

 PDF
PDF Citation
Citation Print
Print


Introduction
Ocular toxicity tests are required to evaluate risks and ensure the safety of ophthalmic administration of drugs (1, 2). Several in vivo animal models have been used in this sense. However, species-related morphological and biochemical differences to the human eye compose major limitations of such models (3, 4). Furthermore, animal experiments have been extensively criticized in terms of cost, time and ethical issues (5). Thus, the search for alternative models for animal experiments has been encouraged for several fields, including ophthalmology. Cell culture models derived from human resources offer the advantage of constituting highly defined systems and may result in more reproducible data (6-9).
The retinal pigmented epithelium (RPE) is a monolayer of pigmented epithelial cells that reside between the neural retina and Bruch’s membrane (BM). Even though RPE is not an intrinsic component of the visual signaling pathway, it is a highly metabolically active cell layer, which is vital to the health, survival, and function of the overlying photoreceptors (10, 11). Considering that RPE is critically important for normal function of the retina, intraocular drug or compound administration must be evaluated regarding possible toxicity against this cell layer (1, 12).
ARPE-19 was established and characterized in 1996 (6). Despite being considered a representative RPE cell line, these cells display poor transepithelial resistance values of ∼100 Ω.m2 and seem to lose RPE-specific genes when maintained in suboptimal culturing conditions (13). These limitations have encouraged the search for protocols for in vitro RPE generation from human pluripotent stem cells (hES-RPE) (14). hES-RPE closely resembles human fetal RPE and were capable of phagocytosis of fluorescently labeled rod outer segments. Therefore, hES-RPE have been investigated for cellular therapy, disease modeling and drug screening (2, 10, 11, 14-16).
Several diseases that cause ocular inflammation, including uveitis, scleritis, and orbital inflammatory disease result in impairment or loss of vision (17). The mainstay treatment is the use of corticosteroids, but the prolonged treatments and high doses of these drugs are associated with significant side effects (18). For this reason, corticosteroid-sparing agents like Cyclosporin (CSA) (19), Sirolimus (SRL) (20), Tacrolimus (TAC) (21), Leflunomide (LEF) (22) and its active metabolite teriflunomide (TER) have been investigated as alternatives to the use of corticosteroids.
While CSA, SRL, and TAC have already been applied for ocular diseases, there are few studies investigating LEF for this purpose (22-24). Nevertheless, the effect of these chemicals on RPE has not been verified in vitro, since the technology for producing functional hES-RPE is quite recent. Considering that, here, we aim to evaluate the differences between the RPE cell line ARPE-19 and hES-RPE cells, in order to underscore the importance of determining the best RPE source for preclinical toxicity drug testing. To do so, both RPE cell sources were characterized molecularly and functionally and used to assess the toxicity of the aforementioned drugs.
Materials and Methods
Cell culture
H1 (25) (National Institutes of Health–registered as WA01) were maintained in Matrigel (BD Biosciences, USA) using mTeSR (StemCell Technologies, USA) and subcultured using Dispase (BD Biosciences, USA). Differentiation protocol was performed by allowing H1 to overgrow until the hES colonies became multilayered. Culture media was then replaced with RPE differentiation medium (RPE medium), composed of knockout high glucose DMEM supplemented with 0.1 mg/ml Normocin (Invivogen, USA), 1% nonessential amino acids solution, 2 mM GlutaMAX-I (Invitrogen, USA), 0.1 mM mercaptoethanol (Invitrogen, USA), 13% Serum Replacement (Invitrogen, USA) and 5% Fetal Bovine Serum (FBS) (Cripion Biotecnologia LTDA, BRA) (Fig. 1A).
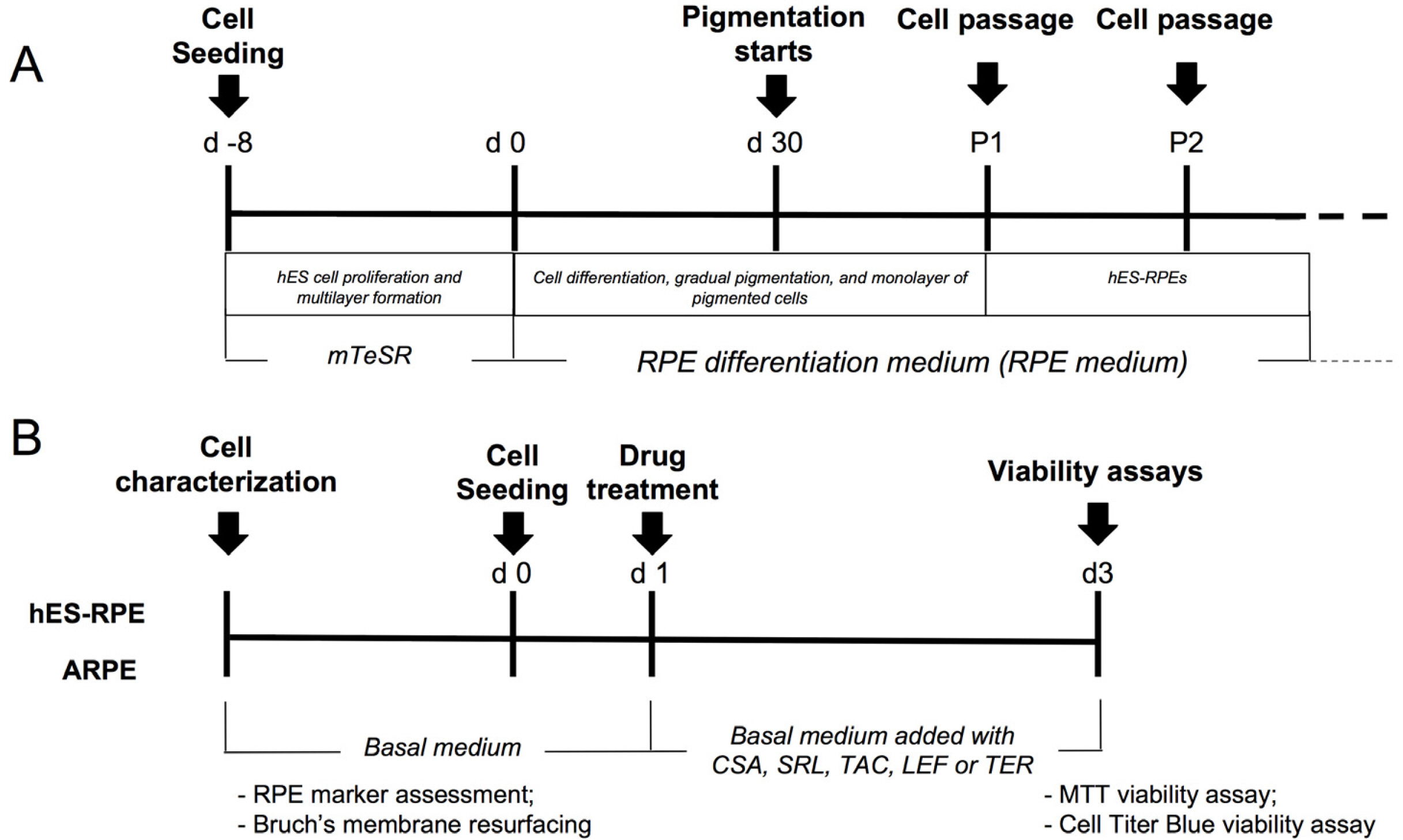
Fig. 1
hES-RPE differentiation and experimental design. (A) hES-RPE were differentiated by proliferation and multilayer formation of hES cells and removal of bFGF of the medium. (B) hES-RPE and ARPE-19 were first characterized according to mRNA expression of RPE markers, then according to Bruch’s membrane resurface ability. Finally, hES-RPE and ARPE-19 were used for cytotoxicity assays.
![]()
ARPE-19 cell line, previously described as a human RPE cell line, was cultured in DMEM/F12 supplemented with 10% Fetal Bovine Serum. The medium was changed every 2 days and cells were used between passages 9-19.
Primary RPE isolation
Primary RPE cells were isolated from fetal (18∼22 weeks gestation) and adult (59∼63 years old) eyes (Advanced Biosciences Resources, Inc, USA). Cells were obtained following RPE layer digestion using collagenase IV (Gibco, USA) at 0.8 mg/ml (fetal eyes) or 0.4 mg/ml (adult eyes). RPE cells were cultured using RPE medium and tissue culture flasks covered with the extracellular matrix produced by bovine corneal endothelial cells (26). After reaching confluence, cultures were expanded using 0.25% trypsin-EDTA.
Quantitative PCR
Cells were harvested from fetal and adult RPE (fRPE and ARPE, respectively), ARPE-19 and hES-RPE cultures for mRNA characterization (Fig. 1B). fRPE were collected at Passage 1, day 4 for experiments. ARPE were dissected from a donor eye (age 59∼63) and directly processed for RNA isolation using Trizol. Total RNA samples were treated with DNase (Promega, USA) and quantified by spectrophotometry. cDNA was obtained using the RevertAidTM H Minus M-MuLV RT (Fermentas, USA). Next, PCR amplification for pluripotent stem cell markers OCT-4 and NANOG, and RPE markers RPE-65, bestrophin, CRLBP, MITF, PEDF, and ZO-1, was performed using the TaqMan Gene Expression Assay kit (Applied Byosystems, USA). The relative level of gene expression was determined and normalized to 18s rRNA. Each sample was run in technical triplicates and biological dup-licates. Comparisons were made considering samples processed at the same time.
Western blotting
The protein expression of MITF and BEST was also evaluated using Western Blotting. Cell lysates were prepared from RPE-hESC in passages 1 to 3. The cells were lysed on ice using cold lysis buffer (20 mM Tris, 1 mM EDTA, 150 mM NaCl, 1% Triton X-100, 10% glycerol) containing protease inhibitor (Protease Inhibitor cocktail II, Calbiochem). Then, samples were vortexed for 15 seconds and incubated on ice for 5 min. After repeating this step two more times, the lysate was centrifuged for 15 min at 13500 g at 4℃. Protein concentration was measured by Bradford assay (BioRad). Cell lysates (30 μg) were mixed with Laemmli buffer, added with β-mercaptoethanol and boiled for 5 min. Proteins were separated using 12% SDS-PAGE polyacrylamide gel and then transferred to a nitrocellulose membrane for 2 h at 75V in transfer buffer (25 mM Tris, 192 mM glycine, 20% methanol, pH 8.3). The membrane was washed with PBS-T (Tween-20 0.05%) and blocked with 5% BSA in PBS-T for 1 h. The membranes were incubated with mouse anti-RPE-65 1:500 (Pierce) and mouse anti-CRLBP 1:750 (Abcam) overnight at 4℃. After 3 washes of 15 min in PBS-T, the membranes were incubated with secondary antibody anti-mouse IgG diluted 1:10,000. Chemiluminescence was developed using the ECL Plus kit (Amersham Biosciences) and the signal scanned and collected using a Typhoon imager (Amersham Biosciences).
Immunofluorescence
RPE markers were also analyzed in hESC-RPE using immunofluorescence. For this, passage 2 RPE cells were fixed with 4% paraformaldehyde for 15 min, permea-bilized with 0.1% Triton X-100, and blocked with a solution containing 2% goat serum (Normal Goat Serum - NGS) 0.5% BSA, and 0.1% Triton X-100 diluted in PBS for 1 hour. Primary antibodies were diluted in 2% NGS / PBS-T 0.3% and incubated at 4℃ overnight, using the following dilutions: 1:50 of the rabbit polyclonal transcription factor associated with microphthalmia, MITF (Abcam); 1:250 of mouse monoclonal bestrofin membrane protein, BEST (Pierce). Goat anti-mouse IgG FITC (1:32; Sigma), and goat anti-rabbit IgG rhodamine (Jackson ImmunoResearch Laboratories) were diluted in blocking solution and incubated with samples for 1 hour at room temperature. Nuclear staining was executed using 0.2 μg/ml DAPI (Sigma) diluted in PBS. The slides were assembled using Vectashield (Vector Laboratories). The images were obtained using a confocal microscope (Confocal Zeiss 5 LIVE).
Melanin content analysis
Melanin content is one of the criteria used to select and characterize hESC-RPE batches used for cellular therapy (27-29). In order to quantify the intracellular melanin content of hESC-RPE, passage 2 cells were harvested on days 8, 12 and 16 after passage. The cells were centrifuged at 160 g for 5 min at room temperature and counted. Pellets were resuspended in 1M NaOH and heated at 80℃ for 10 min, vortexed, and the absorbance measured at 475 nm against a standard synthetic melanin curve (Sigma) ranging from 5 to 180 μg/ml. Samples were analyzed in triplicates and the data normalized to the total number of cells.
VEGF synthesis
hESC-RPE were plated at 6×104 cells/cm2 on Transwell plates (Corning) prepared with Matrigel or BM substrates (Sigma). The cell culture supernatants on the apical and basal sides (the compartment above and below the Transwell, respectively), were collected 48 hours after plating and the samples assayed, in duplicates, with the DuoSet ELISA Human VEGF R&D Systems kit.
Explant assay - Bruch’s membrane organ culture
The ability of ARPE-19 and hES-RPE cells to resurface aged human BM was assessed since this is the area where RPE is first affected in diseases like AMD, and also as an additional evidence of the resemblance of ARPE-19 and hES-RPE to the native RPE. In this assay, cellular attachment and survival were analyzed by scanning electron microscopy (SEM).
In order to prepare the ex vivo culture experiment, adult donor eyes (age 59∼63) were received from the eye bank of the city of Belo Horizonte, MG - Brazil (Ethical Committee approval no. ETIC 33734514.7.0000.5149). Acceptance criteria followed previous studies (30, 31). Six-millimeter-diameter corneal trephines (Bausch and Lomb, USA) was used to create macula-centered BM explants which were debrided. Explants were seeded with 3,164 cells/mm2, shown to yield a monolayer of cells with 24 h after seeding (30). Explants were harvested at day 7, fixed in paraformaldehyde-glutaraldehyde Karnovsky’s Fixative solution and processed for SEM.
Scanning electron microscopy: Explants were fixed as mentioned and washed 3 times with PBS and incubated with 1% osmium tetroxide (Electron Microscopy Sciences, USA) in PBS at 4℃ in the dark for 1 h. After, samples were washed with PBS and incubated in 1% tannic acid in water. Explants were subsequently washed with PSB and incubated in 1% osmium tetroxide in water at 4℃ in the dark for 1 h. Then, samples were washed with PBS and dehydrated through stepwise incubation in a series of graded ethanol baths at 35%, 50%, 70%, 85%, 95% and 100% concentration. Samples were submitted to critical point drying using CO2 (Leica EM CPD030, USA). Finally, explants were coated with gold (BALTEC MED020 Coating System, LIE) and analyzed in the Microscopy Center of Biological Sciences Institute, Federal University of Minas Gerais, Brazil (DSM950- Zeiss, GER).
Drug toxicity testing
hES-RPE cells and ARPE-19 were seeded on 96-well plates at 1×104 cells/well. After 24 h, the medium was changed and CSA, SRL, TAC, LEF, and TER were added at increasing doses of 31.6 μM (101.5), 56 μM (101.75), 100 μM (102), 177.8 μM (102.25) and 316 μM (102.5). Cells were incubated with respective drugs for 72h and had their viability assessed by the 3-(4,5-dimethylthiazol-2-yl)- 2,5-diphenyl tetrazolium bromide (MTT) assay (32) or CellTiter Blue, following manufacturer’s instructions. Data were obtained from three independent experiments. The IC50 (concentration of the drug in which cell viability decreased by 50%) was determined for each drug incubated with each cell.
Results
Analysis of RPE marker expression in hES-RPE and ARPE-19
hES-RPE and ARPE-19 are considered relevant experimental models of the native RPE, for in vitro investigations. In order to assess the similarity of hES- RPE and ARPE-19 relative to fRPE and in situ ARPE, these cells were obtained and characterized morphologically, molecularly and functionally (Fig. 1B, Supplementary Fig. S1). mRNA expression of proteins involved in visual cycle (CRLBP and RPE-65), RPE-specific transcription factors (MITF), membrane-associated proteins (BEST), secreted factors (PEDF) and tight junction proteins (ZO-1) was performed. Obtained data showed that hES-RPE mRNA levels were similar to fRPE and ARPE (Fig. 2A). On the other hand, ARPE-19 revealed lower expression levels of all RPE markers compared to other groups, with ZO-1 being the only exception, presenting statistically similar expression between ARPE and ARPE-19. Despite constituting an important gene for RPE function, due to its contribution to tight junction formation and integrity of the blood-retina barrier, ZO-1 is not a specific RPE marker. Compared to undifferentiated pluripotent stem cells, hESC-RPE presented lower expression of pluripotency markers, as well as higher expression of RPE markers (Fig. 2B).

Fig. 2
mRNA and protein expression of RPE markers. (A) The mRNA expression of RPE-65, BEST, CRLBP, MITF, PEDF, and ZO-1 was evaluated by quantitative real-time polymerase chain reaction. mRNAs were quantified from hES-RPE, ARPE-19, fetal RPE, and adult RPE normalized to the geometric mean of a housekeeping gene (18s rRNA). (B) The mRNA expression of pluripotency (OCT4 and NANOG), neuroectoderm (PAX6) and RPE markers (MITF, CRLBP, BEST, RPE-65, PEDF and ZO-1) was also compared between undifferentiated hESCs and hESC-RPE. As expected, compared to undifferentiated H1, differentiated cells at passage 2 presented lower expression of pluripotency genes, and higher expression of neuroectoderm and RPE differentiation markers. (C) Protein expression of BEST and MITF was evaluated in p2 hESC-RPE cells by immunofluorescence. As expected, BEST presented membrane localization, and MITF, nuclear localization. (D) Western Blotting analysis of RPE-65 and CRLBP expression by p1, p2 and p3 hESC-RPE cells. Similar to fetal RPE, hESC-RPE cells from all the analysed passages presented detectable expression of both RPE markers. Bars represent standard error of the mean. Abbreviations: hES-RPE, RPE derived from human embryonic stem cells; fRPE, fetal RPE.
![]()
The RPE differentiation was also investigated using protein markers. Immunofluorescence analysis confirmed that hESC-RPE cells expressed BEST, MITF, with membrane and nuclear localization, respectively (Fig. 2C).
Finally, the hESC-RPE differentiation was confirmed using western blotting analysis of RPE-65 and CRLBP, which were expressed by hESC-RPE in different passages, similar to fRPE cells (Fig. 2D).
Pigment production is also a determining factor in the maturation stage of cells, this being one of the criteria used to determine the degree of differentiation that cells should be used for therapy. Therefore, the functionality of hESC-RPE was analyzed according to the melanin content of cells in different time-points of differentiation (Fig. 3A), and revealed that the differentiated cells accumulated intracellular melanin over time. The verified melanin content surpassed the minimal hESC-RPE melanin content for clinical application (27-29).
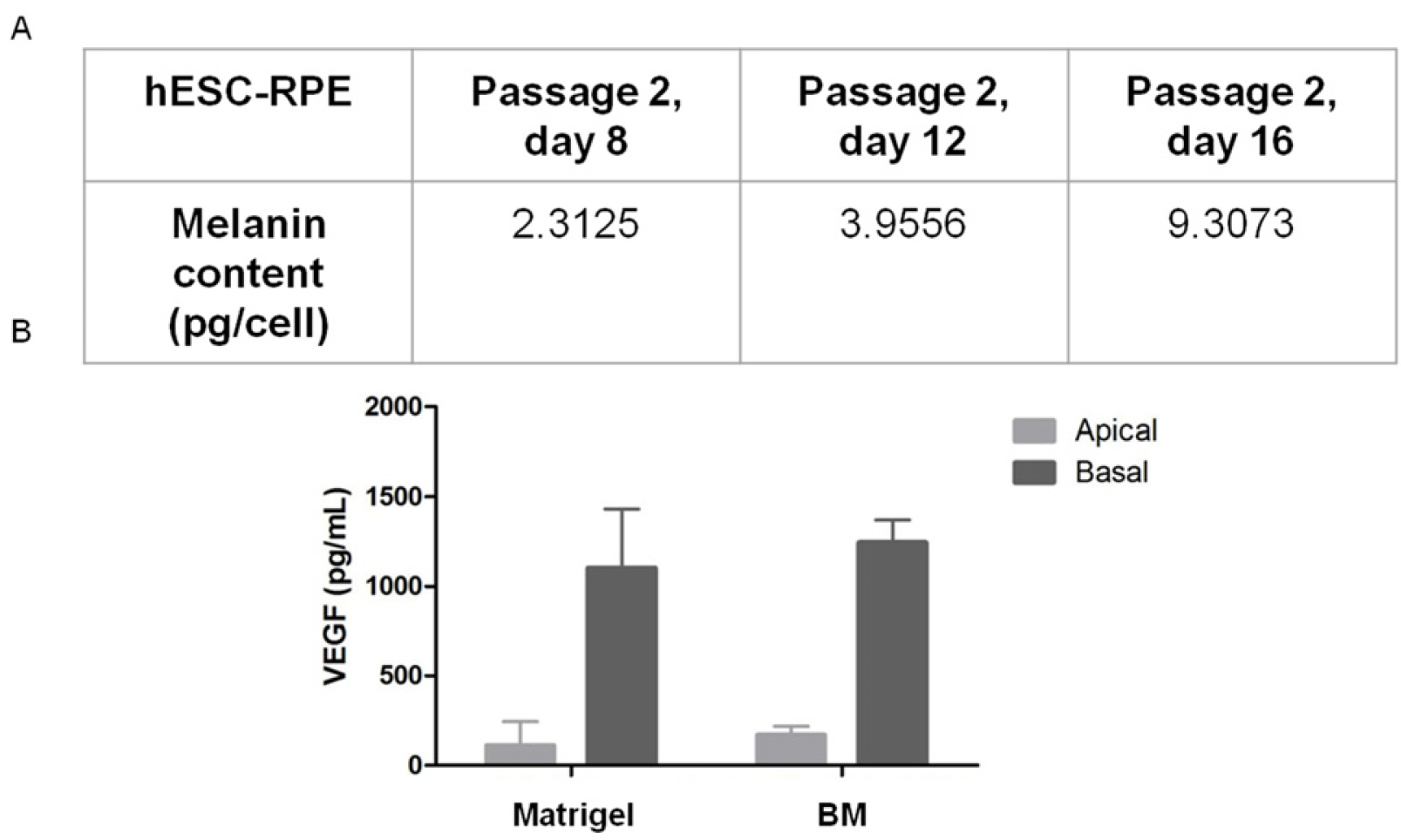
Fig. 3
Functional analysis of hESC-RPE cells. (A) Melanin content of passage 2 hESC-RPE cells in different times of culture. (B) Polarized VEGF synthesis of hESC-RPE cells. As depicted, hESC-RPE cells preferentially secrete VEGF in the basal side. Bars represent standard error of the mean. Abbreviations: hES-RPE, RPE derived from human embryonic stem cells; BM, human Bruch’s membrane.
![]()
RPE cells in situ are known to secrete growth factors, such as VEGF, in a polarized manner (10). Similarly, RPE monolayers cultured in vitro show preferential secretion of VEGF in the basal side (33). In order to verify the functional character of the EPR-hESC cells, they were seeded in transwells and the culture medium was collected from both upper and lower reservoirs, as previously described (34). The hESC-RPE secreted VEGF preferentially in the basal side, with a basal/apical ratio ranging from 7.4 to 9.9 (Fig. 3B). The production of VEGF on both the apical and basal side was similar for cells cultured on both types of MEC substrates of Matrigel and also BM. The resurfacing experiments of hBM are further described below.
Aged BM resurfacing by hES-RPE cells and ARPE-19
The capacity of hES-RPE and ARPE-19 to resurface the BM surface was assessed in order to investigate their ability to attach and survive on the area where RPE cells are critical for the maintenance of vision, the macula. To do so, hES-RPE and ARPE-19 were seeded on paired donor aged BM after a debridement protocol (Fig. 4A and 4B). SEM analysis revealed that ARPE-19 cells failed to attach and survive in this biologically relevant substrate, once only a few cells could be seen spread on BM (Fig. 4C and 4D). On the other hand, not only hES-RPE cells were able to completely resurface BM but also revealed a morphological resemblance to the native RPE (Fig. 4E and 4F).
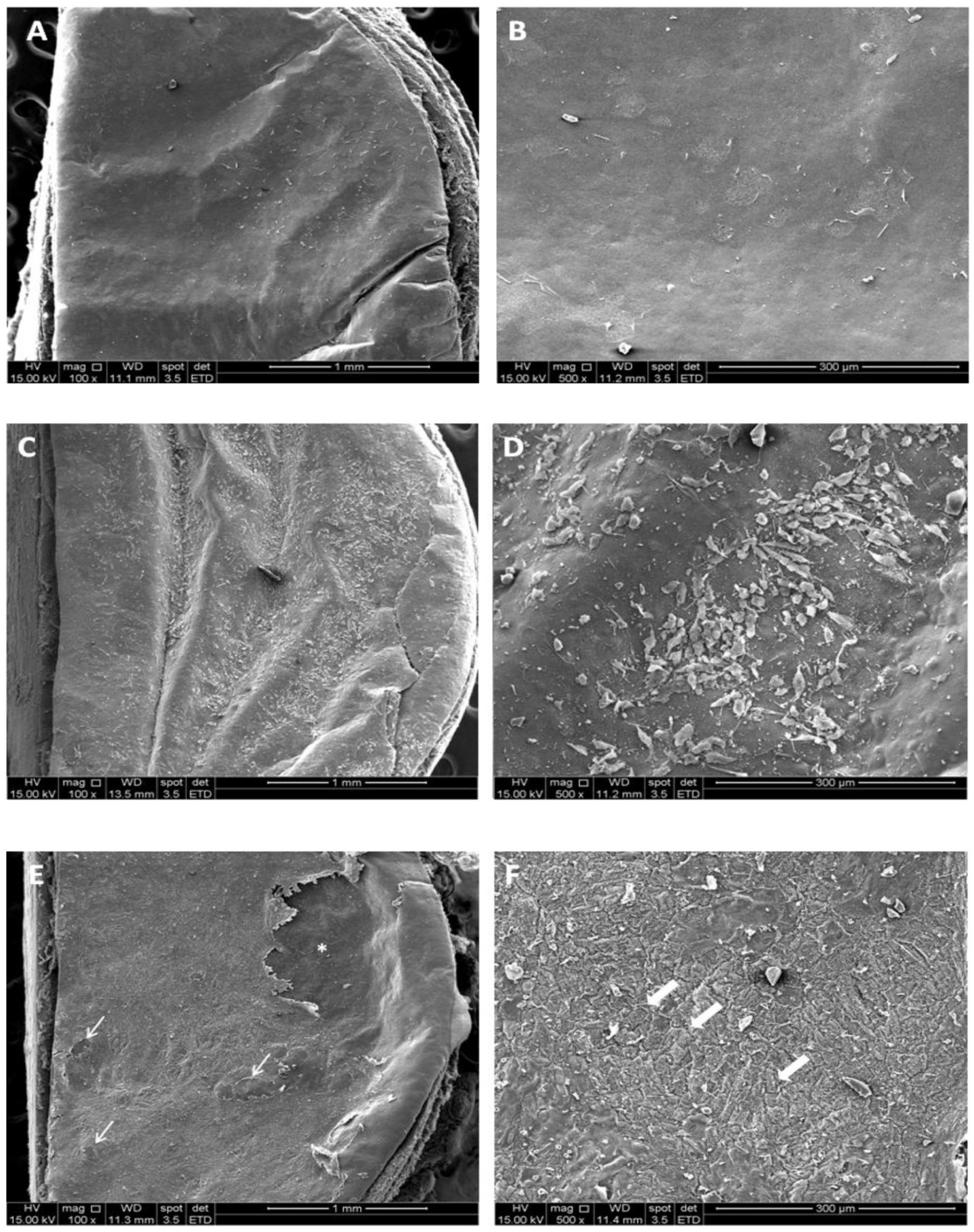
Fig. 4
Analysis of RPE cells’ ability to resurface aged human Bruch’s membrane (BM) explants. (A, B) SEM micrography of the superficial structure of the BM’s illustrating exposition of the basement membrane layer. (C, D) ARPE-19 cells seeded and cultured on BM’s were not able to proliferate and the explant was only partially resurfaced; (E, F) hES- RPE cells seeded and cultured on BM’s were able to survive, proliferate and virtually resurface the whole explant, with small flaws (narrow arrows). hES-RPE formed a cell layer with a morphology that resembles the hexagonal character of native fetal RPE (wide arrows). *Represents an artifact occurred during manipulation of the sample.
![]()
Viability assessment of hES-RPE and ARPE-19 treated with different drugs
Following the introduction of in vitro assays, which aroused as alternative methods for in vivo toxicity assessment, several groups moved to in vitro models to test drug toxicity and efficiency (4). Since our previous experiments have highlighted phenotypic and functional differences between hES-RPE and ARPE-19, we proceeded with the analysis of another possible difference between those cells - their sensibility to specific drugs. Our data showed that CSA, SRL, TAC, LEF, and TER significantly decreased cell viability in vitro in a dose-dependent manner (Fig. 5 I e II). Interestingly, for most of the tested drugs, ARPE- 19 cells viability was accentuated reduced compared to hES-RPE cells, particularly at higher drug dosages. Indeed, according to MTT cell viability assay, TER treatment (Fig. 5 IE and IIE) was significantly toxic to ARPE- 19 at >31.6 μM (p<0.0001), while to hES-RPE cells, significantly decreased cell viability at >100 μM (p< 0.0001) (IC50 values were 133.2 μg.ml−1 for hES-RPE and 103.9 μg.ml−1 for ARPE-19). LEF treatment (Fig. 5 ID and IID) results also confirmed this evidence and showed that, considering each tested concentration of this drug, ARPE-19 showed less cell viability level than hES-RPE cells, for MTT and CellTiter-BlueⓇ cell viability assays. LEF significantly reduced cell viability in both assays and for both cells at concentrations >31.6 μM (p<0.0001), and showed IC50>316 μM for hES-RPE cells, while for ARPE-19 the IC50 value was 166.70 μM.
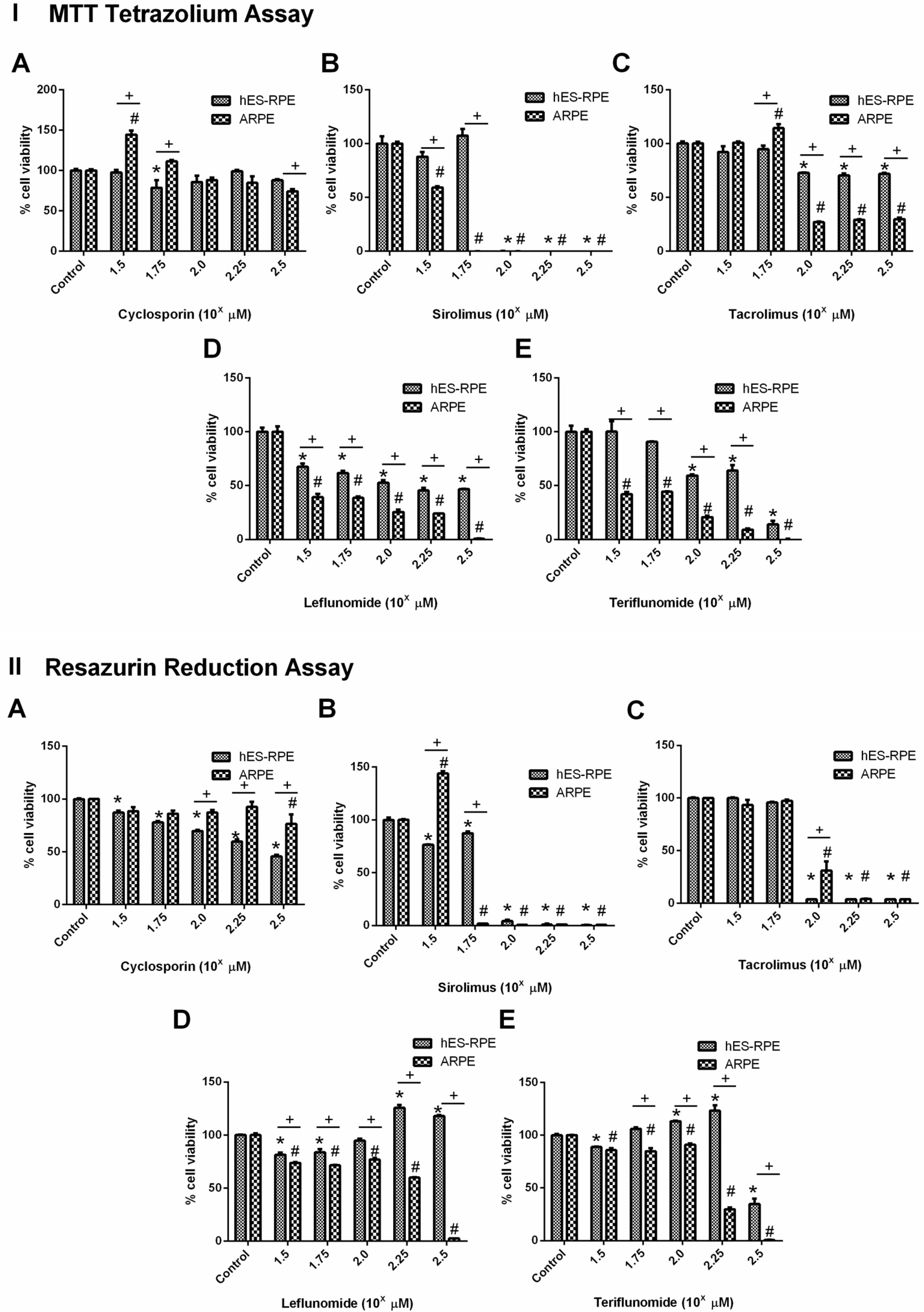
Fig. 5
Drug cytotoxicity analysis of sirolimus, leflunomide, and teriflunomide evaluated by MTT Tetrazolium Assay (I) and Resazurin Reduction Assay (CellTiter-BlueⓇ Cell Viability Assay) (II). Cyclosporin was not cytotoxic for both cell types in the analyzed doses, however, showed a reduction of cell viability greater than 50% for hES-RPE at the dose of 100 μM. Sirolimus (IB and IIB) showed higher cytotoxicity than leflunomide and its active metabolite, teriflunomide. TAC treatment (IC and IIC) showed cytotoxic for ARPE-19 at dose evaluated 100 μM (102) while it was cytotoxic for hES-ARPE at dose evaluated 176 μM (102.25). Leflunomide treatment (ID and IID) was cytotoxic for ARPE-19 at the highest dose evaluated (178 μM, 102.25) while it was not cytotoxic for hES-ARPE cells at this concentration. Teriflunomide treatment (IE and IIE), however, showed cytotoxicity for both cell types with the dose of 178 μM (102.25), and at 100 μM (102) ARPE-19 cells were also susceptible, but not hES-RPE cells. *significant difference comparing hES-RPE control and hES-RPE treatments; #significant difference comparing ARPE-19 control and ARPE-19 treatments; +significant difference comparing hES-RPE and ARPE-19 at same concentration.
![]()
Regarding SRL (Fig. 5 IB and IIB), the most toxic drug among those evaluated, we observed different cytotoxic effects on retinal cells at the lowest concentration of this chemical, according to the MTT cell viability assay. However, at 56 μM, hES-RPE cells were more resistant to SRL compared to ARPE-19 cells, for both assays. SRL was significantly toxic for ARPE-19 cells at concentrations >56 μM (p<0.0001), and >100 μM for hES-RPE (p< 0.0001) (IC50 value of 75.1 μg.ml−1 for hES-RPE while the IC50 value for ARPE-19 was lower, 48.7 μg.ml−1).
On the other hand, the results suggested that hES-RPE cells were more sensitive to CSA and TAC treatments, considering resorufin assay. TAC treatment (Fig. 5 IC and IIC) significantly reduced cell viability on both cell lines at concentrations >100 μM (p<0.0001), showing IC50= 127.74 μM for ARPE-19 and IC50=117.03 μM for hES- RPE. The CSA treatment (Fig. 5 IA and IIA) was the least toxic drug tested and the viability of the tested cells was statistically different at concentrations >10 μM (p= 0.0093) on CellTiter-BlueⓇ assay, showing IC50>316 μM for ARPE-19 and 260.4 μM for hES-RPE. Even with these differences, MTT assay demonstrated at higher concentrations of these drugs that ARPE-19 cells were more sensitive to all drugs tested. Therefore, it can be observed that, overall, ARPE-19 revealed higher sensibility to toxicological assault than hES-RPE.
Discussion
RPE cells constitute a highly desirable source of cells for therapy in AMD (14, 15), but may also be used as an animal-free option for ocular drug discovery, toxicity screening and therapy (4, 35). hES-RPE and ARPE-19 are considered two suitable options for in vitro RPE modeling (36). However, properties of the ARPE-19 line seem to change according to the way cells are maintained. In different laboratories, this line is reported to exhibit different cell morphologies, ranging from those that resemble the native RPE to morphologies that are barely compatible with an epithelial phenotype (37). Here, ARPE-19 were correctly maintained and used at the low passage to prevent loss of RPE-markers and behavior. In order to verify the molecular profile of each cell line, we have assessed the expression of RPE-specific markers. The RPE phenotype was also assessed morphologically, molecularly and functionally, by investigating hES-RPE and ARPE-19 VEGF production, melanin synthesis, as well as cellular behavior when seeded on human BM. Finally, susceptibility to chemical treatments was investigated, in order to assess the suitability of each cell model for in vitro toxicity research (Fig. 1B).
The level of mRNA expression of RPE-markers was assessed by Real-time PCR and compared to primary human RPE cells from both fetal and adult tissues since it is known that RPE goes through significant maturation during life. Obtained results show that hES-RPE cells differentiated in our hands present an expression status highly similar to fetal RPE, but also intermediate compared to adult RPE, allowing us to classify such cells as “young RPE”. In an opposite direction, ARPE-19 mRNA profile was far different from both primary RPE cells, confirming that this cell line has lost important RPE markers, with the exception of ZO-1, under aforementioned conditions of cellular maintenance and expansion. As expected, hESC-RPE presented a lower expression of pluripotency genes, compared to undifferentiated counterparts.
Even though mRNA expression may be an important indication of cellular phenotype, protein and functional assays are important for phenotype validation. Therefore, the protein expression of MITF and BEST were assessed using immunofluorescence, and the expression of RPE-65 and CRLBP were assessed using western blotting. Both analysis confirmed the mRNA expression analysis, further supporting the notion that hESC-RPE were differentiated and presented an RPE phenotype.
The functionality of both hES-RPE and ARPE-19 was investigated according to the capacity of polarized VEGF synthesis and melanin content. Furthermore, cellular behavior of both cell samples was assessed when they were seeded on a surface that mimics the area where RPE cells are originally found in the ocular globe. Due to the fact that the changes in BM engendered by aging and AMD are complex and may not be fully reversible (38, 39), any source of RPE able to regenerate this area is desired. In this sense, not only BM adhesion may add a functional evidence to support their similarity compared to functional native RPE, but also reveals a possible suitability of those cells for AMD cellular therapy. SEM analysis showed that hES-RPE were able to resurface aged BM, in stark contrast to ARPE-19, which failed to attach, survive and proliferate on this surface.
Finally, the use of stem cell-derived RPE for drug screening constitutes a powerful approach for the development of new agents. Indeed, previous studies have been shown that the safety and efficacy can be enhanced by the use of relevant human cellular models, at the same time that the use of animals for such purpose can be reduced (27, 40) CsA, SRL, TAC, and LEF are drugs with known immunosuppressive properties, fully explored as corticosteroid-sparing agents for the treatment of many inflammation processes, including ocular diseases. However, these drugs had their use limited by the poor water solubility and severe side effects. Considering their intraocular administration, it becomes crucial to analyze their cytotoxicity, for instance, by evaluating each chemical in contact with the RPE cells. According to MTT assay results, ARPE-19 is more sensitive to the toxicity effects of tested drugs compared to hES-RPE. This behavior was observed in both MTT and CellTiter-BlueⓇ cell viability assays for the majority of the tested chemicals. Despite differences, both MTT and CellTiter Blue assays indicate a more “robust” phenotype of hES-RPE cells compared to ARPE-19.
In vivo, one of the major functions of the RPE layer is the composition of blood-retina-barrier, which regulates the movement of solutes between the fenestrated capillaries of the choroid and the photoreceptor layer of the retina. Several membrane-associated transport proteins, such as multidrug resistance-associated proteins (MRPs), have been characterized at the blood-retinal-barrier and play an important role in regulating tissue bioavailability of several pharmacologic agents (41). Furthermore, has been described that the recorded levels of MRPs are significantly higher in hES-RPE compared with ARPE-19, mainly in most mature cobblestone hES-RPE. Considering that these ATP-dependent efflux transporters have a major role in drug delivery in human RPE, we can suggest that the presence of these proteins could be related to the observed results.
In summary, we have highlighted how hES-RPE and ARPE-19 cells show different profiles in vitro, in a way that the former can closely resemble native RPE while the latter fails in this aspect. It is important to use a cell source able to faithfully mimic the behavior of a healthy human cell type in order to predict important effects during preclinical research. In addition, the phenotype analysis compared to native cell types also reinforces the choice of the best source of cells for transplantation, in order to achieve better success rates in clinical phases. The generated hES-RPE seems to present the desirable features for these purposes and should, therefore, be fully explored, substituting any cell lineage that may be less representative, as well as the increasingly condemned animal-based models.
Supplementary Materials
Supplementary data including one figure can be found with this article online at https://doi.org/10.15283/ijsc20094
 XML Download
XML Download