

 PDF
PDF Citation
Citation Print
Print


Introduction
A stable humanized mice (hu-mice) model is imperative for understanding information regarding the immune response in a specific microenvironment (1). Hu-mice model is a valuable platform for investigating the effects of cell therapies and drug screening in biomedical research (2). The hu-mice model is reconstructed with human CD34+ hematopoietic stem cells (HSCs) using immune-compromised mice such as NOD/LtSz-scid IL2Rγnull (termed NSG, generated from Jackson Laboratory) and NODShi. Cg-PrkdcscidIL2Rγtrunc (termed NOG, generated from the Central Institute for Experimental Animals) mice, (3, 4) which are depleted in T, B, and natural killer cells. Potent stem cells and immunocompromised mice must be used in hu-mice model to promote human cell engraft-ment. Hu-mice model harbor a human immune system; therefore, they are focused on the myelopoiesis system (5, 6), with advances in the establishment of lymphopoiesis in hu-mice developed via technical improvements, such as the transplantation of fetal bone marrow (BM), liver, and thymus (BLT) (2, 7-9). Human leukapheresis peripheral blood (LPB)-mononuclear cells (MNCs) are one possible cell source for developing hu-mice; however, these cells lead to acute graft-versus-host disease (aGVHD) due to T cell expansion. The long-term study of lymphoid lineage cells, including T cells, has failed to develop a hu-mice model using human PB-MNCs. Using CD34+ HSCs from many origins, such as umbilical cord blood (CB), BM, fetal liver, or PB, hu-mice have been established. An advanced protocol including HSCs can lead to a stable hu-mice model (10). Despite such advances, several issues remain when creating lymphoid lineage cells, including major histocompatibility complex (MHC) antigen specificity in humans and mice and B cell maturation in the hu-mice model. Because of preclinical needs that are not met by hu-mice, which can directly assess human immune responses, in vivo settings have been rapidly increased (11, 12), and a hu-mice model using more convenient and potent HSCs must be developed. Previous studies have revealed important biological information, such as cell identification regarding HSC origins and appropriate selection to apply in the model for preclinical purposes and better understanding of the key factors for generating a hu-mice model (1, 13). Among several issues, less potent HSCs and poorly developed lymphoid architecture are the main obstacles in developing hu-mice containing human lymphoid lineage cells (14). To successfully develop a lymphoid system in hu-mice model, biased HSCs committed to lymphoid lineage cells and cytokines and maintenance of the tolerance of host immune cells are required (15, 16). Transcriptional regulators are the main factors for determining the capacities of cell fate and differentiation, with previous studies focused on the identification of transcription factors (TFs) that govern lymphoid development and cell differentiation using genome-wide expression profiling (17-19). Limitations associated with the acquisition of human fetal BLT exist when generating hu-mice possessing lymphoid lineage cells. Thus, an understanding of the HSC properties related to TF expression is required. Although TFs are important for the enrichment of stem cells in the further development of hu-mice, there are no data for comparisons between stem cells, including CB and LPB cells. Thus, we investigated the expression levels of TFs involved in lymphopoiesis in CB-CD34+ cells and explored the expression of TFs of lymphoid lineage cells including RUNX1, Wnt5A, Notch3, HLA-DQB1, PU.1, ID2, and E12 between both LPB-CD34+ cells and CB-CD34+ cells and MNCs.
In the present study, we established a hu-mice model using CB-CD34+ cells and found that these cells displayed increased transcriptional expression of the lymphoid lineage cell program, contributed to establishing a hu-mice model and could be used as a potent cell source for reconstituting the hematolymphoid system. Hu-mice established with CB-CD34+ cells could be used as an in vivo platform for investigating the immune response against lymphocytes, compared to hu-mice established with LPB-CD34+ cells, which have low expression of TFs for lymphoid lineage cells.
Go to : 
Materials and Methods
Humanized mice
NOD.Cg-PrkdcscidIL2tmlWjl/Szj (NOD-SCID IL-2rgnull and NSG) mice were purchased from The Jackson Laboratory (Bar Harbor, ME, USA) and bred under pathogen-free conditions in the Department of Laboratory Animals at the CHA University of Korea. All animal experiments were approved by the Institutional Animal Care and Use Committee of the CHA University of Korea. To establish a hu-mice model, 1×105 CD34+ cells were intravenously injected into 3-week-old NSG mice that were sublethally irradiated with 300 cGy of total body irradiation 24 h before cell injection. At 12 weeks after cell injection, FACS analysis was performed to confirm human cell engraftment in the PB, spleen, and BM. Mice were monitored daily for symptoms of GVHD, including a ruffled coat, hunched back, weakness, and reduced motility. Mice were monitored for 180 days after transplantation for survival.
Preparation of human CD34+ cell
All experiments were performed with authorization from the Institutional Review Board for Human Research at CHA University of Korea. CB and LPB samples were obtained from the Cord Blood Bank at the Bundang CHA Hospital of Korea (protocols 1044308-201702-BR-017-03 and 1044308-201803-BR-014-02). CD34+ cells from CB and LPB were isolated with magnetic-activated cell sorting (MACS) using an anti-human CD34 progenitor cell isolation kit (Miltenyi Biotec, Germany). The experimental information regarding donors with numbers of CD34+ cells enrolled in the present study are listed in Table 1.
Table 1
Information of CD34+ cells from donors
![]()
Real-time qPCR
Total RNA was extracted from human cells using RNAiso Plus reagent (9109, Takara). cDNA was synthesized using a reverse transcriptase kit (RT200, Enzynomics). RT-qPCR was performed with SYBR Green (RT500M, Enzynomics) using a Real-Time System (Bio-Rad, CFX96TM). All data were normalized to glyceraldehyde-3-phosphate dehydro-genase expression. Information on the primer sets (Biosearch Technologies, Novato, CA) used in the present study is listed in Table 2.
Table 2
Primers for quantitative RT-PCR
![]()
Flow cytometry and immunocytochemistry
Blood, spleen, and BM cells from hu-mice were stained with antibodies and analyzed using a BD Accuri C6 Plus (BD Biosciences). The antibodies used to detect human cells included APC-conjugated anti-human CD45 (555485, BD PharmingenTM), FITC-conjugated anti-mouse CD45 (553080, BD PharmingenTM), APC-conjugated anti-human CD4 (555349, BD PharmingenTM), PE-conjugated anti-human CD8 (555367, BD PharmingenTM), PE-conjugated anti-human CD19 (302208, BioLegend), and PE-conjugated anti-human CD33 (303403, BioLegend). For immunocy-tochemistry, anti-CD45 (ab8216, Abcam) and anti-CD34 (MAB72271, R&D Systems) antibodies were used, and proper isotype-matched IgG antibodies were used to detect the primary signals. Flow cytometric data were analyzed using the CSamplerTM Plus software program (BD Bio-sciences).
Statistical analysis
All results are presented as the mean±standard error of the mean (SEM). Statistical analyses were performed using the Mann–Whitney U test for comparisons between two groups. Values of p<0.05 were considered to indicate statistical significance. GraphPad Prism version 5 software (GraphPad software) was used for analysis.
Go to :
Results
Successful hu-mice model established using potent CB-CD34+ cells
We previously developed a functional hu-mice model to study crosstalk in the immune response in vivo (13, 20). To understand the lymphoid biology of hu-mice, we investigated whether generation of the lymphoid system in hu-mice is affected by HSCs and CB- and LPB-CD34+ cells from various origins. We established a hu-mice model that contained human HSCs using CB- or LPB-CD34+ cells. MACS-sorted CB- and LPB-CD34+ cells were injected into irradiated 3-week-old NSG mice. At 8 weeks after cell injection, we found that the pan-hematopoietic marker CD45 was expressed in cells in the PB, BM, and spleen in both groups at least 20% of the time, suggesting a humanized condition (in CB-CD34+ cells: PB=37.7±8.6%, BM=44.6±7.3%, and spleen=51.4±8.9%). The hu-mice that received the LPB-CD34+ cells were divided into two groups for human cell engraftment, i.e., high-frequency and low-frequency groups. In the high-frequency group, PB=81.9±12.0%, BM=95.1±0.8%, and spleen=20.0±12.0%, whereas in the low-frequency group, PB=1.6±1.3%, BM= 11.4±10.6%, and spleen=5.9±5.7% (Fig. 1A). Inconsistent with mouse HSCs, human HSCs have individual variation with functional properties leading to different potencies for generating lymphoid and myeloid lineage cells in hu-mice (13, 20, 21), and these distinct differences in the human cell reconstitution of hu-mice were unevenly observed depending on the cell origin from donors. However, regardless of the origin of the HSCs, CD45+ hematopoietic cells were detected in the PB, implying successful generation of hu-mice (Fig. 1B). We next examined whether the generation of myeloid and lymphoid lineage cells was affected by cell origin and found that the pan-myeloid marker CD33 was stably expressed in cells in the BM and spleen of hu-mice (in the BM, CB=45.1±12.5%, and PB= 21.1±17.5%, whereas in the spleen, CB=12.7±3.1%, and PB=41.4±22.1%). In B cells, CD19 was highly expressed in the BM and spleen in mice generated with CB-CD34+ cells compared to those generated with LPB-CD34+ cells (Fig. 1C). Both CD33- and CD19-expressing cells showed patterns similar to that of CD45-expressing cells with regard to high and low frequencies. CD3+ T cells were rarely detected in the BM and spleen of hu-mice that received LPB-CD34+ cells; however, CD3+ T cells were successfully induced by CB-CD34+ cells in the BM and spleen of hu-mice (in the BM, CD4=15.3±3.6% and CD8=5.9±3.6%, whereas in the spleen, CD4=20.0±4.8% and CD8=12.6± 5.0%) (Fig. 1D), suggesting the specificity of CB-HSCs for the reconstitution of lymphoid lineage cells. Next, we investigated the expression of CD34, an HSC marker, in hu-mice to confirm that HSC homing in the BM exists in hu-mice. As shown in Figure 1E, the PB, BM, and spleen of hu-mice receiving CB-CD34+ cells displayed high expression of the CD34 phenotype compared to those of hu-mice receiving LPB-CD34+ cells. Consistent with CD45+ cell results, PB-CD34+ cells in the PB and BM but not in the spleen were divided into two groups for human HSC homing, i.e., high-frequency and low-frequency groups (in the high-frequency group, PB=99.9±1.1% and BM=99.6±0.1%, whereas in the low-frequency group, PB=0.7±0.3% and BM=0.8±0.3) (Fig. 1E). CD34+ cells in the CD45+ cell population were also clearly detected in the BM of hu-mice receiving PB-CD34+ cells (Fig. 1F), suggesting that CB-CD34+ cells had more potent stem cell function in generating the hu-mice model and that not all PB-CD34+ cells failed in homing and engraftment due to the poor functioning of HSCs.
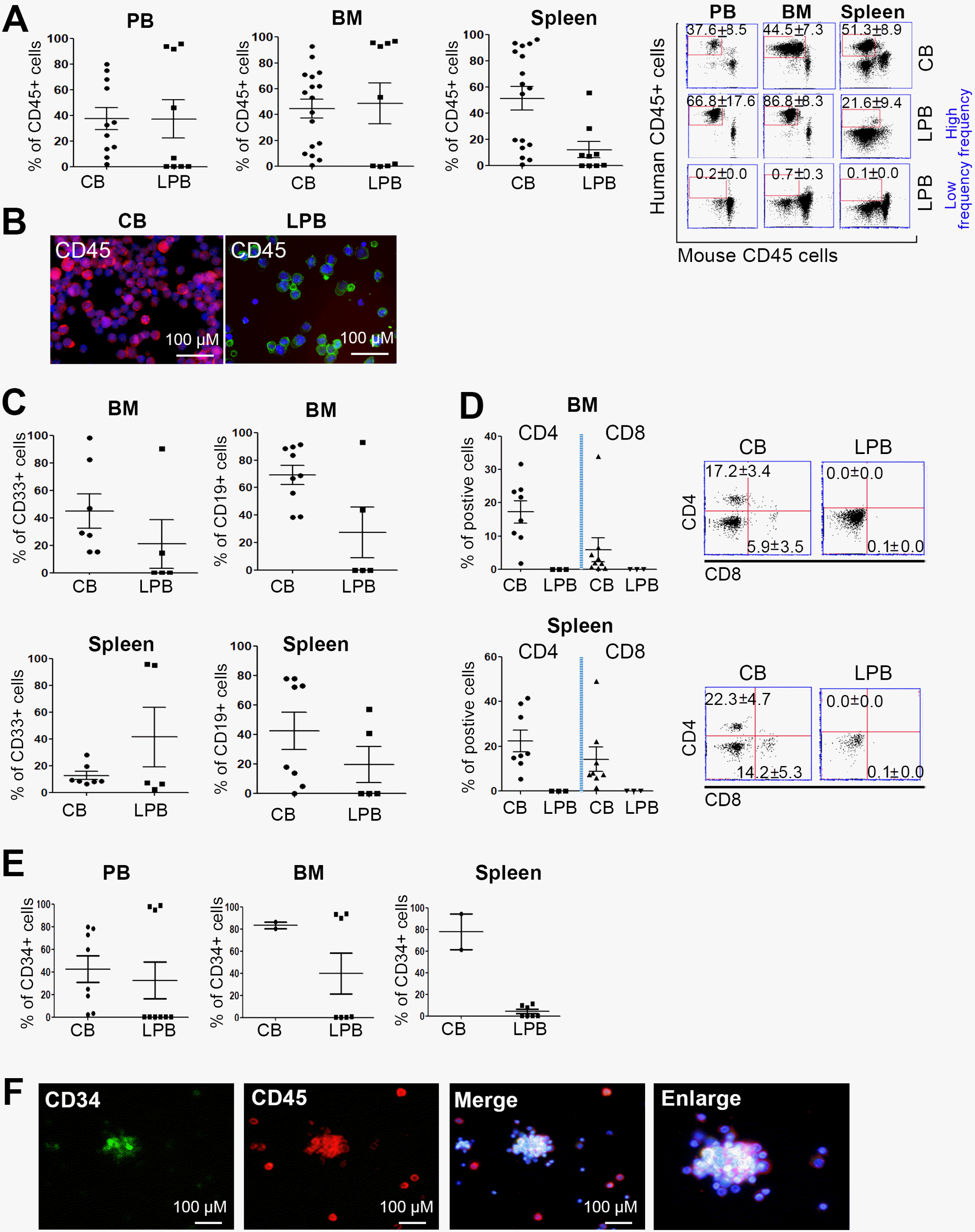 | Fig. 1Successful establishment of hu-mice using CB- and LPB-CD34+ cells. (A) CD45+ pan hematopoietic cells were stably engrafted into xenografted mouse tissues including PB, BM, and spleen (n=11∼17 in CB, n=9 in LPB). (B) Fluorescence microscopic imaging of CB and LPB cells, with CD45 expression. DAPI: blue. Colors: CD45. Scale bar=100 μm. (C) Myeloid lineage marker CD33 and B cell marker CD19 were evaluated and their frequency determined using CB and LPB cells. (In CD33, n=7 in CB, n=5 in LPB; In CD19, n=9 in CB, n=5 in LPB) (D) Markers for lymphoid lineage cells CD4 and CD8 were rarely detected in LPB cells. (n=8 in CB, n=3 in LPB) (E) Stem cell marker CD34 was highly expressed in hu-mice tissues at 8∼18 hr post CD34+ cell injection. (n=2∼8 in CB, n=7∼9 in LPB) (F) Images from at least two independent experiments are shown. CD34 (green), CD45 (red), and DAPI (blue) are demarcated in BM cells from hu-mice. Scale bar=100 μM.
|
TFs for lymphoid lineage cells were highly expressed in CB-CD34+ cells
We examined the enrichment of TFs in lymphoid lineage cells derived from CB-CD34+ cells. The identification of transcriptional activity in HSCs, which controls cell fate and multipotent differentiation, is of great importance for understanding the biology of lymphocytes. In particular, some defects in the HSCs of hu-mice, including lymphoid lineage cells, can limit the contribution to a long lifespan (17, 19). Because HSCs may not fully sustain their function under various conditions, real-time PCR analysis of TFs involved in lymphopoiesis was performed to gain insight into the molecular mechanisms that underlie these deficits. For MNCs, we found that the levels of TFs in lymphoid cells were significantly increased in CB-MNCs. The Runx TF family member RUNX1 is known to function as a key regulator of normal HSCs and lymphoid progenitors (22). RUNX3 is also a main TF that mediates cytotoxic lymphocyte differentiation, showing strong relevance to lymphoid lineage cells (23). Wnt5a, which is a member of the Wnt family, and Notch3 drive the formation of lymphocytes and can enhance the Th1 adaptive immune reaction (24-26). ID2, E12, and HLA-DQB1 are relevant to stimulating lymphoid lineage cells. ID2, as a TF in lymphoid cell precursors, is pivotal in the development of natural killer cells. E12 and HLA-DQB1 can allow commitment to B cell lineage cells and maturation (27-34). The TFs of the lymphoid lineage, RUNX1, Wnt5A, Notch3, PU.1, ID2, and E12, were significantly expressed in CB-CD34+ cells compared to LPB cells, regardless of the frequency in engraftment. Additionally, hu-mice that received LPB cells with high expression of these genes displayed high engraftment compared to hu-mice that received LPB cells with low expression of these genes (Fig. 2A and Table 1). CB-CD34+ cells from donors that induced high engraftment in hu-mice displayed high TF expression, especially expression of RUNX1, Notch3, HLA-DQB1, ID2, and E12, compared with those in hu-mice with a low human cell engraftment frequency. To further examine whether the expression of these genes was also increased in HSCs, we isolated CD34+ cells from both CB samples and LPB samples and found that the levels of ID2, Wnt5A, Notch3, HLA-DQB1, and E12 were significantly increased in CB-CD34+ cells, suggesting strong relevance to the importance of TFs in HSCs during hu-mice development (Fig. 2B). Recently, many transcriptomic approaches, such as single-cell analysis, have identified the critical factors involved in the cell fate commitment of HSCs and progenitors. TFs, acting as regulators of cellular movement, play an important role in regulating hematopoietic cell fate decisions (35, 36). Consistent with previous papers, these results showed that CB-CD34+ cells are beneficial for establishing a hu-mice model.
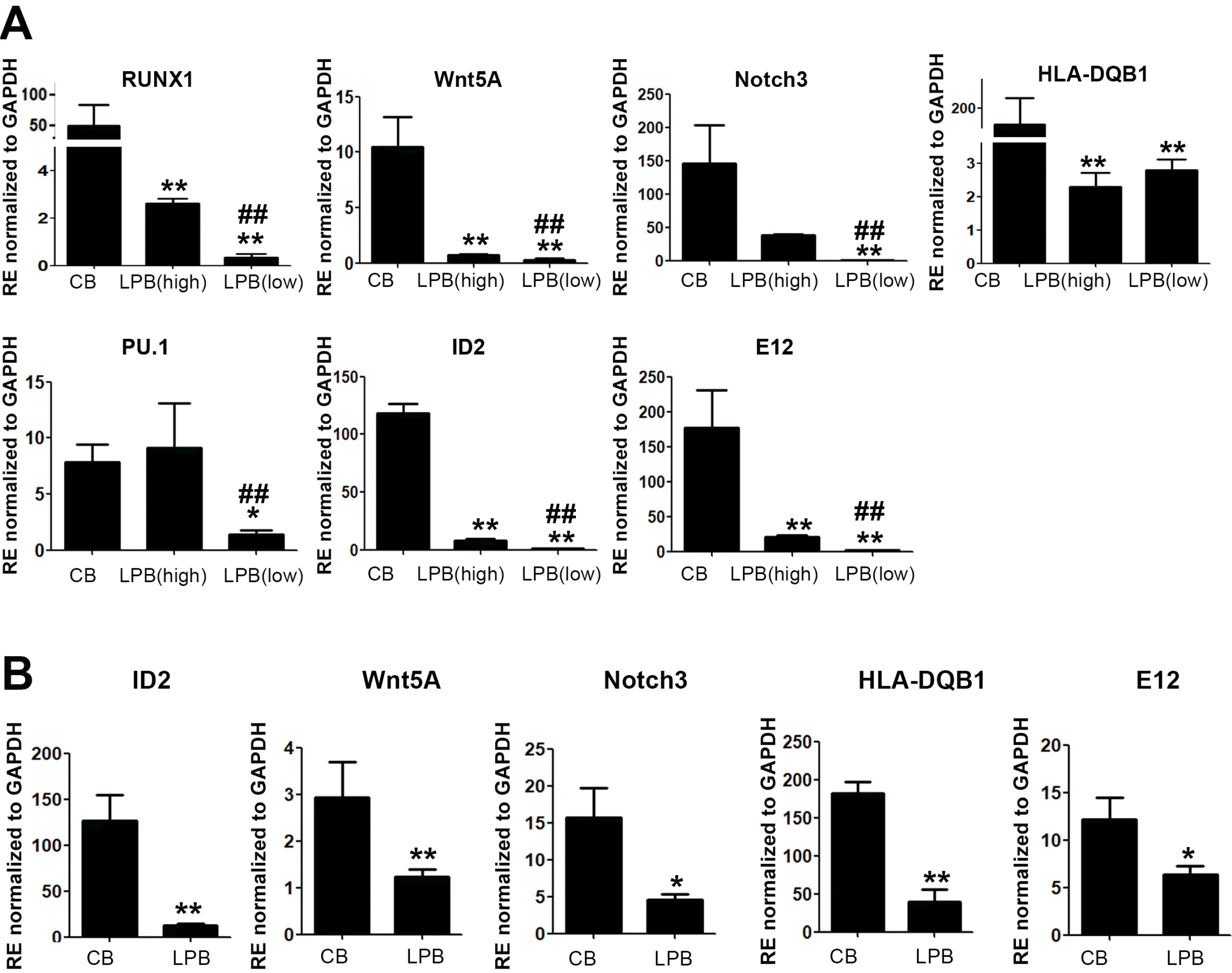 | Fig. 2Transcription factors involving lymphoid lineage cells were highly increased in CB-MNCs as well as in CD34+ cells. (A) qRT-PCR analysis for CB- and LPB-MNCs. Results are shown as mean±SEM for n=3. Each with technical duplicates. * and #p<0.05, ** and ##p< 0.01. Mann–Whitney U test with two-sided p values. * depicts significance for CB and # is the comparison between high and low frequency groups in LPB. (B) qRT-PCR analysis for CB- and LPB-CD34+ cells. Results are shown as mean±SEM for n=3. Each with technical duplicates. *p<0.05, **p<0.01. Mann–Whitney U test with two-sided p values.
|
Hu-mice model using CB-CD34+ cells displayed longevity
To investigate longevity of hu-mice, survival rate of mice was observed until 190 days along with the occurrence of GVHD. GVHD is a reaction to host tissues, occurs after allogeneic stem cell transplantation and is defined by the occurrence of symptoms within 3 weeks (37, 38). This leads to early lethality in hu-mice that is not acceptable for screening the immune response mediated by lymphocytes. Thus, we sought to address the stable longevity of HSCs in engrafted mice. A total of 33 heads from mice that had received human CB-CD34+ cells from six donors and 43 heads from mice that had received LPB-CD34+ cells and MNCs from six humans were utilized to examine longevity. We found that hu-mice receiving LPB-MNCs maintained stable survival until 110 days and survived until 190 days after CB-CD34+ cells were injected (Fig. 3A). Regardless of cell type, hu-mice receiving CB cells had longer longevity than those receiving LPB cells (CB-CD34+ cells=42.4% and LPB-CD34+ cells=29.1%) (Fig. 3B). In present study, we showed the potency of CB-CD34+ cells, which highly express TFs involved in lymphopoiesis, for reconstituting lymphoid lineage cells in a hu-mice model and these cells induced higher longevity than LPB-CD34+ cells.
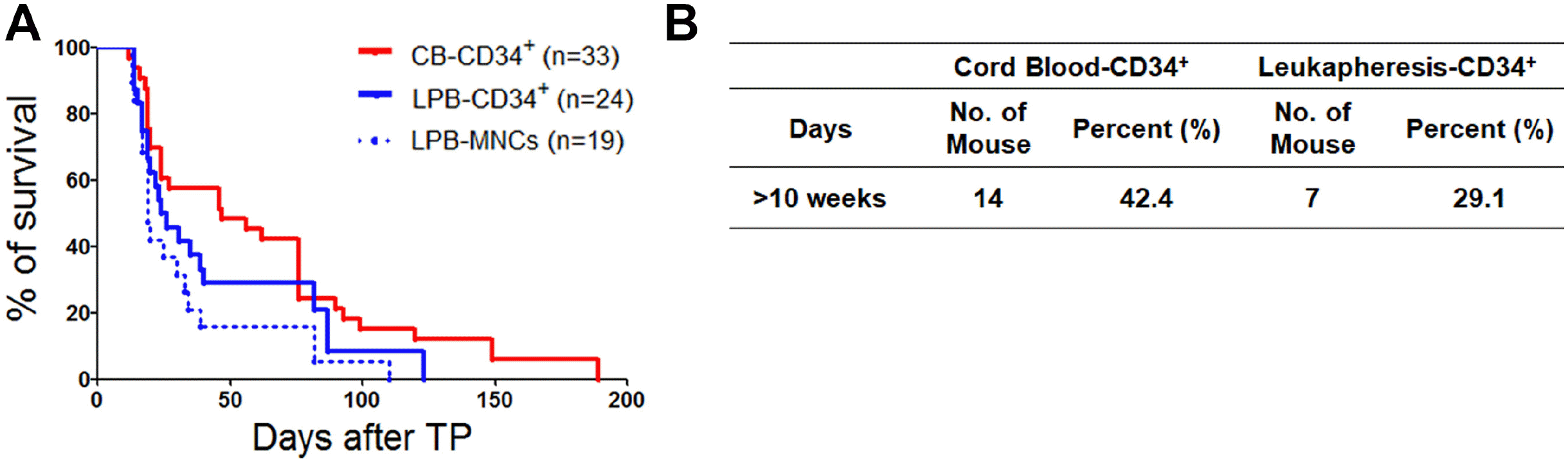 | Fig. 3Comparison of the survival rate in hu-mice over time. (A) Mice injected with CB-CD34+ cells showed the highest lifespan compared to that of the other groups, suggesting CB-CD34+ cells might maintain the hu-mice model with low mortality. LPB-MNC as a control group (B) The percentage of survival rate for (A) in both CB-CD34+ and LPB cells including CD34+ cells and MNCs.
|
Go to :
Discussion
Hu-mice are regarded as a valuable in vivo platform for biomedical research and translational medicine. Many investigators have developed hu-mice with hematolymphoid lineage cells using advanced methods such as BLT transplantation and the activation of dendritic cells for T cell education (14). However, lymphoid cells in the thymus of hu-mice are stimulated with minimal guidance from human cells, and T cells rarely bind to the human MHC complex, resulting in diminished T cell activation. Most previous studies have focused on modulating the hu-mice microenvironment to generate the hematolymphoid system via tissue transplantation; however, the characteristics of CD34+ HSCs remain to be clarified. The identification of transcriptional regulation, which controls tissue-specific functions, is pivotal for understanding the fate of cell lineages. Recently, emerging studies have addressed the identification and function of novel TFs in specific lineage cells, such as T cell and non-T cell subsets, to unravel the functions of these TFs affecting differentiation or crosstalk with other cells in immune networks (19). Gene expression patterns can reflect ontogeny, and HSCs and T cells share a similar transcriptional program in regard to activation mechanisms, leading to lymphocyte development or differentiation (39). To determine the effects of TF expression on HSCs, we focused on the TFs of lymphoid lineage cells to generate hu-mice containing lymphoid lineage cells. We randomly selected several genes uniquely expressed in lymphoid lineage cells (24-34) and then performed PCR analysis of MNCs and CD34+ MACS-sorted HSCs. The expression of these genes in lymphoid lineage cells derived from HSCs and MNCs partly influenced the successful engraftment of hu-mice by enriching specific TFs. Based on gene expression profiling, we determined that the primary TFs in HSCs were relevant to cell-surface molecules, and cell identification and advanced technological breakthroughs, such as single-cell RNA sequencing, could be applied to define HSC heterogeneity based on HSC transcriptomic profiles. In hu-mice, the reconstitution of lymphoid lineage cells remains to be explored. Human BLT tissues are essential for the reconstruction of the mature lymphoid system in hu-mice. Although BLT transplantation allows for T cell maturation, education, and development in hu-mice, reconstruction of the hematolymphoid system in hu-mice fails if the infused HSCs only allow for primitive hematopoiesis due to the inadequate potency of HSCs. Thus, the identification and isolation of potent HSCs are central to establishing hu-mice. Previously, we investigated ALDHdimCD34+CD38− cells as a reliable marker for establishing a xenograft mouse model of leukemia (13). To build a leukemia model with longevity, ALDHdim-expressing leukemic stem cells in the LPB were superior in terms of survival compared to BM cells. Here, we showed that CB-CD34+ cells with TF enrichment involving the activation, maturation, and differentiation of lymphoid lineage cells were an optimized cell source in hu-mice reconstituted with lymphoid lineage cells compared to LPB cells. In the analysis of LPB-CD34+ cells, those with a high frequency of engraftment displayed a significant difference in TF expression from those with a low engraftment frequency. Even though we did not perform PCR using CD34+ cells from many LPB samples (Fig. 2B) owing to limitations, we found the importance of the genomic status in HSCs. Runx1 is an essential factor in the development of definitive hematopoiesis and Runx1 inactivation is relevant for targeting early thymic progenitors (22). Wnt5A, HLA-DQB1, ID2, and E12 were strongly involved in lymphoid lineage cell development and activation. The enrichment of these genes in lymphoid lineage cells derived from HSCs was associated with successful generation of hu-mice with a lymphoid system that had a low incidence of aGVHD, resulting in longevity. This will allow us to perform biomedical and translational medicine research, such as drug screening and stem cell transplantation, in vivo. In conclusion, we developed a hu-mice model using CB-CD34+ cells that express TFs of lymphoid lineage cells. This hu-mice model could serve as a suitable in vivo platform for studying the immune response of lymphoid immune cells in the context of a low aGVHD incidence.
Go to :
 XML Download
XML Download