-STAT3 signalling pathway during iPSC in-duction. Nkx3.1 is a target of STAT3, downstream of IL6r activation and is essential during reprogramming. IL-6r deficiency during reprogramming is reported to be rescued by Nkx3.1 alone or in cooperation with STAT3. However, knockdown of STAT3 decreases Nkx3.1 protein level, whereas knockdown of Nkx3.1 blocks iPSC reprogramming (86). (B) Nkx3.1 can replace Oct4 in cellular reprogramming, and promotes the generation of iPSCs with a similar efficiency to Oct4.
|
Go to :
Perspectives
The androgen-regulated, prostate specific-tumor suppressor, Nkx3.1, plays major roles in controlling epithelial cell growth, cell differentiation, and stem cell maintenance (41, 53, 102). However, as reviewed in this article, loss of expression of this gene has been widely implicated in the development of PCa and thus serves as a marker for cancer progression (54, 68). The clinical relevance of Nkx3.1 has so far been limited to its use as a marker for PCa progression and it has not been subjected to targeted therapeutic approaches (54). Up-regulation of Nkx3.1 in vitro in prostate cancer cell lines has revealed some promising anti-cancerous effects, but these have not been transferred to a clinical setting. Up-regulation of Nkx3.1 using synthetic double stranded ODNs in LNCaP inhibited cell proliferation, while stable expression of Nkx3.1 inhibited the proliferative and invasive activities of PC-3 (103, 104). Nkx3.1 has been reported to be stabilized by inhibition of dual-specificity tyrosine-regulated kinase 1B (DYRK1B) which regulates Nkx3.1 via phosphorylation at serine 185, and inhibition of this kinase in prostate cancer increased the level of Nkx3.1 thereby proving the kinase to be a potential therapeutic target in prostate cancer (105). Nkx3.1 has also been reported to be regulated via other posttranslational modifications, as reviewed above, and these regulators provide alternative potential anti-cancer therapeutic approaches that require further study.
Besides the huge potential of Nkx3.1 as a therapeutic target, it has also been shown to play a role in CARNs and during iPSC differentiation. The oncogene Oct4 serves as a master regulator in maintaining pluripotency and has been frequently used for generating iPSCs (106-109); iPSC generation is indeed dependent on the direct activation of endogenous Oct4 (110-112). However, the recent identification of Nkx3.1 as a potential replacement for Oct4 paves the way for future research focused on Nkx3.1 as a regulator of pluripotency and stem cell differentiation. All these findings suggest that Nkx3.1 may be a transcription factor responsible for the expression of other as yet undefined genes that are activated during repro-gramming. Cellular DNA damage repair pathways have also been reported to be activated by Nkx3.1 and could potentially help maintain the genomic integrity of NSK-derived iPSCs, which promises to be an interesting avenue of investigation (113). Like the tumor suppressor role of Nkx3.1, numerous other genes may have previously overlooked roles that could be identified using similar approaches. Because Nkx3.1 is itself a tumor suppressor gene, its ability to replace Oct4 in iPSC induction has broad implications, especially in the clinical applications of iPSCs.
Go to :
Conclusions
Nkx3.1 is well known as a prostate-specific tumor suppressor gene; however, its role has now been expanded to non-prostate tissues. Nkx3.1 is regulated by androgens in normal prostate cells, but this regulation is consistently lost in prostate tumors, and reportedly triggers a cohort of genes that are overexpressed during tumor initiation. Nkx3.1 also plays an important role in early mouse development and is expressed in the mouse embryo as early as day 15.5 dpc. A previously unrecognized role of this prostate-specific gene is in self-renewing prostate luminal cells called CARNs, whose effect points to the possibility that Nkx3.1 is a stemness gene. Further investigation has revealed a role of Nkx3.1 in reprogramming differentiated cells back into iPSCs, thus opening up the prospect that it can activate Oct4 and promote pluripotency.
Go to :
 XML Download
XML Download