

 PDF
PDF Citation
Citation Print
Print


Introduction
For several decades, developmental biologists have studied aspects that can control stem cell behavior, such as differentiation and self-renewal, along with certain tissue lineages. To understand human biology, researchers have attempted to create models of the human developmental stages in vitro using human pluripotent stem cells (hPSCs), including human embryonic stem cells (hESCs) and induced pluripotent stem cells (iPSCs), even though established animal models such as fruit flies and mice already exist. The concepts and insights gained from the model organism systems have resulted in a significant shift from the use of stem cell models to three-dimensional (3D) structural organoids with organ-like properties, which serve as an ideal platform for studying stem cell development, tissue regeneration, and diseases in humans (1-3). However, generating perfect organs in vitro has been challenging, and scientists are trying to optimize this process.
As one of the promising hPSCs for organ formation, disease modeling, and applications in vitro, hESCs are unfortunately ethically controversial and limited in supply (4). On the other hand, human iPSC-derived organoids have overcome the limitations associated with using human tissue samples, and they maintain the unique cell types found in the long-term environment in vivo. Advances in extracellular matrix (ECM) biology (5, 6) have made it possible to observe in vitro morphology in a 3D laminin-rich matrix (7). Using this culture method, various organ systems have been generated by growing human iPSCs on an ECM-based hydrogel (8). Since the publication of this strategy, representative methods for developing of intestinal organoids (9), optic-cups (10), liver buds (11), cerebral organoids (12), gastric organoids (13), lung organoids (14), nephron organoids (15), ductal organoids (16), inner ear organoids (17), cardiac organoids (18), skin organoids (19), blood vessel organoids (20), and models of other organs have been established (Fig. 1).
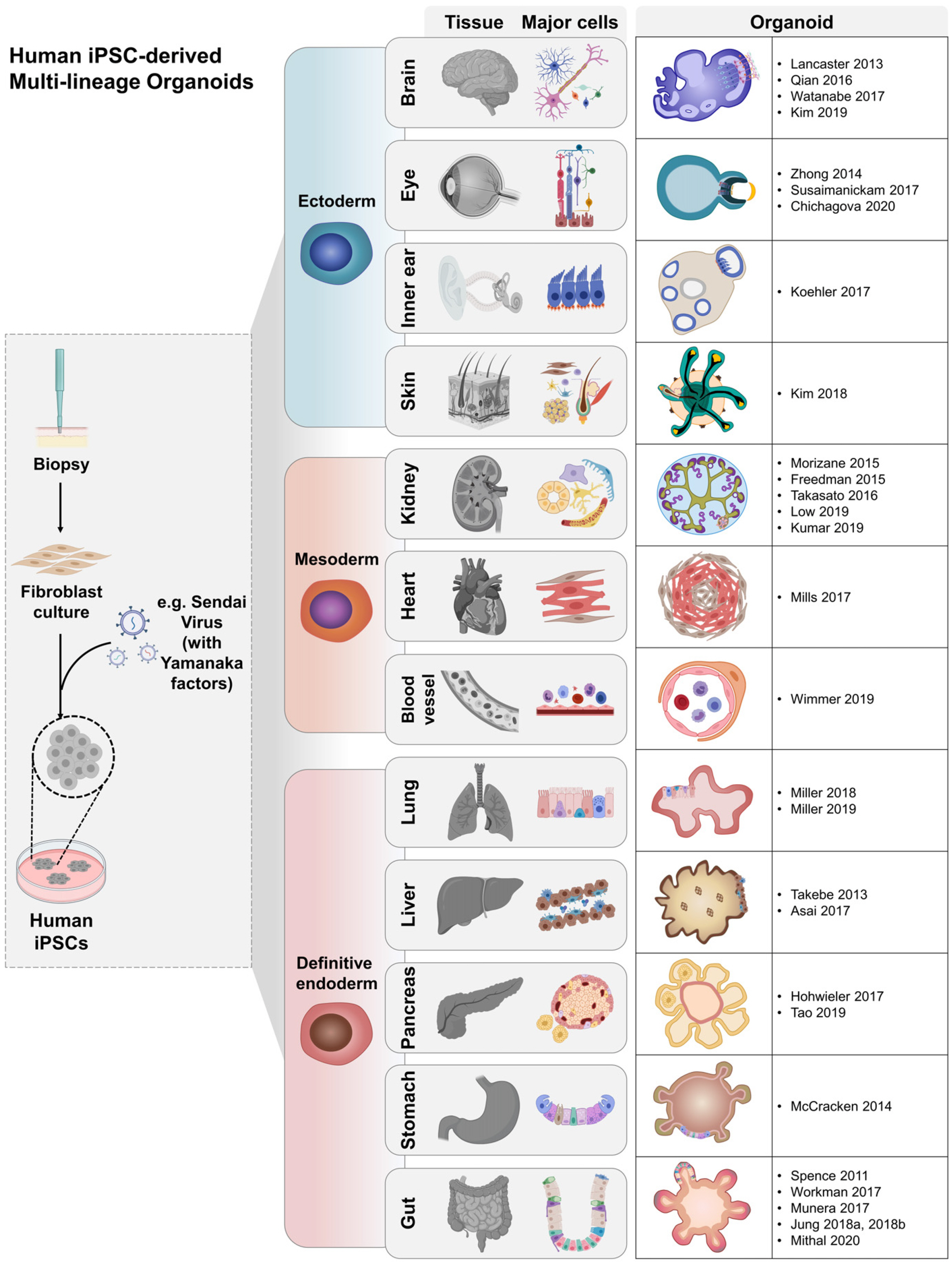 | Fig. 1Summary of the generation of iPSC-derived multilineage organoids. The generation of human iPSCs is depicted, and several organoid systems are listed along with their representative published protocols for genera-tion. This figure was created using BioRender (https://biorender.com/).
|
Organoids have been acknowledged as an important platform for drug screening (21) and have the potential to be used for studying the effect of long-term alternative therapies in regenerative medicine (22). Additionally, organoid technology can be used for numerous purposes by coupling it with the following: genome editing with clustered regularly interspaced short palindromic repeats (CRISPR-Cas9) tools to obtain important insights into genetic disorders (23); co-culturing with pathogens for studying infectious diseases (24); and cancer modeling to understand cancer pathogenesis, development, and progression (25, 26). Principally, organoids serve as exploration tools for understanding the processes underlying human development and diseases (13). However, despite the advances made, most of the human iPSC-derived organoids remain immature in cultures owing to the absence of important features found in adult tissues (27). Therefore, robust in vitro maturation methods that provide the required cellular and environmental constituents for producing functional and mature organoids are needed.
This review will explain the recent efforts at increasing the complexity of organoids and describe the current potential of using mature organoids in applications as diverse as bioengineering, disease modeling, drug discovery and regenerative medicine. We discuss the approaches to build the next-generation organoid, platform and highlight the challenges that need to be addressed for the organoid technology to reach its full potential in the field of regenerative medicine (28).
Go to : 
Generation of Organoids
Organoids are generated by leveraging the self-organizing and self-patterning properties of homogeneous cell populations (3). They are ex vivo “mini-organs” that contain fully differentiated cell types in the same spatial organization, as in the native organ. For the successful production of organoids, factors such as organ/tissue-specific cell types, proper culture conditions, signaling pathways responsible for regulating differentiation, and ECM (that influences the final properties of the organoids) must be considered.
Initial stem cell types
Adult stem cells (ASCs) (24, 29) and tissue-specific stem cells derived from hPSCs (9, 10, 12, 13) differ in their developmental potential, and thus follow their respective differentiation pathways in vitro. Therefore, depending on the purpose for which organoids are used, such as cell therapy, drug screening, or in studies of the basis of organ formation, the starting cell population is important (30). Additional areas that require further study include cell interactions during development, tissue biology, and translational medicine (1, 2). ASC-derived organoids are useful for research on the epithelial lining of the small intestine (29), large intestine (31) or stomach (32) which undergoes rapid regeneration, because these organoids exhibit characteristic intestinal epithelial regeneration and homeostasis even when cultured in vitro. However, there are practical limitations associated with obtaining enough ASCs. In contrast, iPSC-derived organoids have been generated by modulating various differentiation pathways involved in organ and tissue development, therefore iPSCs are usually used in studies that involve the recapitulation of processes associated with organ/tissue development in vitro. However, there have been hurdles in obtaining mature, “adult-like” cell types in hPSC-derived organoids (9). Additionally, co-culture of organoids with other cell types is required such as immune cells, nerve cells, and endothelial cells to improve the functional maturity and complexity of organoids (11, 33).
Signaling pathways involved in organoid differentiation
Most organoids are derived from a population of starting cells exposed to a particular morphogen at a defined point of time, resulting in the activation of the desired developmental signaling pathway. Notably, iPSC-derived organoids require exposure to specific growth factors at a precise time for differentiation into the target organs. Each organ is derived from three germ layers formed during embryonic development that undergo particular differentiation pathways. Diverse organs are formed from the same germ layer depending on the inductive signal. Precisely using these pathways to form organoids, along with the various combinations and concentrations of factors, and timing can cause different outcomes. Typically with intestinal organoids, the hindgut is formed from the posterior endoderm in response to induction by fibroblast growth factor (FGF)/Wnt (9). Gastric organoids induce foregut formation by inhibiting bone morphogenetic protein (BMP) signaling and generate the posterior foregut via retinoic acid signaling (13). Subsequently, the epidermal growth factor promotes the development of antral gastric organoids. Organoids can differentiate into fundic gastric organoids in response to continuous exposure to these conditions–activation of Wnt signaling; mitogen-ac-tivated protein kinase inhibition; BMP activation (34). Human colonic organoids (hCOs) can be produced by regulating BMP signaling after the formation of hindgut spheroids (35). iPSC-derived retinal organoids can be driven to differentiate into retinal tissue in vitro by modulating signaling pathways involving BMP4 and insulin-like growth factor 1 (36). FGF and sonic hedgehog proteins are required to promote the formation of lung organoids (14).
ECM for 3D culture
Currently, organoids are cultured using an ECM to build 3D culture environments. In many reports, the ECM from Engelbreth-Holm-Swarm murine sarcoma, Matrigel, has been mainly used for culturing cells (11, 17). Matrigel has been widely used in epithelial cell culture, but batch- to-batch variability impedes reproducible in vitro studies, and its use in regenerative medicine is limited due to mouse tumor cell origins (5, 7). Therefore, strategies for 3D cultures that use synthetic hydrogels (37) and Type I Collagen gel (38) as alternatives have been reported. Moreover, tissue-specific ECMs were produced using extracellular matrix hydrogels derived from decellularized tissues, increasing the possibility of using organoids clinically (39). In contrast, 3D suspension cultures without solid scaffolds are used to culture renal (40), retinal (41), and cerebral organoids (42). The generation and characterization of cerebral (43), lung (44), and renal organoids (45) using an air-liquid interface method has been demonstrated.
Go to :
Development of iPSC-Derived Organoids
Since 2010, numerous papers have described the generation of various iPSC-derived organoids that represent human tissues. These organoids can be divided into ectoderm, mesoderm, and endoderm according to the lineage-specific differentiation process to describe organoid formation. Brain, eye, inner ear, and skin organoid represent the ectoderm. The Lancaster group first developed the iPSC-derived cerebral organoid culture system, which has shown it can reproduce brain development (12). Models in which cerebral organoids containing neurons and glial cell types, including oligodendrocytes, have been created offer new opportunities to examine processes associated with early neuronal development and diseases (46-48). Even more, to predict the central nervous system permeability of drug compounds, a human choroidal plexus organoid with cerebrospinal fluid secretion was established and could be used in brain homeostasis studies (49). Similarly, retinal and corneal organoids have been generated that contain photoreceptor cells that respond to light stimulus; positive results have been reported following their use in cell therapy (10, 36, 50). Ear organoids that contain sensory neurons and cochlear hair cells have recently been developed (17). Further, skin organoids that simulate the complex structure of human skin have been created (19). In mesoderm, one of the three germ layers, kidney, heart and recently blood vessels were developed. Renal organoids containing nephrons have been established which have recently been vascularized using ECM and suspension culture methods (15, 23, 40, 45, 51). Blood vessel organoids, including epithelial cells and pericytes have been generated; the organoids exhibit self-assembly and vascular tree formation when implanted into mice (20). Importantly, cardiac organoids were developed by forced fusion via a biotechnology approach, and are being actively used in drug screening (18). In endoderm, intestinal organoids were produced and developed in the early stages of organoid advancement (9). Progress has been made in developing mature human intestinal organoids (hIOs) and hCOs by employing various technologies (33, 35, 52-54). Pancreatic organoids can secrete insulin and are evolving as an important model for the treatment of diabetes (16, 55). As reported, mesenchymal stem cells and umbilical vein endothelial cells are used to establish liver organoids, and albumin is produced from vascularized liver organoids are used in drug metabolism studies (11, 56). Lung organoids that constitute the basic bronchiole-like structure and alveoli have been developed (14, 57). Additionally, gastric organoids containing antral or fundic organoids, which differ in the structural location and function of the stomach, have been generated (13). Taken together, organoids of various tissues have been produced, and their number is growing rapidly (Fig. 1). Furthermore, novel multi-organoid systems (boundary-organoid, hepato-biliary-pancreatic) have been reported (58), and mesenchyme-free organoids generated using iPSC-derived hIOs have also been developed (54).
Go to :
Enhancement of the Organoid Maturity
Co-culture of various cells
By stimulating cells with human developmental signals, it is possible to generate a variety of cell types present in vivo, such as neurons, vascular cells, and cells of the immune system (9). Recent studies have shown that hIOs are altered due to the enteric nervous system (ENS) when co-cultured with vagal neural crest cells (52). The co-cultured hIOs exhibited ENS integration effects regarding epithelial development; they also increased intestinal stem cell and transit-amplifying cell counts, and reduced the number of the differentiated absorptive and secretory intestinal cell types. This suggests that ENS could influence the differentiation trajectory of the hIOs. Furthermore, hIOs co-cultured with ENS may experience contractions like the peristaltic motion of the small intestine after implantation under the kidney capsule in vivo. It has been reported that intestinal cell-specific marker expression and functionality have improved in co-culture conditions with stimulated Jurkat cells or in response to treatment with interleukin-2 (IL-2), a major cytokine secreted by T lymphocytes (33). IL-2 was also found to maintain the hIO maturation effect when transplanted under renal capsules. Recently, the hIO maturation factor, IL-2, was reported to exhibit therapeutic effects on dextran sulfate sodium-induced ulcerative colitis (59). Co-culturing with endothelial cells and mesenchymal stem cells was attempted to improve the functions of human iPSC-derived hepatocytes in liver organoid models (11, 56). Mature organoids can be generated by introducing various cell types, such as nerve cells, immune cells, endothelial cells, and mesenchyme, i.e., cells surrounding the organ in vivo, besides improving the functional maturity and complexity of organoids (Fig. 2). In addition to using co-culture with organoids and other cell types to establish multilineage assembloids, it is also possible to create mutiregion assembloids model by fusing region-specific organoids of the same organ and controlling polarity or signal. This approach can explain fundamental questions about the actual organ formation process and disease mechanisms (60).
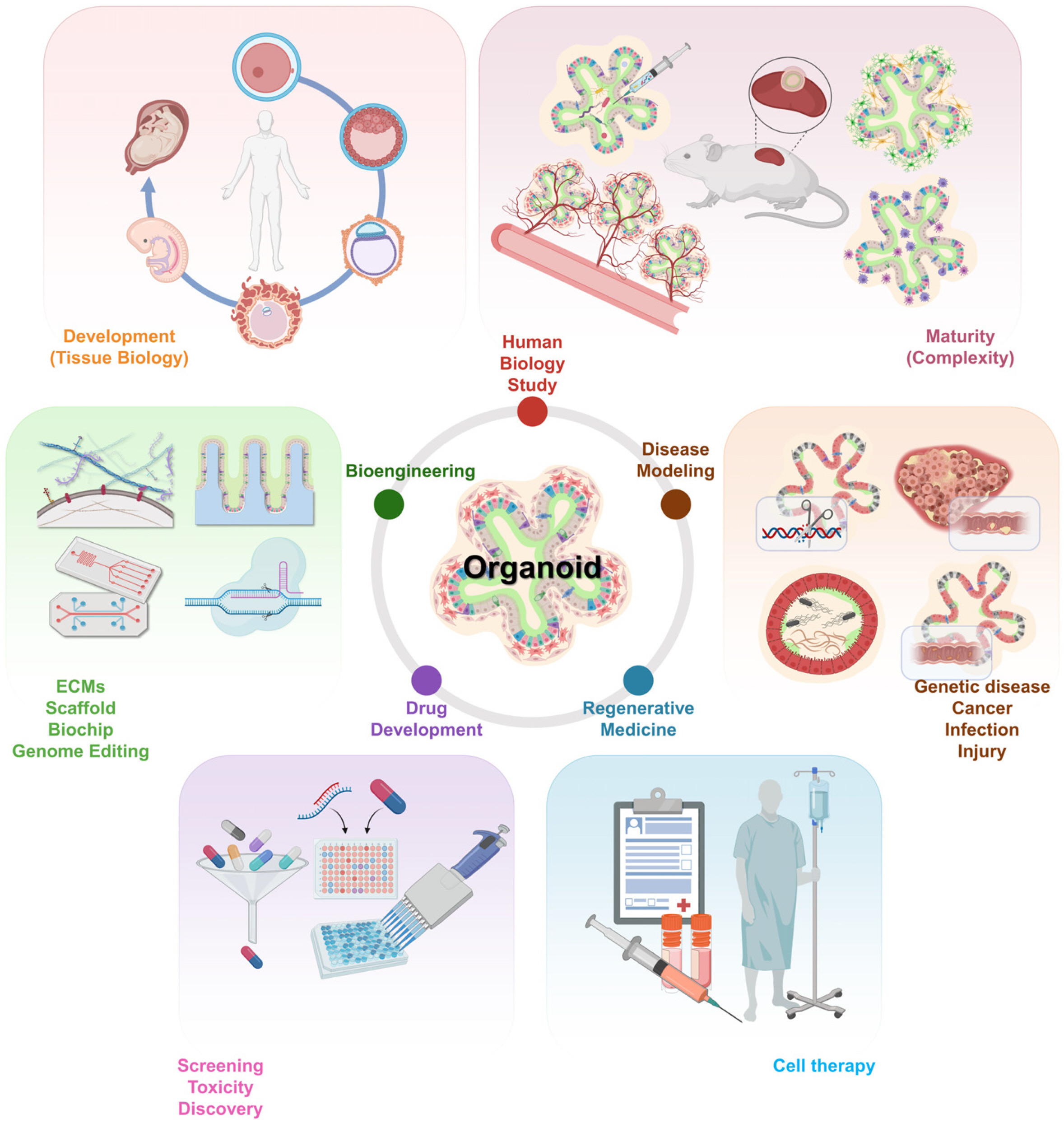 | Fig. 2The various applications of organoids. The scope of organoids in respective applications such as areas of human biology studies, disease modeling, regenerative medicine, drug development, and bioen-gineering has been illustrated. This figure was created using BioRender (https://biorender.com/).
|
In vivo transplantation into mammalian hosts
In many studies, organoids have been transplanted into immunodeficient mice to enable maturation in vivo (Fig. 2). There are two approaches to organoid implantation; these include ectopic and orthotopic transplantation. Ectopic transplantations have been performed in the kidney capsule (35, 61), epididymal fat pad (57), and intestinal mesentery (62), which are highly vascularized sites that have access to nutrients in vivo. A hIO has shown significant maturation after transplantation under the kidney capsule for several weeks (61). The transplanted hIO generated a crypt-villus structure similar to that in adult tissues, and the mesenchymal lineage cells were differentiated to form smooth muscle layers. With orthotopic transplantation, the microenvironment of the native tissue and the therapeutic potential can be studied. For example, an implanted cerebral organoid was maintained for the long term (∼8 months) after in vivo transplantation by intracerebral implantation, following which extensive axonal growth was observed (63). In addition, it has been confirmed that apoptosis decreases in the graft as vascularization takes place (64). Neuronal activities have been also recorded owing to the connection between the graft and the host. Collectively, the transplantation approaches could produce mature organoids and provide critical in vivo environments that influence the enhanced organoids. Although additional efforts are required to define the essential factors for further maturity, this approach made it possible to identify their potential in translational medicine.
Go to :
Organoid Applications
Organoids have great potential for drug development and cell-based therapies, which are further combined with various applications, such as genome editing tools and organs-on-a-chip technologies. Moreover, new applications allow creating advanced organoids, which could have tremendous potential in translational and regenerative medicine.
In vitro model for developmental study, disease modeling and testing
In principle, the generation of iPSC-derived organoids needs to strictly mimic human organ development. Therefore, the process should be required precise spatiotemporal signals and correct concentrations essential for cell differentiation and tissue assembly (14, 35, 57). Organoids as 3D models have contributed to an in-depth understanding of human tissue/organ biology; this approach is more realistic than two-dimensional (2D) cultures and mammalian models (65). Moreover, organoids have provided an easily accessible system for identifying organ formation and have opened new avenues for studying human developmental biology (Fig. 2).
Compared to the conventional 2D model, the organoid systems can better simulate histopathological characteristics by assembling various cells. In addition, genetic disorders can be recapitulated using genome editing technology, and in the case of infectious diseases, organoids can be directly co-cultured with pathogens, providing models for disease mechanisms and pathophysiology. Based on these advantages of organoid systems, certain diseases have been studied, such as genetic disorders (12, 21), host- pathogen diseases (13, 24), and cancers (25, 26). Organoids generated from patient-derived iPSCs should clearly recapitulate human pathophysiology to better predict the efficacy and toxicity of drugs at a tissue/organ level by reflecting unique clinical responses to drugs in individual patients. Disease-specific biobanks can be used as a source of samples to test powerful alternative tools for drug screening and precision medicine approaches (66) (Fig. 2). In particular, organoids have been used in a system to test drugs to treat of Zika virus infection (46), and to develop personalized medicine for cystic fibrosis (21) and colorectal cancer (67).
Improvement of organoid systems via bioengineering approaches
Human iPSC-derived organoids are generated by simulating a human developmental process, but their structure differs from that of real organs. In addition, since it is difficult to deliver growth factors that induce the in vitro maturation into 3D structured organoids, except for in vivo transplantation, there are challenges in the continuous culture process of expandable and organ-mimicking organoids. Therefore, to address these limitations, novel biotechnology approaches, including genome engineering, organ-on-a-chip platforms, and ECM-based scaffolds, have been proposed (37, 55, 68) (Fig. 2). A fluorescent reporter iPSC was created using CRISPR-Cas9 to expand the utili-zation of organoids, and the fluorescence of specific cell markers was observed during differentiation (53). Furthermore, LGR5-CreER knock-in colon organoids were generated for lineage tracing of LGR5+ intestinal stem cells (ISCs), and were transplanted into immunocompromised mice to observe the differences in stem cell cycling between species (28).
Organ-like bioengineered scaffolds are required for the cellular differentiation, organization, and activity, and they can also enhance the continuous culture process of organoids, to simulate the size and shape of actual organs by providing their physiological environment. For example, ISC cultures have maintained the shape of native small intestine with a crypt-villus structure when grown on a scaffold (69, 70). Besides, several studies have reported the use of chemically defined hydrogels (37) and collagen I (38) instead of Matrigel, to improve the growth of organoids and demonstrate clinical use. Applying hydrogels has led to developing photodegradable systems, whereby poly (ethylene glycol) hydrogels can also encapsulate embryonic stem cell-derived motor neurons and use infrared radiation for three-dimensional control. For physical micropatterning on the 3D tissue engineering scaffold, the structure within the hydrogel can be created using pulsed lasers and precisely controlled to enable the development of 3D cultured neural networks as well (71, 72). Alternatively, 3D bioprinting approaches that uniformly assemble cells in structures similar to those of adult organs, may decrease the variability of the organoid phenotypes and may be highly reproducible (73). The generation of organoids can be improved by growing microtubules on chips that mimic the scaffold and blood vessels, reproducing the required gradient of signal molecules (74). Organ-on-a-chip manufactured for organ-specific cell types can simulate a circulatory system with a microfluidic channel and has high reproducibility as an automated system. Organ-on-a-chip based on microfluidic technology allows cells to adapt to the culture environment of the chip system in which the medium is circulated within hollow microchannels (75). These devices can culture and maintain various types of cells simultaneously and predict drug response and toxicity at the organ- or body-levels, even as a multi-organ-on-a-chip. A multi-organ-on-a-chip, also known as a human-on-a-chip, contains various cell lines, including liver, lung, kidney, and adipose tissues, and a multi-channel 3D microfluidic system (76). Furthermore, an organ-on-a-chip is suitable for high-throughput systems including the drug or toxin screening, growth factor and/or signal identification, and may additionally study organ-organ and organ-vessel interaction (77).
Host-microbial communications in organoid culture
The human intestinal mucosal barrier and immune system definitively develop during late gestation and infancy periods, which are associated with the first exposure of commensal and pathogenic microorganisms in vivo (78). The interactions between organoids with commensal and pathogenic microbes and viruses play important roles in the physiology and function of the human body. Intestinal ecology affects important features such as digestion (79), metabolism (80), immune function (81) and brain development (82) through microbial colonization in early childhood. Therefore, introducing microbes into hIOs is necessary to understand colonization of various microorganisms in the human gut and their impact on early intestinal development. Especially gastrointestinal organoids which have niche components such as mucus layers will provide new insights into gastroenterological health and disease mechanisms. Several microbes have been microinjected into the lumen of closed hIOs to mimic intestinal host-microbial communications (83, 84) (Fig. 2). It was possible to observe the structural and functional changes in hIOs due to microbial colonization and confirm the interaction with microorganisms. Further research will broaden our understanding of the interactions of balanced microbiota with organs during human development (85), and to demonstrate the mechanisms of pathogenic microbial infection (86).
Potential applications of organoids in cell therapy
Organoids are an important component of cell therapy in regenerative medicine, and the implantation of organoids in animal models has been demonstrated with various approaches. For example, hIOs were transplanted into colonic injuries in mice, following which the colonic mucosal damage was regenerated after 4 weeks (28). In addition, human iPSC-derived liver (11), kidney (87), and lung bud tips (14) were transplanted into each of the chemically induced damaged organs. An optic-cup was also transplanted in a mouse model with retinal degeneration to induce the photoreceptors and restore the synaptic connection to recover function (88). Human iPSC-derived brain organoids have been successfully implanted into the brains of adult mice, increasing the production of mature neuronal cells and the formation of synapses with host neurons (63). Overall, using orthotopic transplantation, it is possible to study the environment during the engraftment of organoids in vivo and confirm the application prospects of organoids in cell therapy (Fig. 2).
Go to :
Future Directions for Organoids in Regenerative Medicine
Several studies have demonstrated rapid progress in organoid application, suggesting that organoids are a promising source for disease modeling, tissue engineering, and cell-based therapy in regenerative medicine. The production of mature organoids combined with challengeable applications can aid in the clinical treatment of tissues. Despite the aforementioned potential of organoids, numerous hurdles remain for achieving successful drug discovery and cell-based therapy. Ideally, protocols for differentiation of iPSC-derived organoids should be standardized for reproducibility and mass-production. Presently, iPSC-derived organoid systems are difficult to homogenize and lack scalability for high-throughput screens and large-scale cell therapy. Improving culture methods, including defined ECMs and multiscale micropatterning, may facilitate the production of more reproducible organoids. Additionally, for cell transplantation therapy, a defined ECM with excellent biocompatibility should be developed to replace Matrigel, and a scaffold that mimics real organ structure could be established. Conventional organoid differentiation protocols can generate a population of diverse cell types that occur spontaneously, but reducing their variability is a major challenge. Organoids with different characteristics are produced depending on the culture protocol, and specific mutations have been reported in patient-specific iPSC-derived organoids according to genetic background (12, 89). The introduction of gene editing technology can help generate new isogenic organoids that reduce background-related variability.
Based on the development methods of organoids mentioned in this review, organoids with stable scale-up capabilities and reproducibility should undergo three examinations before they are industrially produced as therapeutics. The first is to ensure a pathogen-free state that is directly linked to safety throughout the entire process from cell production to validation. Second, the use of advanced organoids in the industrial pharmaceutical pipeline requires the development of a storage or delivery method for organoids to optimize manufacturing costs. Finally, objective validation techniques for measuring the safety and efficacy of therapeutics in vivo should be established (90). These criteria must be met for organoids to work as drugs and some progress is already being made. Ultimately, multi-lineage organoid models can help reduce our reliance on animal models and provide new tools to treat human diseases. The various applications of the human iPSC-derived organoid systems presented here will play an important role in the future of regenerative medicine (Fig. 3).
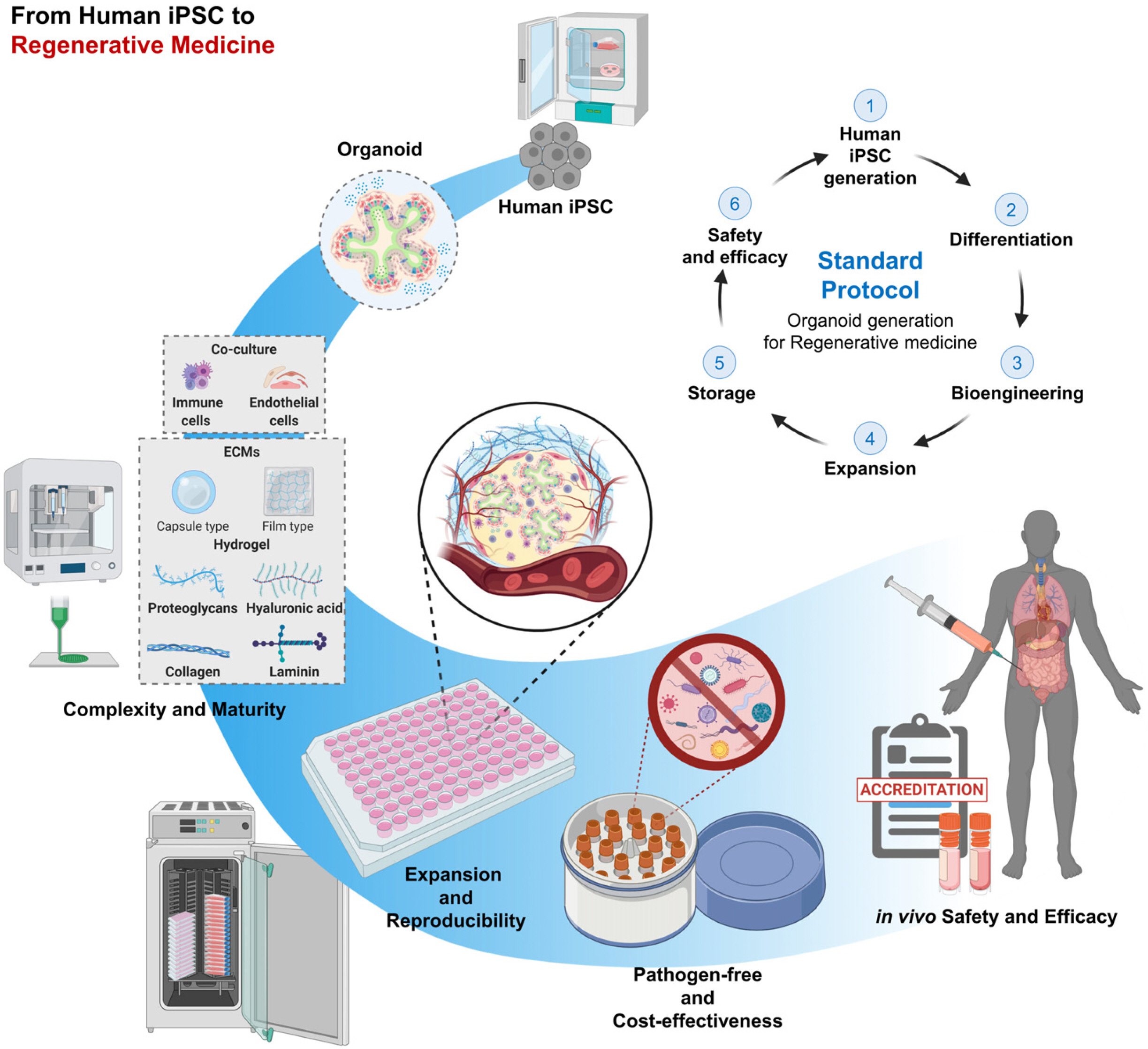 | Fig. 3Overview of the scale-up process for organoid applications covering iPSC generation to regene-rative medicine. Developing a safe and cost-effective treatment based on a standardized protocol for organoid generation and scale-up of the culture process is depicted. This figure was created using BioRender (https://biorender.com/).
|
Go to :
Conclusions
Human iPSC-derived organoids are an accessible and physiologically relevant model system that can mimic the functions of human organs without ethical concerns regarding human embryos and interspecies differences. Notably, organoids have tremendous potential in tissue biology research, disease modeling and alternative cell-based therapy. Moreover, in recent years, rapid progress has been made on the bioengineering aspects of ECM scaffolds, genome editing and organ-on-a-chip approaches to improve organoid functionality. Therefore, advanced organoid systems combined with various applications will undoubtedly expand the scope of regenerative medicine.
Go to :
Abbreviations
2D: two-dimensional; 3D: three-dimensional; ASCs: adult stem cells; BMP: bone morphogenetic protein; CRISPR: clustered regularly interspaced short palindromic repeats; ECM: extracellular matrix; ENS: enteric nervous system; FGF: fibroblast growth factor; hCOs: human colonic organoids; hESCs: human embryonic stem cells; hIOs: human intestinal organoids; hPSCs: human pluripotent stem cells; iPSCs: induced pluripotent stem cells; ISCs: intestinal stem cells
Go to :
 XML Download
XML Download