

 PDF
PDF Citation
Citation Print
Print


Introduction
Psoriasis is a chronic inflammatory skin disease with a long course and a tendency towards palindromia, and it has a great impact on the patient’s quality of life, its rates are much higher than those of cancer, myocardial infarction and congestive heart failure, and they are only second to mental diseases (1-3). While keratinocyte proliferation, lymphocyte infiltration, dermal vascular hyperplasia and dilation are the main histopathological features of psoriasis, it is also considered a systemic disease with multiple cardiovascular and metabolic comorbidities (4). Zhang et al. (5) found glucose uptake is critical for rapid keratinocyte proliferation and is an efficient response to psoriasis of physiologically relevant stressors including full-thickness wounds and UV-B irradiation.
MSCs (mesenchymal stem cells) participate in the occurrence and development of multifarious diseases and play a role in immune regulation and angiogenesis (6), therefore, they have great potential in the treatment of diseases (7). Clinical studies have shown that MSC infusion can significantly reduce the severity of psoriasis (8, 9). DMSCs are important members of the skin microenvironment, and they have gradually become the focus of dermatological researchers in recent years.
The biological activity profile of DMSCs influenced by psoriatic microenvironment remain unknown. We previously conducted a large number of studies on DMSCs in psoriasis, and the results showed that DMSCs from psoriatic patients secreted abnormal cytokines and that their DNA methylation was abnormal (10, 11). Li et al. (12) demonstrate that psoriatic DMSCs increase keratinocyte proliferation and glycolysis, and reduce cell junctions, suggesting a pathogenic role of psoriatic DMSCs in epidermal hyperplasia, aberrant differentiation, and reduction in turnover time of keratinocytes in psoriasis. How glucose metabolish influenced the DMSCs in psoriasis has not been explored. To further clarify the biologically abnormal mechanism of DMSCs in psoriasis, we detected the glucose metabolism level, cell proliferation,angiogenesis and VEC differentiation in psoriatic DMSCs.
Go to : 
Materials and Methods
Subjects
We chose 16 psoriasis patients (8 men and 8 women, ages 19∼55, PASI score 17.67±6.19) who visited our Hospital and had no systemic diseases or treatments with corticosteroids, immunosuppressive agents, or ultraviolet phototherapy within the 3 months before enrolment. The control samples (8 men and 8 women, age 25∼55) were enrolled from the Urology and Plastic Surgery Department of our Hospital.
All participants signed informed consent, and the protocol was approved by the Medical Ethics Committee of our Hospital and was in accord with the Declaration of Helsinki.
Isolation and identification of DMSCs
The specimens without adipose layers that were cut into pieces by scissors, the epidermis and dermis were separated by treatment with 0.25% dispase enzyme (Sigma Aldrich, St. Louis, MO, USA). The dermis was minced finely and turned into a single cell suspension. Cells were then cultured in DMEM/F12 (HyClone, USA)containing 10% FBS (HyClone, USA), 10 μl/ml B27 (Invitrogen, US), 100 U/ml penicillin (Sigma-Aldrich, St. Louis, MO, USA) and 100 μg/ml streptomycin (Sigma-Aldrich, St. Louis, MO, USA) at 37℃ with 5% CO2. The non-adherent cells were removed after 48 h, and the remaining cells were cultured with passaging. DMSCs were identified using flow cytometry to analyse CD29 (Backman, USA), CD44 (Backman, USA), CD105 (Backman, USA), CD34 (Backman, USA), CD45 (Backman, USA), HLA-DR (Backman, USA), Dil-LDL (Sigma-Aldrich, St. Louis, MO, USA), and UEA-1-FITC (Sigma-Aldrich, St. Louis, MO, USA). Iden-tification was also performed on differentiated adipocytes, osteoclasts and VECs. The specific methods followed published protocols (13, 14).
Cellular bioenergetic function analysis
The DMSC OCRs were measured with an XF Cell Mito Stress Test Kit (Seahorse Bioscience) (15, 16). The kit includes the following inhibitors which were oligomycin, FCCP and rotenone/antimycin-A. Oligomycin was added to evaluate ATP-linked and proton leak respiration. FCCP was added to evaluate maximal respiration and reserve capacity. Antimycin A was added to evaluate non-mitochondrial OCR (Fig. 1A).
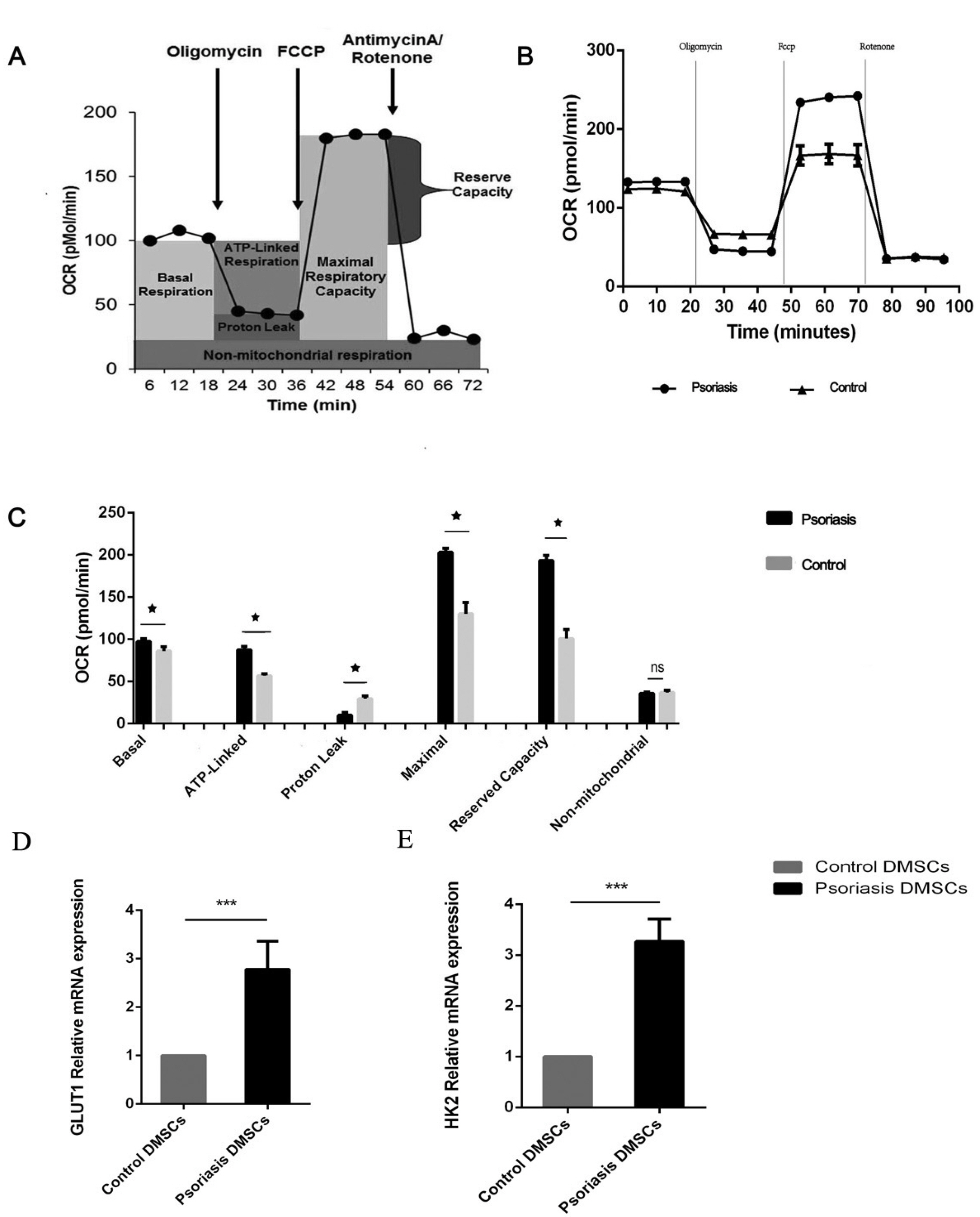 | Fig. 1The metabolic profile of DMSCs. Basal OCR was measured before oligomycin treatment, and it was subtracted after rotenone/antimycin treatment. (A) ATP-linked OCR was calculated by subtracting the basal OCR from the amount of respiration left after oligomycin was added. Proton leakage was determined by subtracting the non-mitochondrial OCR from the amount of respiration left after oligomycin was added. Maximal respiration was calculated by subtracting the maximal OCR from the non-mitochondrial OCR. The reserve capacity was determined by subtracting the maximal OCR from the basal OCR. (B) O2 consumption in the DMSCs of the psoriasis and control groups. The rates of O2 (OCR) were first measured in DMSCs under basal conditions and then sequentially after the addition of oligomycin (1 µM), carbonylcyanidep-(trifluoromethoxy), phenylhydrazone (FCCP) (2 μM), and rotenone (0.5 μM). (C) Graphs show the ATP-linked OCR, proton leak OCR, maximal OCR, reserve capacity and non-mitochondrial OCR in the psoriasis and control groups. (D, E) Histograms showing the relative mRNA expression levels of GLUT1 and HK2. ns p>0.05, *p< 0.05, ***p<0.001.
|
Analysis of DMSC proliferation
The isolated DMSCs were diluted with DMEM/F12 medium and then inoculated into the 24-hole culture plate at a density of 2×105/ml to be maintained at 37℃ and 5% CO2. Half of the medium was replaced every third day. DMSCs were stained with trypan blue and counted every 3 days using an Invitrogen cell counter (Countess II Automated Cell Counter, Thermo Fisher Scientific).
Blood vessel counts and pathology
Standard HE staining procedures were followed. First, samples were preserved in 4% formaldehyde, followed by dehydration, embedding, impregnation, staining and sealing. An inverted phase contrast microscope (Olympus- IMT2) was used for microscopic observation. Five visual fields were randomly selected from each section, blood vessels were identified, and the mean number of blood vessels was calculated.
RNA extraction and qRT-PCR
Total RNA from DMSCs was extracted using Trizol (Invitrogen, USA), RNA purity was determined (Biodrop, CO, UK), and cDNA was synthesized (Takara, Shiga, Japan). cDNA was synthesized using 5X PrimeScript RT Master Mix (2 μl) and total RNA and RNase Free dH2O in a total volume of 10 μl (TaKaRa, Shiga, Japan). PCR amplification was performed in a 20 μl reaction system containing 2 μl template, 10 μl TB Green Premix Ex Taq II (TaKaRa), 0.4 μl Rox Reference Dye, 6 μl sterile purified water, and 0.8 μl of PCR primers. The primer sequences are shown in Table 1. Initial denaturation was performed at 95℃ for 30 s and was followed by 40 cycles of denaturation at 95℃ for 10 s, annealing at 60℃ for 30 s, dissociation at 95℃ for 15 s, 60℃ for 1 min, and 95℃ for 15 s. PCR products were identified using 2% agarose gel electrophoresis.
Table 1
The information of primers
| Targets | Primer sequence (5’-3’) | Annealing temperature (℃) | Function | Referenced website of function |
|---|---|---|---|---|
| JUNB | Sense: ACGACTCATACACAGCTACGG | 58 | Basic cellular activities | https://www.ncbi.nlm.nih.gov/gene/3726 |
| Antisense: GCTCGGTTTCAGGAGTTTGTAGT | ||||
| FOS | Sense: GGGGCAAGGTGGAACAGTTAT | 58 | Cell proliferation | https://www.ncbi.nlm.nih.gov/gene/2353 |
| Antisense: CCGCTTGGAGTGTATCAGTCA | ||||
| CXCR7 | Sense: TCTGCATCTCTTCGACTACTCA | 60 | Angiogenic | https://www.ncbi.nlm.nih.gov/gene/3726 |
| Antisense: GTAGAGCAGGACGCTTTTGTT | ||||
| CXCL12 | Sense: ATTCTCAACACTCCAAACTGTGC | 56 | Angiogenic | https://www.ncbi.nlm.nih.gov/gene/6387 |
| Antisense: ACTTTAGCTTCGGGTCAATGC | ||||
| RGS5 | Sense: GACATGGCCCAGAAAAGAATCC | 58 | Angiogenic | https://www.ncbi.nlm.nih.gov/gene/8490 |
| Antisense: CACAAAGCGAGGCAGAGAATC | ||||
| HEYL | Sense: GGCTGCTTACGTGGCTGTT | 58 | Differentiation | https://www.ncbi.nlm.nih.gov/gene/26508 |
| Antisense: GACCCAGGAGTGGTAGAGCAT | ||||
| Notch3 | Sense: TGGCGACCTCACTTACGACT | 58 | Differentiation | https://www.ncbi.nlm.nih.gov/gene/4854 |
| Antisense: CACTGGCAGTTATAGGTGTTGAC | ||||
| Glut1 | Sense: CTGGCATCAACGCTGTCTTC | 60 | Glucose transporter | https://www.ncbi.nlm.nih.gov/gene/38109 |
| Antisense: GCCTATGAGGTGCAGGGTC | ||||
| Hk2 | Sense: GAGCCACCACTCACCCTACT | 60 | Glycolysis | https://www.ncbi.nlm.nih.gov/gene/3099 |
| Antisense: CCAGGCATTCGGCAATGTG |
![]()
Statistical analysis
SPSS 18.0 software was used to perform statistical analysis. The number of blood vessels and the number of cells (from proliferation) were analysed by Student’s t-test, the dates expressed as mean±SD and p<0.05 was considered to indicate statistical significance. The mRNA expression was analysed using the 2−ΔΔCt method.
Go to :
Results
Identification of DMSCs
The cell morphology of DMSCs was fibroblast-like in appearance with no distinct morphological differences among groups (Fig. 2A∼C). All cells showed high expression levels of CD29 (94.6%), CD44 (92.8%) and CD105 (95.0%) but low expression of CD34 (0.6%), CD45 (0.2%) and HLA-DR (1.2%), confirming the MSC phenotype (Fig. 2G). By culturing in specified induction medium, the cells exhibited the ability to differentiate into adipocytes (Fig 2D), osteoclasts (Fig 2E) and VECs (Fig 2F), confirming the multidirectional differentiation ability of MSCs. Psoriatic DMSCs have stronger differentiation function compared to normal DMSCs, the result has been reported in our previous study (17).
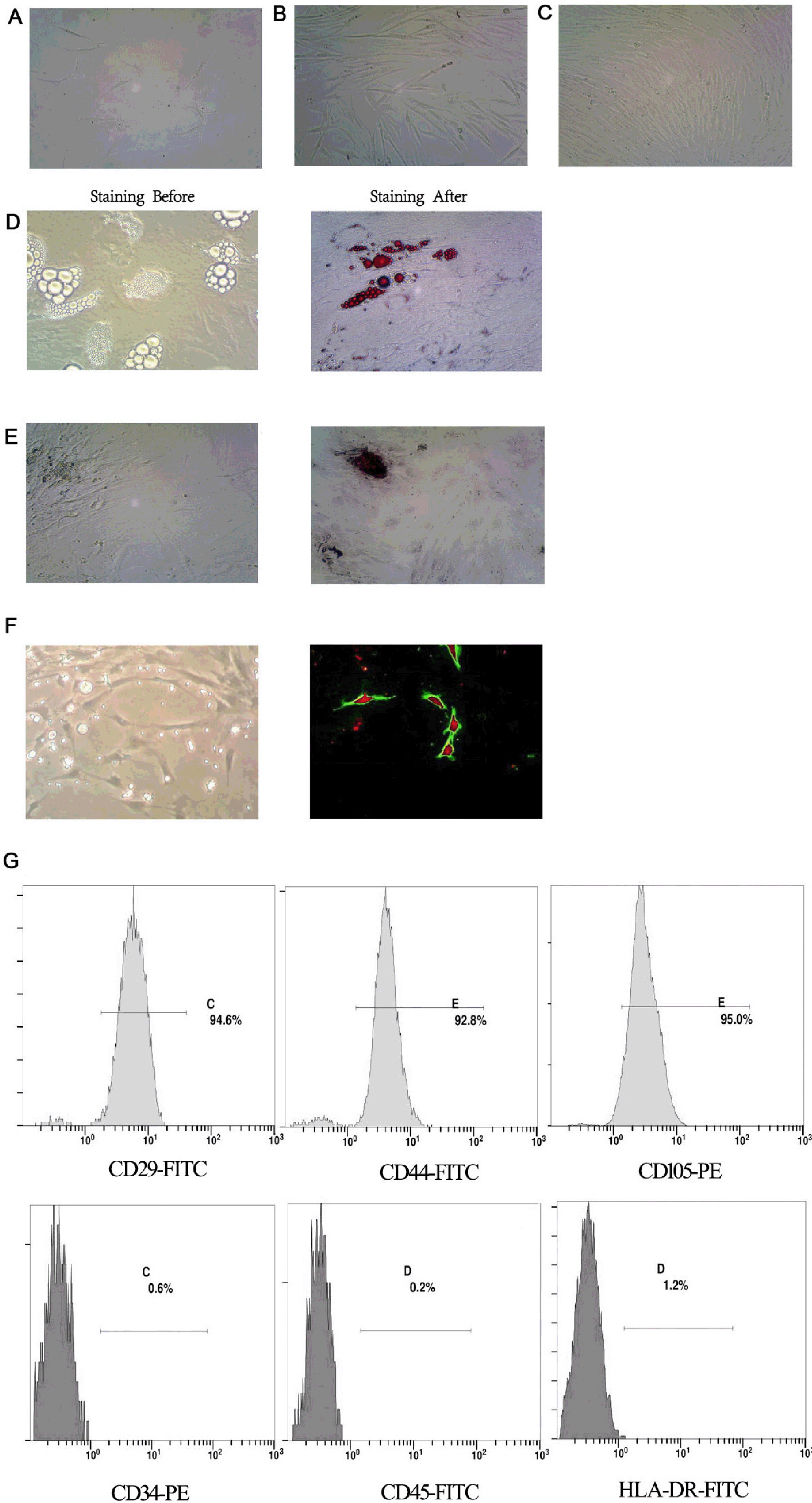 | Fig. 2Identification of DMSCs. (A∼C) Morphological characteristics of DMSCs cultured for 6, 12 and 15 days (×200). (D, E) Morphology of adipocytes and osteoblasts generated by the induction of DMSCs; adipocytes stained before; adipocytes stained with oil red O; osteoblasts stained before; osteoblasts stained with alizarin red (×400). (F) Angiogenic network in DMSCs after differentiation (×400). Overlay of ac-LDL and UEA-1 in DMSCs after differentiation, the uptake of ac-LDL stimulated red fluorescence, the UEA-1stimulated green fluorescence, and the ac-LDL and UEA-1stimulated red and green fluorescence (×200). (G) Identification of DMSCs by flow cytometry.
|
The glucose metabolic profile of DMSCs
The overall OCR profile was higher in the psoriatic DMSCs compared to those from healthy subjects (Fig. 1B). The basal respiration (p<0.05), ATP-linked respiration (p<0.05) and maximal respiration (p<0.05) were significantly higher in the psoriatic DMSCs compared to the control DMSCs, but they had decreased proton leak respiration. Moreover, the reserve capacity respiration (p< 0.05) was also significantly higher in the psoriatic DMSCs, but non-mitochondrial respiration had no statistical significance (Fig. 1C). The results indicated the increased glucose levels and mitochondrial metabolism supported cellular activity.
Glucose metabolism, especially when ATP-linked, is essential for cellular activity. Glucose metabolism is linked to cell proliferation and differentiation. The link is present in many cells ranging from microorganism to T cells, but there is no report of such a link in DMSCs, particularly in psoriatic DMSCs.
Expression of mRNA related to glucose metabolic in DMSCs
Glut’s (glucose transporters) regulate the availability of glucose for most tissues. Recent studies reveal that GLUT1 which is a member of facilitative sugar transporters, up regulation is considered to promote epidermal hyperproliferation, inflammation and angiogenesis in the pathogenesis of psoriasis (18). Hexokinases (HKs), which play a key role in the first step of glucose metabolism (19). In order to clarify the mRNA differences of glucose metabolism between psoriasis patients and normal control derived DMSCs, we detected the expression levels of GLUT1 and HK2. GLUT1 (2.78±058-fold) and HK2 (3.27±0.44-fold) were up-regulated in the psoriatic DMSCs (Fig 1D and 1E).
Proliferation of DMSCs
The growth rates of DMSCs were estimated by cell count for each group, and the values are shown in Fig. 3A. On day 3, the cell number in DMSCs from the psoriasis group was lower (3.76±0.32) than that in DMSCs from the control group (4.0±0.18), p<0.05 (Fig 3B). However, after long-term cultures (15 days), the cell proliferation rate of psoriatic DMSCs was similar to that of the control cells (p>0.05).
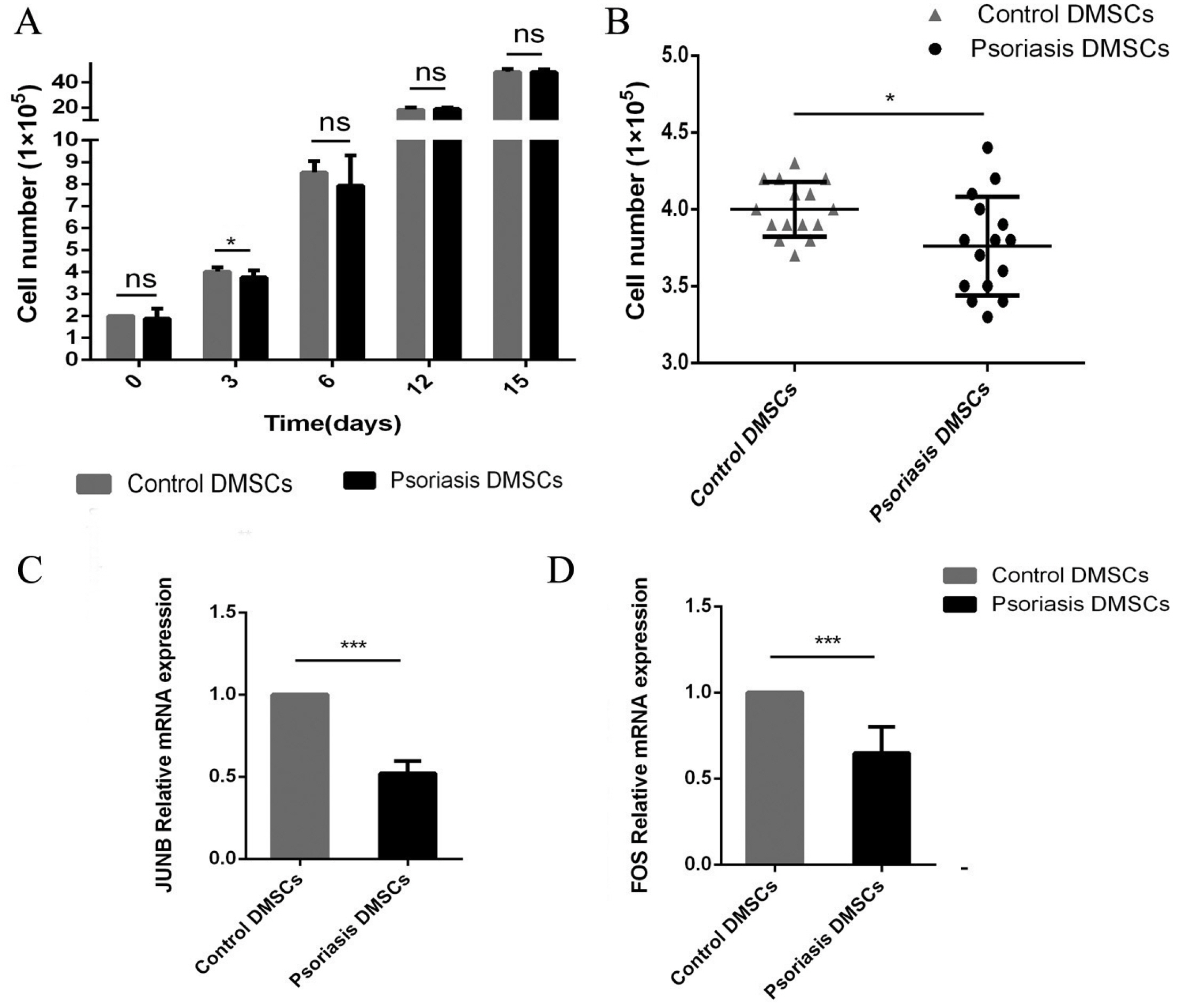
Expression of mRNA related to cell proliferation in DMSCs
To investigate the different molecular mechanisms of cell proliferation after 3 days, we evaluated the mRNA expression of genes related to cell proliferation, including JUNB and FOS (Fig 3C and 3D). The mRNA expression of JUNB in psoriatic DMSCs was (0.52±0.08) times what was observed in normal DMSCs. Similarly, the mRNA expression of FOS was downregulated in psoriatic DMSCs to (0.65±0.15) times the levels seen in the control.
Number of dermal vessels in the two groups
To investigate the link between glucose metabolism, angiogenesis and VEC differentiation, we examined the blood vessels in psoriatic lesions. After staining, dermal vessels were counted under the microscope. The results showed that the number of dermal vessels in psoriatic lesions and controls were (42.2±4.13) and (18.2±3.00) respectively. The number of blood vessels in psoriatic lesions was significantly higher than that in the control group (p<0.001). The results are shown in Fig. 4A∼E.
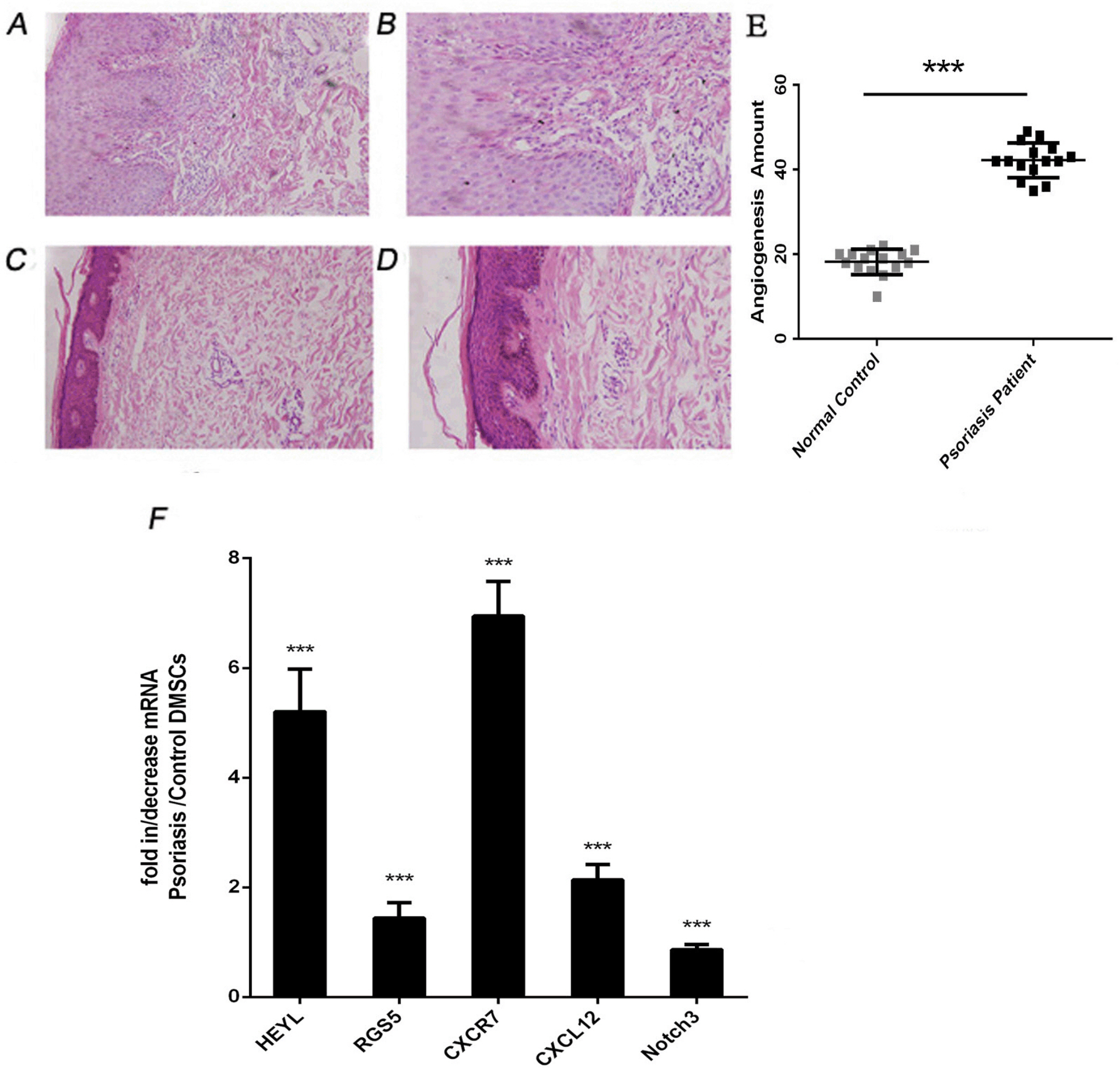 | Fig. 4The number of local blood vessels and the expression of mRNA related to VEC differentiation in the psoriasis and control groups. (A∼D) Histopathology skin tissue by haematoxylin and eosin (HE) staining as seen under a microscope; A&B were psoriatic lesions; C&D were control skin tissue (A&C ×200 and B&D ×400, respectively). (E) Blood vessel counts between the psoriasis and control groups. (F) HEYL, CXCR7, CXCL12, RGS5 and Notch3 mRNA relative expression. ***p<0.001.
|
Expression of mRNA related to angiogenesis and VEC differentiation
Our previous research showed that the ability of psoriatic DMSCs to differentiate into VECs in vitro was enhanced compared to normal DMSCs (17, 20). To analyse the molecular mechanism of abnormity of local psoriatic blood vessels, the mRNA expression of five mRNAs associated with angiogenesis, VEC differentiation and major factor of signaling pathways was examined with real-time RT-PCR. The analysed mRNAs were CXCR7, CXCL12, RGS5, HEYL, and Notch3. Foure of the five mRNAs were upregulated in psoriatic DMSCs which HEYL (5.20± 0.78-fold), RGS5 (1.44±0.28-fold), CXCR7 (6.95±0.63-fold) and CXCL12 (2.13±0.29-fold). The mRNA expression of Notch3 was downregulated in psoriatic DMSCs to (0.86± 0.09-fold) the levels seen in the control. The results are shown in Fig 4F.
Go to :
Discussion
Glucose metabolism is necessary for basic cellular activities and an important metabolic pathway by regulating ATP, nucleotides, and amino acids. It has obvious changes in the development of diseases, and is an important biological indicator (15). Brownstein (21) first suggested that psoriasis may be associated with type 2 diabetes mellitus in 1966 and since then most studies is accumulating suggesting that glucose metabolism in the pathogenesis of psoriatic (5, 22). But the results of N. U. Friis were widely conflicting with the studies (16). Therefore, it is necessary to study the metabolic status of DMSCs in patients with psoriasis. We detected the OCR of DMSCs with the Extracellular Flux Analyzer XFe24. The results showed that the OCR of DMSCs from psoriasis patients was significantly higher than control group. Basal respiration, ATP-linked respiration, maximal respiration and reserve capacity were also significantly higher in the psoriatic DMSCs. Choi et al. (23) reported that ablation of Glut1 attenuates animal models of psoriasis. HK2 is critically important for aerobic glycolysis (24). We resulted that GLUT1 and HK2 were up-regulated in the psoriatic DMSCs. Upregulation of glucose metabolism is considered to promote hyperproliferation, inflammation and angiogenesis in the pathogenesis of psoriasis. To examine the link between glucose metabolism with DMSC proliferation and local psoriatic blood vessels, we compared the DMSCs of proliferation and angiogenesis derived from psoriatic patient and normal control.
Cell proliferation is an important feature of life and controlled by many factors. FOS are primary oncogenes and subunits of the JUNB (AP-1) transcription factor. FOS combines with JUNB to perform specific cellular functions (25). JUNB participates in multiple physiological processes of cells, including proliferation (26, 27). Fan et al. (28) confirmed that knockdown of JUNB significantly inhibited in MM (multiple myeloma) cell proliferation and survival. Martins GA showed that Fos is a component of JUNB, a strong activator of IL-2, which associated with the cell proliferation and survival (29). He et al. (30) suggest that c-fos transcription via the Ras/ ERK1/2 pathway to promote the proliferation of (sper-matogonial stem cells) SSC. Glucose is a preferred bioenergetic and synthetic substrate for rapidly proliferating cells, but our results were that the expression of JUNB and FOS was lower in psoriatic DMSCs, corresponding that the cell number on the third day. We preliminarily concluded that FOS and JUNB were involved in the early proliferation regulation of DMSCs in psoriasis.
The typical clinical feature of psoriasis is a squamous erythema or plaque, and vascular hyperplasia is obvious in psoriasis lesions. Vascular abnormalities are affected by many factors, but the exact mechanism remains unclear. Abnormal glucose has been found in many diseases including psoriasis. Our study showed that the number of blood vessels in psoriasis lesions was significantly increased compared to the control group, consistenting with the previous experimental results which psoriatic DMSCs have stronger differentiation function compared to normal DMSCs (17, 31). In addition, we studied the mRNA expression levels associated with angiogenesis and differen-tiation. CXCL12, known as SDF1 (chemokine stromal- derived factor-1). The Virgintino et al. (32) supported that in human brain, CXCL12/CXCR7 system appears to be directly involved in microvessel growth, its members being differentially expressed in angiogenically activated microvessels and vascular sprouts. Weng et al. (33) showed that CXCR4/SDF 1 axis was involvement the effects of high glucose on proliferation and function of circulating fibrocytes. RGS5 (regulator of G protein signalling 5) is correlated with vascular abnormalities (34). A study confirmed that consistently inhibiting the expression of Notch3, HeyL and Rgs5, affecting the tumor vasculature (35). Notch3 is an important factor in the NOTCH signalling pathway, the NOTCH signalling pathway is an important pathwy related to differentiation, vascular hyperplasia and glucose metabolism (36). HEYL, a primary target genes of Notch, cause congenital heart defects (37). The level of mRNAs showed that CXCL12, CXCR7, HEYL and RGS5 were upregulated in psoriatic DMSCs, in addition to Notch3.
Angiogenic switch has long been considered to be dictated by angiogenic growth factors (eg, vascular endothelial growth factor) and other signals (eg, Notch) alone, but recent findings show that it is also driven by a metabolic switch (38). Vigo et al. (39) pointed out that modifications of MSC energetic metabolism supported a role for metabolic pathways in the therapeutic function of MSC. Zhang Hui, who showed that hPSC (Human Pluripotent Stem Cells) differentiation was accompanied by increased glycolysis and oxidation metabolism levels (40). A number of studies have shown that proliferation, angiogenesis were simultaneously affected by metabolism.
Psoriatic lesions are a complex three-dimensional cell network composed of many cell types. We speculated that abnormal glucose metabolism level of DMSCs affected their own capacity for proliferation, vascular differentiation and the local vascular abnormalities in patients with psoriasis, which resulted in the pathophysiological process of psoriasis. The studies valuable for understanding the pathogenesis of psoriasis.
Go to :
 XML Download
XML Download