

 PDF
PDF Citation
Citation Print
Print


Introduction
Mesenchymal stem cells (MSCs) are multipotent progenitor cells capable of self-renewing, and also undergo osteogenic, adipogenic, and chondrogenic differentiations into different types of tissues. MSCs are non-hematopoietic cells which are present fundamentally in the bone marrow and in multiple other tissues and fluids including fat tissues, umbilical cord, and molar cells of dental pulp (1-5). It has been reported that MSCs are also found in the Wharton’s jelly, amniotic fluid, lymph nodes, and in the subcutaneous tissues of the scalp (6). MSC does not express major histocompatibility complex (MHC) type II (HLA-DR). So, it cannot trigger an immune response in the recipient after allogeneic transfusion. Because of their ample availability and poor allogeneic antigenicity, MSCs have attracted a lot of attention in regenerative medicine, and in the treatment of autoimmune diseases and aging-related diseases (7). MSCs can promote the stromal environment of hematopoiesis (8). In addition, indoleamine 2,3 deoxygenase (IDO), transforming growth factor-beta (TGF-β), hepatocyte growth factor (HGF), and prostaglandin (PGE2) are very important effector molecules produced by MSCs which have a role in the immu-noregulatory characteristics of MSCs (9, 10). Stimulation of MSCs with some cytokines, such as epidermal growth factor (EGF) and basic fibroblast growth factor (bFGF) may enhance their differentiation capacity. As well, MSCs ability to regulate immune response through MHC-independent pathways enables them to be an issue of study in the treatment of diseases related to adverse’ immune response, such as autoimmune diseases (11).
Tumor necrosis factor (TNF)-like ligand 1A (TL1A) is one of the members of TNF superfamily which is referred to as TNF superfamily member 15 (TNFSF15) (12). The typical receptor for TL1A ligand is the death receptor 3 (DR3) which contain decoy receptor (DcR3) domain (13). However, it has been reported that TL1A can exert its pro- inflammatory effects on fibroblast-like synoviocytes (FLSs) through a novel receptor, TNF-receptor 2 (TNFR2) (14). Additionally, TL1A mediates inflammation in autoimmu-ne diseases, particularly those related to mucosal tissues, such as inflammatory bowel diseases. This function of TL1A has been attributed to its potential to activate Th1, Th2, and Th17, and other lymphocytes (15). Stimulation of MSCs by cytokines is becoming a hot topic because of their vital roles in MSCs autocrine, paracrine, and endocrine signaling, which provide positive outputs toward cellular therapeutic potency enhancement. One example, IL- 1β pretreatment of MSCs from synovial tissue promoted their proliferation and enhanced chondrogenic differentiation ability (16). Besides, it has been reported that IL-1β enhanced MSCs migration through MMP1/protein- activator receptor-1 (PAR1)/G-protein-coupled pathway (17). Indeed, stimulation of MSCs by TNF-α induced IL-10 and TGF-β thereby recruiting MSCs to suppress inflammation (18). But, the effect of TL1A on bone marrow MSCs (BMSCs) is still not clear. Therefore, exploring the role of TL1A cytokine ligand in the biological activities of BMSCs using its novel receptor, TNFR2 may provide a new approach to enhancing BMSCs immunomodu-latory capacity.
Hedgehog (HH) is an important signaling pathway involved in tissues growth and morphogenesis at the level of embryonic development (19). Sonic HH (SHH), Indian HH (IHH), and desert (DHH) ligands, and patched 1/2 (PTCH1/2) and smoothened (Smo) trans-membranous receptors, as well as the target gene of Hedgehog, Gli, are all the members of HH signaling pathway. Recently, scientists declared a promising potential for SHH in the regeneration of cardiac tissues in an animal model, suggesting a role for HH signaling in improving symptoms of aging-related diseases (20). Another study revealed that IHH may induce the Wnt antagonist, SFRP1 (21), which may keep normal differentiation of MSCs and secure them from the aging fate. In our previous results, we discovered that IHH has an anti-aging effect on BMSCs and can promote the potency of BMSCs (22). Moreover, it has been reported that hedgehog signaling induces osteogenesis at the cost of adipogenesis in MSCs (23). Furthermore, IHH promoted the function of BMSCs in cartilage differen-tiation (24). The interaction of IHH with TL1A has been underscored in the inflammatory process through TNFR2 (25). Thus, the study of TL1A/IHH signaling in the therapeutic potency of BMSCs may introduce a new pathway to enhance cellular therapy using BMSCs.
In the current study, we aimed to investigate the roles of TL1A in BMSCs’ biological behaviors through the novel receptor TNFR2. In addition, we tested the effects of TL1A stimulation on the anti-inflammatory property of BMSCs using FLSs from patients with rheumatoid arthritis (RA) and T cells. Moreover, due to its importance in the immunoregulatory potency of MSCs, we studied the interaction between TL1A and IHH using IHH small interference RNAs (siRNAs).
Go to : 
Materials and Methods
Cell lines and human samples
Approval for the study was granted by the Dalian Medical University ethical committee on human and animal research. BMSCs and H9 human T lymphocytes (H9 T) cell lines were purchased from Dalian Chenyu Biotech-nology Co., Ltd, China. Jurkat human T lymphocytes were collected as a gift from Peking University People’s Hospital. Synovial tissues from RA patients were collected from the Orthopedics Department of the Second Affiliate Hospital of Dalian Medical University, Dalian, China, for the isolation of FLSs.
FLSs isolation
Minced synovial tissues from RA patients were enzymatically digested with collagenase type I from Clostridium histolyticum (Biotopped, China) at a concentration of 3 mg/ml in DMEM (Biological Industries, USA) in a petri dish and at 37℃ on a shaker for 2 h. The tissues filtrate from 70 μM pores of nylon mesh was centrifuged for 5 minutes at 300 g and the sediment was resuspended in adequate volume of Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 20% fetal bovine serum (FBS) (Biological Industries, USA), 100 U/ml penicillin, and 100 μg/ml streptomycin (P/S), and cultured in humidified environment at 37℃ and 5% CO2. Three to five passages of RA-FLS were used for all experiments.
IHH silencing
Lipofectamine 2000 (Invitrogen, Guangzhou, China) and small interference RNAs (siRNAs) were used for IHH knockdown (22). Both siRNA and lipofectamine were separately incubated in 150 μl DMEM at room temperature (RT) for 5 minutes, mixed thoroughly in RNase free sterile tube, and incubated again for 25 minutes at RT. The mixture was then added to BMSC of 60∼90% confluence in 6-well plat in 1ml DMEM without FBS, and incubated at 37℃, 5% CO2 and 95% humidity. The cells were used in experiments after 24 or 48 h incubation. The transfection effect tested by both reverse transcription-polymerase chain reaction (RT-PCR) and western blot (22). Validation of the transfection done with FAM negative control, GAPDH positive control, and siRNA negative control (Table 1) (Gene Pharma, Shanghai, China).
Table 1
List of primers
![]()
Cell culture and treatments
Adequate-confluence of BMSCs were propagated in DMEM with 10% FBS, 100 U/ml/100 μg/ml P/S at 37℃, and 5% CO2 in a humidified environment. BMSC characteristics tests were performed using primary antibodies conjugated with fluorescein isothiocyanate (FITC), phycoerythrin (PE), or phycoerythrin-cyanin 5/7 (PC5/7) against CD14, CD34, CD45, CD73, CD90, and CD105 by Accuri C6 flow cytometer (BD Biosciences, San Jose, CA, USA). The results are shown in our previous article (22). 1×106 cells per well were stimulated with TL1A (100 ng/ml), and treated with 1.5 μg/ml TNFR2 antibodies (R&D, USA) or recombinant IHH (rIHH) 300 ng/ml (Prospec, USA) in serum-free DMEM for 24 h. Conditioned medium (CM) from TL1A-stimulated, non-stimulated BMSC, BMSCs with IHH siRNA and TL1A, and rIHH- treated BMSCs after incubation in serum-free DMEM for 24 h were collected, and 20% of CM in either RPMI-1640 or DMEM were used to incubate human T lymphocytes and RA-FLSs respectively. The CM is new medium added to the BMSCs cells after 24h stimulation by TL1A or rIHH treatment.
Flow cytometry analysis of TNFR2 expression
Flow cytometry analysis was performed using fluorochrome-conjugated antibodies. Briefly, 106 BMSCs were trypsinized and washed in PBS, and then the pellets were re-suspended in fluorescent-activated cell sorting (FACS) buffer. These single-cell suspensions were incubated in the dark at 4℃ for 30 min with PE-conjugated antibodies against human TNFR2 and isotype control antibody (from eBioscience, USA). Then, the cells were centrifuged, re-suspended in PBS and analyzed using a flow cytometer (Agilent, USA). The data were collected and analyzed using the novoexpress (Agilent, USA) software (Fig. 1G).
 | Fig. 1TL1A induced paracrine activity of BMSCs. (a∼f) Cytokines expression in BMSCs. GAPDH used as reference. (g) The expre-ssion of TNFR2 in BMSCs. Results are representative of three inde-pendent experiment, and data presented as mean±SEM. *p< 0.05, **p<0.01. ****p<0.0001 compared to control. ##p<0.01, ###p<0.001 compared to TL1A treatment.
|
RT-PCR
RNAiso (Takara Bio, Japan) was used to extract mRNA of BMSCs, H9 human T lymphocytes, or FLSs as described in the previous article (26). cDNA was prepared using PrimeScriptTM 1st strand cDNA Synthesis Kit (Takara Bio, Japan) and 1 μg of extracted mRNA. The internal control was Glyceraldehyde 3-phosphate dehydrogenase (GAPDH). Primers (Takara Bio, Japan) shown in Table 1 were used to amplify genes after being mixed with PCR MasterMix 2X Power Taq (Bio Teke Corporation, China), cDNA, RNase Free dH2O in micro-tubes according to the manufacturer’s instructions, and then incubated in the thermal cycler (Bio-Rad, USA). Each cycle consisted of 30s for denaturation at 95℃, 30s of annealing at 56.0, 56.5, 57, 57.5, or 58.0℃, and 30s for extension at 72℃, for a total of 35 cycles. Electrophoresis in tris acetic acid EDTA (TAE) buffer using 2% agarose gel was applied to separate and PCR products. Image Lab detection system (Bio-Rad, USA) or Image J (ImageJ2x, Rawak Software Inc., Germany) were used to quantify PCR bands.
BMSCs in vitro bi-lineage differentiation assay
For adipogenesis, BMSCs of 80% confluency in 24-well plate were incubated with 10% FBS in MEM-ALPHA medium (Biological Industries, USA) and differentiation mixture containing 100 U/ml/100 μg/ml P/S, 0.5 mM 1-methyl-3-isobutylxanthine, 1 μM dexamethasone, 10 μg/ml insulin, and 0.2 μM indomethacin for 21 days at 37℃ with 5% CO2 and in humidified atmosphere. The medium was replaced every three days. Adipogenesis was assayed by the quantity of lipid or droplets accumulation which appear yellow-red after staining with Oil-Red-O lipophilic dye (Coolaber, China). 0.1 μM dexamethasone, 0.05 mM ascorbic acid, and 10 mM glycerophosphate were added to prepare osteogenic differentiation medium. Osteogenesis was measured by the intensity of calcium deposition which stains red-orange by Alizarine-Red-S stain (Coolaber, China).
BMSC colony-forming assay
Complete medium; 10% FBS in MEM-ALPHA and 100 U/ml/100 μg/ml P/S was used for seeding 2600 BMSCs up to 12 days at 37℃ 5% CO2 and humidity. The cells were stained by 0.2% crystal violet after being washed using PBS and fixed by 1% paraformaldehyde. Photos were taken from at least five different fields per well, and the average colony count estimated.
Tranwell migration assay
1×105 cells/ml of BMSCs were suspended in 100 μl 5% FBS in DMEM medium and loaded into the upper chamber of the transwell inserts of 8 μm pore membrane (Costar, USA). 350 μl chemo-attractant (20% FBS in DMEM medium) was added to the lower chamber in a 24-well plate. The non-migrated cells were carefully removed from the inner part of the insert by a cotton-swab applicator. After 24h incubation at 37℃, and 5% CO2, 70% ethanol was used to fix the cells for 10 minutes, and then stained with 0.2% crystal violet at RT° for 20 minutes. Migrated BMSCs were counted from five random fields at 100 magnifications in each membrane.
Wound healing migration assay
A 200 μl pipette tip was used vertically to generate wound through 100% confluency BMSCs in a 12-well plate. Cells debris were removed using culture medium and then pictures of the wound were taken at 0 h by an inverted microscope (Olympus 1X71, Japan). Cells were incubated at 37℃ and 5% CO2. Snapshots were taken after 24 hours. Evaluation of BMSCs migration was done by measuring the distance crossed by the migrated cells through the wound.
ROS quantification
ROS generation levels were measured using 2’,7’- Dichlorofluorescin diacetate (DCFHDA; Sigma-Aldrich, USA). 70% confluency BMSCs were incubated in DMEM with 5 μM DCFHDA at 37℃ with 5% CO2 and humidity for 30 min. Cells were washed two times with PBS and trypsinized, and analyzed by Accuri C6 flow cytometer (BD Biosciences, San Jose, CA, USA) at 488 nm and 538 nm wavelength. In addition, stained cells were tested by fluorescence microscope (Olympus1X71, Japan) to view and capture images.
Western blot
More than 106 harvested BMSCs were digested in a lysis buffer cocktail and centrifuged at 10000 xg for 15 minutes at 4℃ to get the supernatant as the total protein according to the kit protocol (KeyGen Biotech, China). 20 μg of extracted protein with a loading buffer were loaded in wells of 10% sodium dodecyl sulfate-polyacrylamide gel (SDS-PAGE) for electrophoresis, and then blotted to a polyvinylidene fluoride (PVDF) membrane (Millipore Co., USA). Membranes were incubated in blocking buffer (5% skimmed milk in tris-buffered saline tween 20, TBST,) and then with indicated primary antibodies overnight at 4℃. After washing the primary antibodies, the membranes were incubated for 1h at RT° with secondary antibody conjugated with horseradish peroxidase. After TBST- washing, the membranes were treated with enhanced chemiluminescent (ECL) reagents (Bio-Rad, USA) for imaging. Image Lab detection system (Bio-Rad, USA) was used for protein bands imaging and analysis.
ELISA
The IL-6, IL-18, IL-1β and IL-8 levels in the supernatants of each coculture system (BMSCs co-cultured with Jurkat T cells and BMSCs co-cultured with RA FLS) were measured using their respective ELISA kits (R&D Systems, USA). All procedures were performed as described in the manufacturer’s instructions. The absorbance was measured at 450 nm. Three wells were used for each medium sample. The assays were repeated three times.
Apoptosis assay
5 μl fluorescein isothiocyanate-conjugated Annexin V (FITC Annexin V; BD Biosciences, USA) and 5 μl propidium iodide (PI; BD Biosciences, USA) were used to stain 1 million washed BMSCs in 200 μl 1X Annexin V binding buffer, and then incubated for 15 minutes at RT°. Flow cytometry Accuri C6 (BD Biosciences, San Jose, CA, USA) was used to analyze the suspended cells.
Statistical analysis
Statistical analysis was performed using Prism 5 software (GraphPad Software Inc.). Paired sample T-test was used to compare differences between and among the group. All values are presented as means±SEM, and a p-value less than 0.05 was considered significant.
Go to :
Results
TL1A increased expressions of BMSCs paracrine-related genes
To assess the role of TL1A on BMSCs paracrine activity, we treated the cells with 100 ng/ml TL1A or TL1A+anti-TNFR2 and then investigated the expressions of some paracrine-related genes. TL1A significantly induced mRNA expression of COX-2, IL-6, IDO, TGF-β, and HGF in BMSCs (Fig. 1). However, when the cells were treated with TL1A+anti-TNFR2, the increase expression of all the genes was reversed with variable significance, as the decrease in IL-6 and HGF were not statistically significant (Fig. 1C and 1D). Collectively, these findings suggest that TL1A induced the paracrine secretions of BMSCs via TNFR2.
TL1A induced osteogenesis, decreased adipogenesis and has no effect on apoptosis of BMSCs
Differentiation capacity is one of the most important characteristics which give the BMSCs the ability to regenerate tissues in cellular therapy. To investigate this property in presence of TL1A, we incubated BMSCs in adipogenic and osteogenic differentiation media after treatment with TL1A or TNFR2. Differentiation was measured by Alizarin Red S staining for osteogenic differentiation and Oil Red O for adipogenic differentiation. TL1A increased osteogenesis shown by increased red-orange calcium depositions, but anti-TNFR2 decreased this surge (Fig. 2A and 2B). Experiments of adipogenic differentiation revealed that TL1A inhibited adipogenesis indicated by decreasing yellow-red fat droplet, whereas anti-TNFR2 treatment restored this inhibition (Fig. 2C and 2D). In other experiments, TL1A was not affect the survival of BMSCs through TNFR2 (Fig. 2E and 2F). Our results revealed that TL1A could correct the biased differentiation of BMSCs to favor osteogenesis.
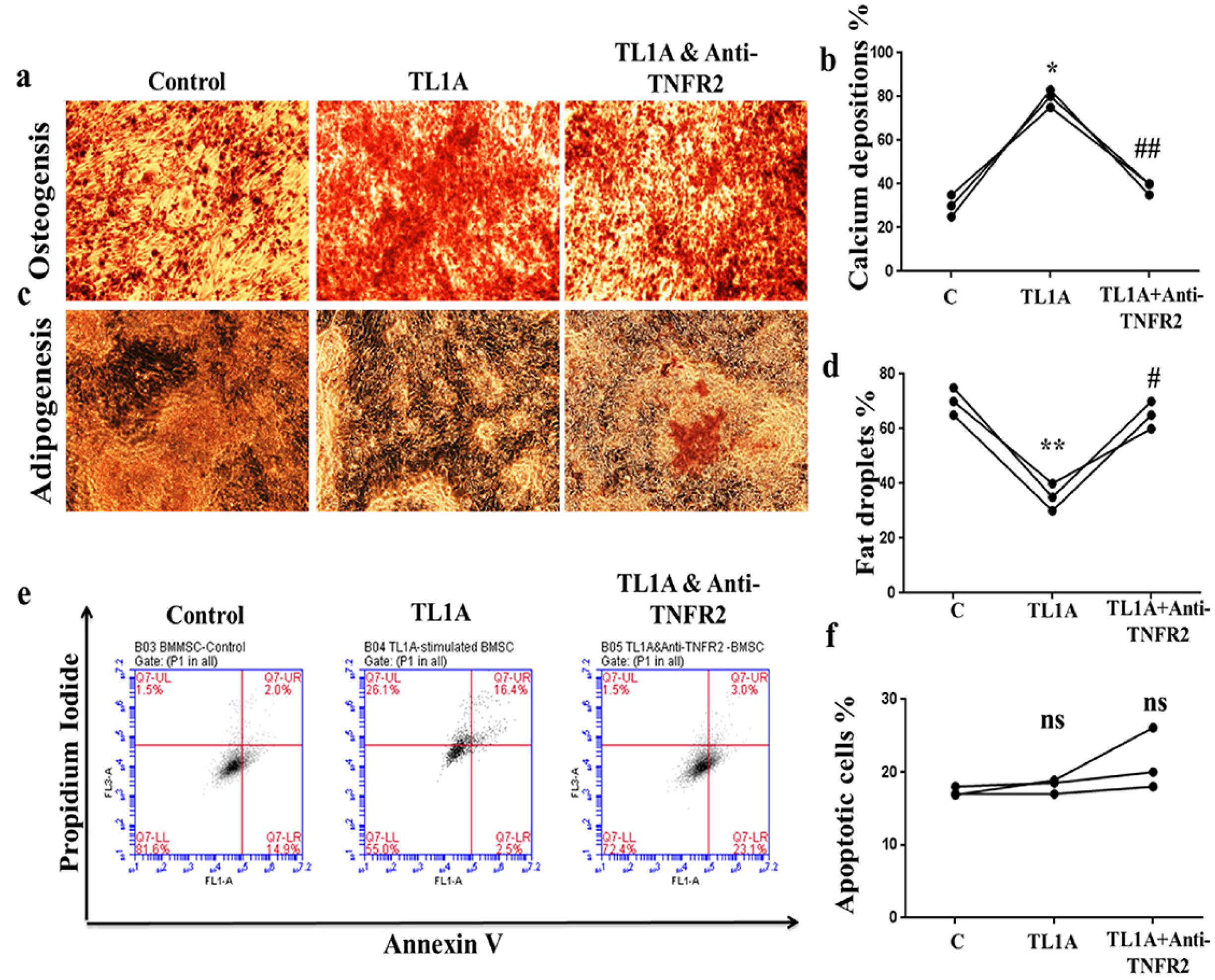 | Fig. 2TL1A promoted osteogenic differentiation at the cost of adipogenic differentiation. (a, b) Osteoge-nic stain and measure of stain inten-sity. (c, d) Adipogenic stain and measure of stain intensity. (e, f) Flow cytometry of cells apoptosis and apoptotic cells percentages. Data presented as mean±SEM from three independent experiments. ns>0.05, *p<0.05, **p<0.01 compared to control. #p<0.05, ##p<0.01 compared to TL1A treatment.
|
TL1A enhanced BMSCs proliferation and migration
Proliferation and migration of BMSCs are the main properties in the therapeutic activities MSC. Thus, colony formation assay, and transwell and wound closure assays were applied to measure the roles of TL1A on BMSCs proliferation and migration. The results of the CFU showed an increased count of colonies after 12 days of treatment and continuous culture of BMSCs treated with TL1A. However, the colony count was reversed after treatment with anti-TNFR2. (Fig. 3A and 3B). Interestingly, the results of cell migration revealed that TL1A significantly increased migrated cells in the transwell experiments (Fig. 3C and 3D) and the speed of cells in the wound healing assay (Fig. 3E and 3F). More importantly, treatment of BMSCs with anti-TNFR2 in the presence of TL1A decreased the migration ability. These observations indicate that TL1A enhances BMSCs’ biological activities through promoting both proliferation and migration via TNFR2.
 | Fig. 3TL1A promoted BMSCs proliferation and migration. (a, b) Colony forming assay after 12 days of treatments and continuous culture. (c, d) Transwell migration assay after 24h of treatment. (e, f) Wound healing assay for 0 h and 24 h after scratch. Results are representative of three independent experiment, and data presented as mean±SEM. *p<0.05, **p<0.01 compared to control. #p<0.05, ##p<0.001 compared to TL1A treatment.
|
TL1A-stimulated BMSCs conditioned medium has an anti-inflammatory effect on human T lymphocytes and FLSs from rheumatoid arthritis patients
To assess the effect of TL1A induced stimulation on the therapeutic efficiency of BMSCs, we incubated H9 T cells and RA-FLSs with conditioned medium collected from TL1A-stimulated BMSCs (TL1A stimulated-BMSC-CM) or without TL1A (BMSC-CM). In H9 T cells, genes expressions of inflammatory markers, IL-6, IL-8, TNFα, MMP2, MMP9, C-fos, C-Jun, and Sp-1 were down-regulated significantly by TL1A stimulated-BMSC-CM coupled with mild down-regulation of IL-1β, RANKL, and COX2 expressions (Fig. 4A∼L). Also, ELISA results showed that TL1A stimulated-BMSCs-CM suppressed inflammatory cytokines; IL-6, IL-18, IL-1β, and IL-8 in jurkat T lymphocytes (Fig. 4M∼P). In RA-FLS, RT-PCR results showed down-regulation in expressions of RANKL, COX2, MMP1, MMP2, MMP9, MMP13 and, C-Jun following treatment with TL1A stimulated-BMSC-CM. However, genes transcription of NF-kB and VEGF assay revealed undetectable bands with TL1A stimulated- BMSC-CM treatment (Fig. 5A∼J). In accordance with these data, checking the levels of inflammatory cytokines; IL-6, IL-18, IL-1β, and IL-8 by ELISA revealed anti-inflammatory effect for TL1A stimulated-BMSC-CM (Fig. 5K∼N). These findings suggest TL1A could promote BMSCs’ ability in alleviating inflammation in patients with inflammation, for example RA patients.
 | Fig. 4TL1A increased anti-inflammatory effect of BMSCs on human T cells. Representative RT-PCR bands and quantification of inflammatory cytokines expressed by H9 T cells (a∼l). GAPDH used as reference. (m∼p) IL-6, IL-18, IL-1β, and IL-8 levels in jurkat T cells measured by ELISA. Results are representative of three independent experiment, and data presented as mean±SEM. ns>0.05, *p<0.05, **p<0.01, ***p<0.001 compared to BMSC-CM.
|
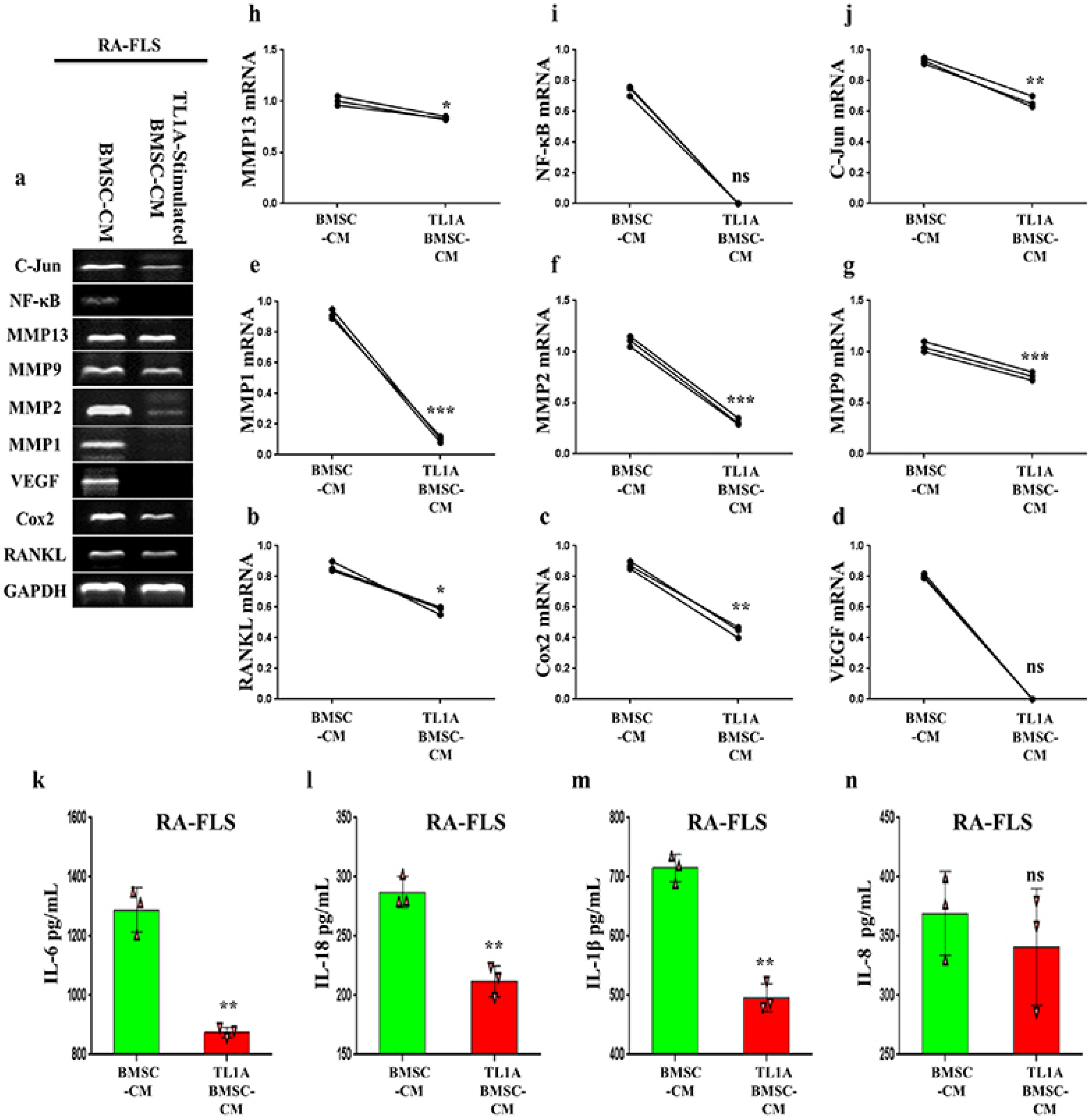 | Fig. 5TL1A induced BMSCs anti-inflammatory effect on FLSs from rheumatoid arthritis patients. Representative RT-PCR bands and quantification of inflammatory cytokines expressed by RA-FLSs (a∼j). GAPDH used as reference. (k∼n) IL-6, IL-18, IL-1β, and IL-8 levels in RA-FLSs cells measured by ELISA. Results are representative of three independent experiment, and data presented as mean±SEM. ns>0.05, *p<0.05, **p<0.01, ***p<0.001 compared to BMSC-CM.
|
TL1A-promoted IHH associated with NF-κB and STAT3 signaling up-regulation, and p53 and oxidative stress down-regulation in BMSCs
To investigate how TL1A activate BMSCs, we decided to assay IHH pathway due to its importance in MSCs potency (22), and because our previous results discovered that TL1A can regulate IHH pathway in FLSs (25). Surprisingly, results of RT-PCR experiments showed that TL1A promotes transcription of IHH ligand and its trans-membranous receptors, PTCH1/2, but reversed by anti-TNFR2 in BMSCs (Fig. 6E∼G). Also, IHH translation was assayed by western blot, and consistently; the results presented increased IHH protein after TL1A stimulation and decreased due to anti-TNFR2 treatment (Fig. 6A and 6D). In further experiments, we assessed the activity of NF-kβ, STAT3, p53, and ROS signaling because of their importance in BMSCs stemness and association with HH signaling. Results of the western blot assay revealed up-regulation of p65-NF-kβ, and p-STAT3 through TL1A/TNFR2 axis (Fig. 6B and 6C). However, RT-PCR results of p53 displayed down-regulation by TL1A stimulation but not through TNFR2 (Fig. 6E and 6H). Additionally, stimulation of BMSCs by TL1A decreased ROS generation (Fig. 6I∼K). These findings suggest an important role for IHH signaling in the effects exerted by TL1A on BMSCs stemness. For further confirmation, we used rIHH protein to explore the role of IHH in BMSCs stemness. BMSCs were treated with rIHH and incubated for 24 h before collection of rIHH-treated BMSC-CM. After incubation of jurkat T lymphocytes and RA-FLSs with rIHH-treated BMSC-CM for 24 h, we found that IL-18, IL-1β, and IL-8 cytokines levels were compromised in both cells (Fig. 7).
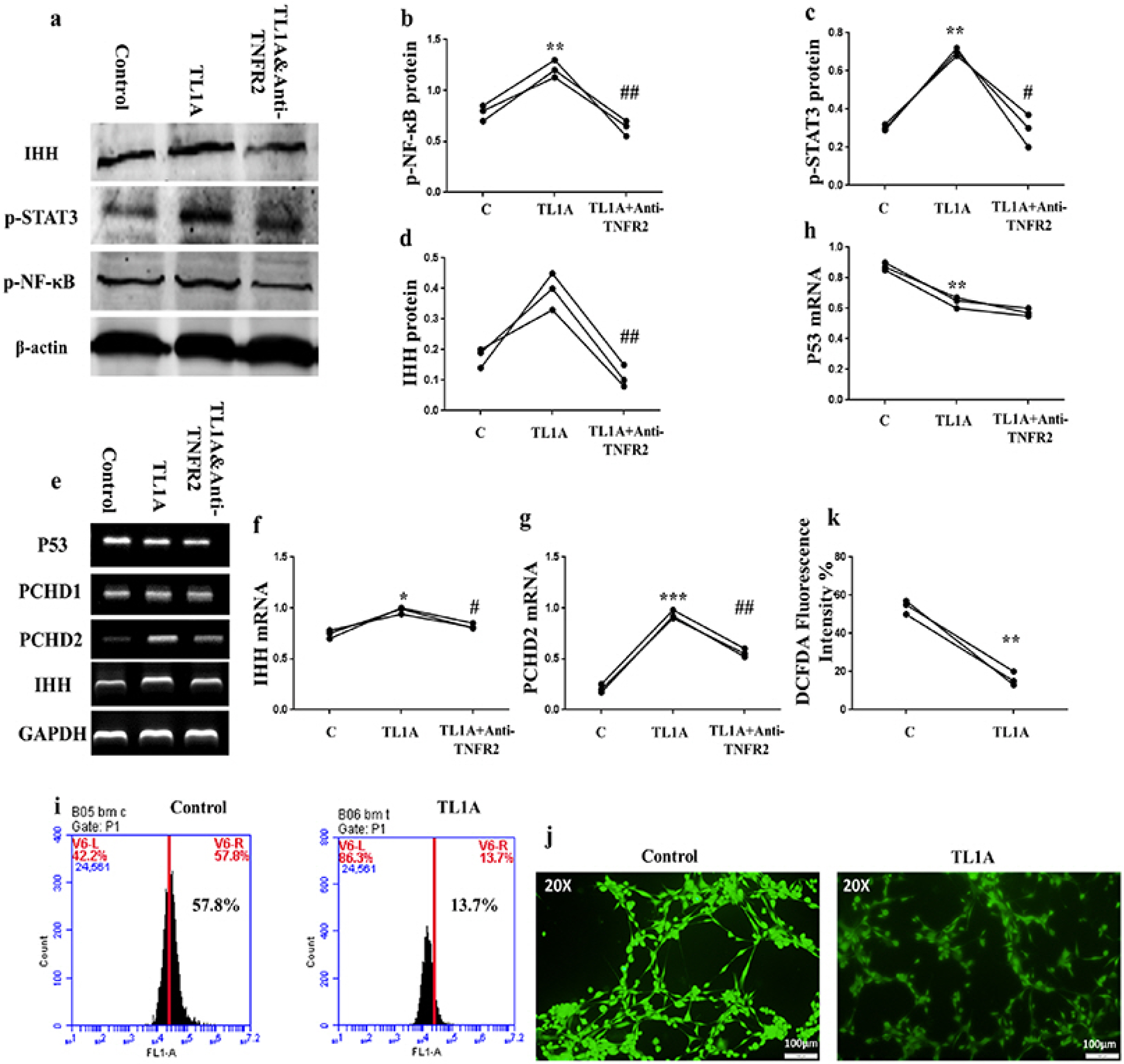 | Fig. 6TL1A modulated IHH expression and stemness-related signaling pathways in BMSCs. (a∼d) Representative western blot bands and relative band intensity of p65-NF-kβ, p-STAT3, and IHH. (e∼h) PCR bands and estimation of mRNA expression of some aging-related genes. β-actin and GAPDH used as references. (i∼k) Representative flow cytometry and microscopic (at 20X magnification) images and quantification of ROS generation. Results are representative of three independent experiment, and data presented as mean±SEM. *p<0.05, **p<0.01, ***p<0.001, ****p<0.0001 compared to control. #p<0.05, ##p<0.001 compared to TL1A treatment.
|
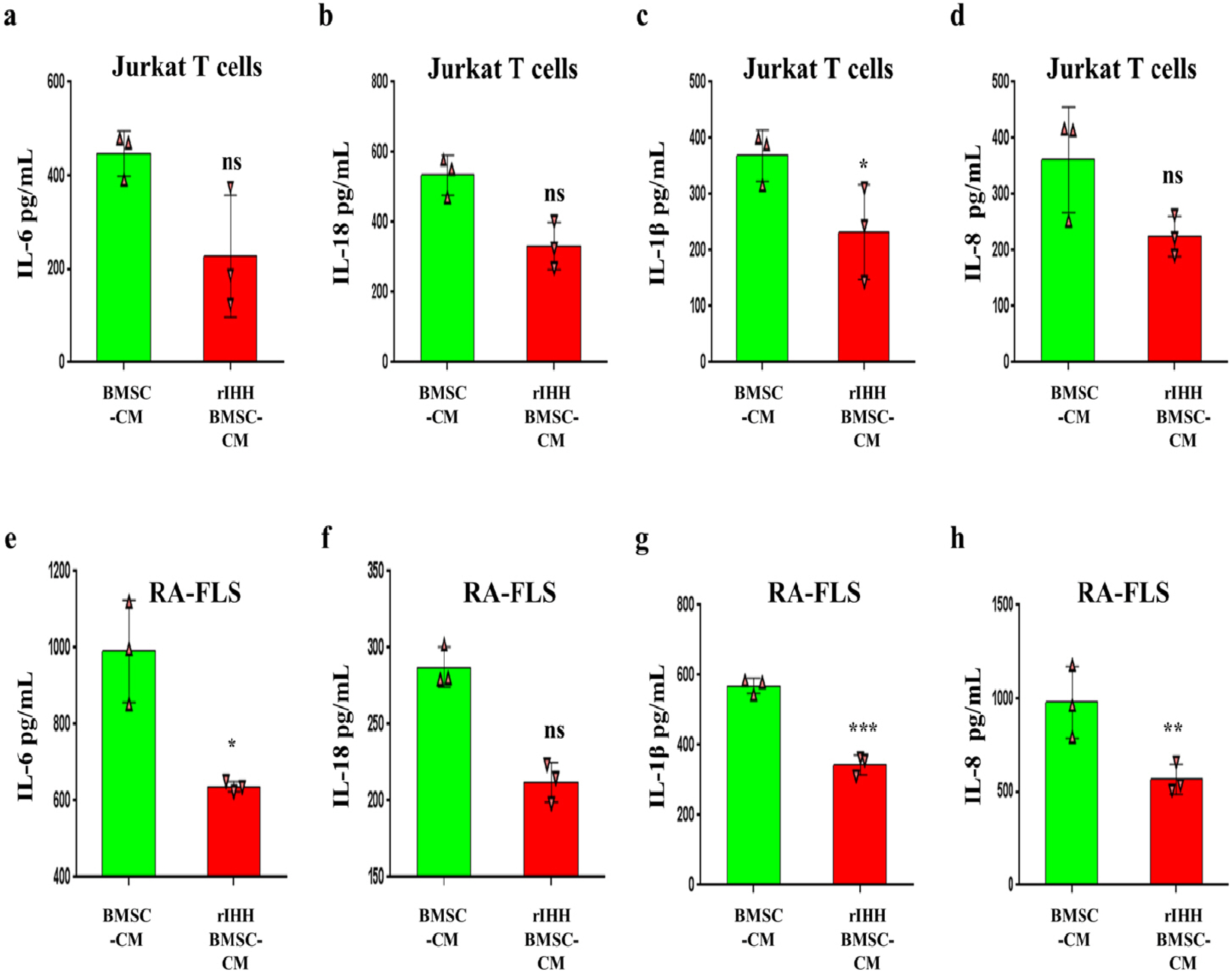 | Fig. 7IHH induced anti-inflammatory power of BMSCs. (a∼d) IL-6, IL-18, IL-1β, and IL-8 levels in jurkat T cells measured by ELISA. (e∼h) IL-6, IL-18, IL-1β, and IL-8 levels in RA-FLSs cells measured by ELISA. Results are representative of three independent experiment, and data presented as mean±SEM. ns>0.05, *p<0.05, **p<0.01, ***p<0.001 compared to BMSC-CM.
|
IHH is involved in TL1A-dependent BMSCs immunomodulation promotion
To evaluate the interaction of TL1A with IHH in the therapeutic potency of BMSCs, we used IHH siRNA to silence IHH gene and prepared conditioned medium in the presence of TL1A (TL1A-BMSC-IHH-CM). The efficiency of IHH was assessed by RT-PCR, western blot and FAM negative control (22). The control group’s conditioned medium was TL1A-stimulated BMSCs with negative control siRNA (Control-CM). After incubating H9 T cells with the two conditioned media, we assayed for the gene expression of some inflammatory mediators; IL-1β, IL-8, RANKL, TNFα, MMP1, MMP13, and COX2. Intere-stingly, we observed that H9 T cells treated with TL1A- BMSC-IHH-CM expressed the inflammatory mediators (Fig. 8A∼D, F∼H) significantly more than the H9 T cells treated with Control-CM except for MMP13 (Fig. 8A and 8E) which showed no difference between the groups. In addition, incubation of RA-FLSs with conditioned medium of the control and BMSCs with IHH siRNA revealed increased expression of IL-6, RANKL, C-fos and Sp-1 inflammatory molecules in TL1A-BMSC-IHH-CM groups compared to control groups (Fig. 8I∼M). The findings indicated that TL1A may play a role in increasing BMSCs stemness through IHH signaling, as IHH-depleted BMSCs failed to inhibit inflammation in H9 T cells and FLSs of rheumatoid arthritis patients despite the presence of TL1A.
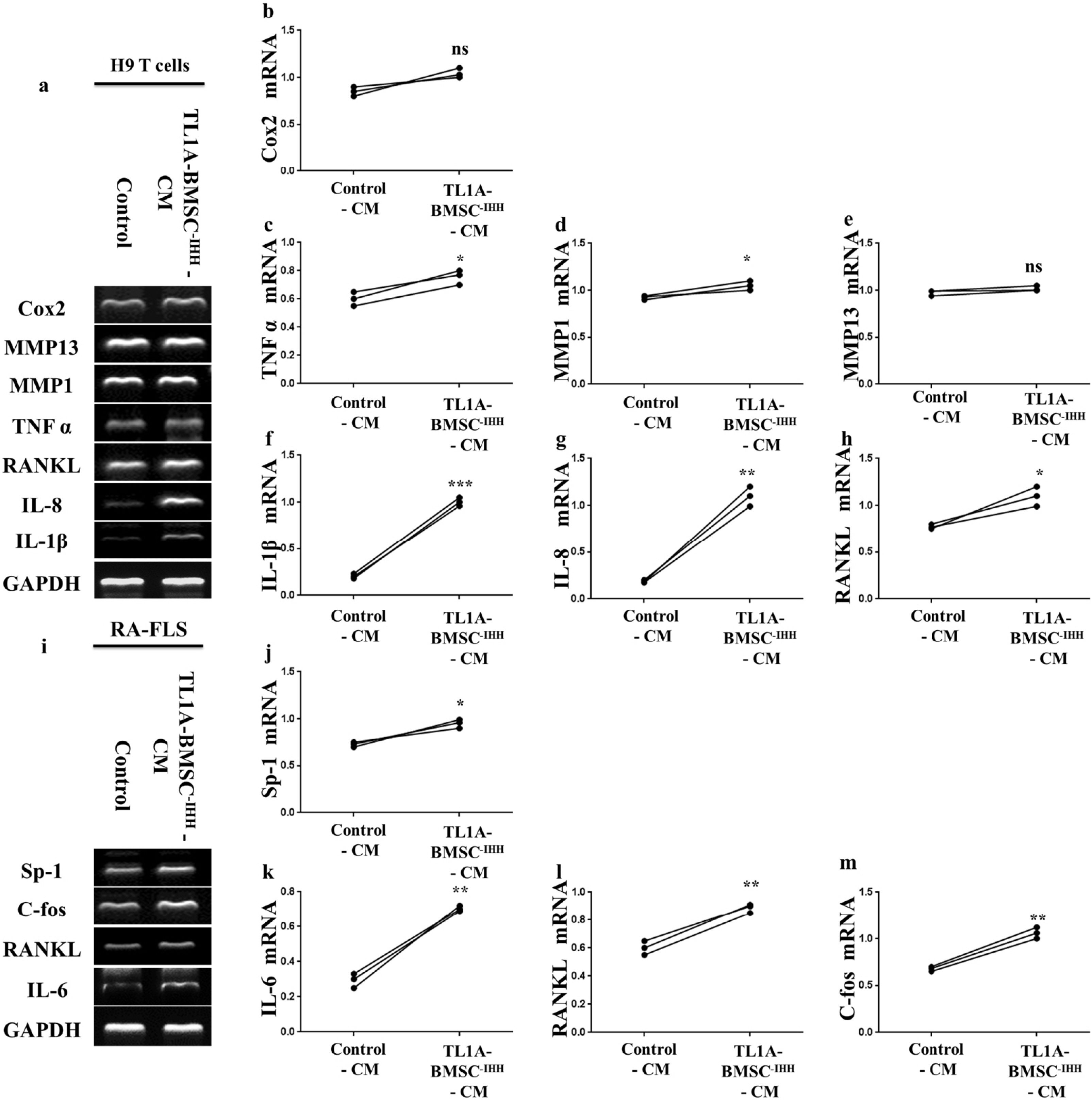 | Fig. 8Effect of TL1A-BMSCs-IHH conditioned medium on H9 T cells and FLSs from rheumatoid arthritis patients. Representative RT-PCR bands and quantification of inflammatory mediators expressed by H9 T cells (a∼h), and RA-FLS (i∼m). GAPDH used as reference. Results are representative of three independent experiment, and data presented as mean±SEM. ns>0.05, *p<0.05, **p<0.01, ***p<0.001 compared to BMSC-CM.
|
Go to :
Discussion
This study revealed an activation effect of TL1A on BMSCs in which the paracrine parameters, COX-2, IL-6, IDO, TGF-β, and HGF were induced after treatment with TL1A and reversed by anti-TNFR2 antagonist. These genes are closely related to the paracrine activity of BMSCs, and the findings suggest that TL1A could promote the immunoregulatory capacity of BMSCs. In a previous study, it was shown that preconditioning MSCs with IL-1β suppressed LPS-induced inflammation in microglial cells via IL-1 receptor type1 (IL-1R1) and granulocyte-colony stimulating factor (G-CSF) (27). Also, stimulation of BMSCs with IL-1β and IFN-γ modulated ma-crophage polarization and promoted its anti-inflamma-tory ability (28), while human amnion-derived MSCs enhanced the functions of rat ovaries through their paracrine secretions including HGF (29).
TL1A/TNFR2 axis promoted BMSCs differentiation toward osteogenesis at the cost of adipogenic differentiation, and enhanced proliferation and migration. Such observations introduce new proof in TL1A’s ability to support BMSCs in the treatment of bone-loss diseases. Another cytokine from the same family, TNF-α promoted adverse microRNAs expression in MSCs exosomes (30), and induced bone regeneration and immunoregulatory potency following TNF-α and IFN-γ preconditioning (31). On the other hand, this study showed that TL1A cannot change apoptotic cells counts after stimulation of BMSCs in cell culture. Conversely, another cytokine, IL-8 was reported as a protector for BMSCs which supported their proliferation through decreased apoptotic cells percentage by Akt-STAT3 pathway (32). Furthermore, BMSCs treated with IL-8 displayed enhanced therapeutic potency in bone formation characterized by osteogenesis and chondrogenesis up-regulation by activating PI3K/Akt pathway in CXCR2-dependent manner (33). It has also been reported that MSCs migration could be stimulated through cytokines (34). Seemingly, TL1A enhances BMSCs stemness through correcting biased differentiation and increasing proliferation and migration.
Our results showed that TL1A induced IHH, NF-κB, and STAT3 pathways and down-regulated p53 and ROS in BMSCs. In addition, the conditioned medium of TL1A-stimulated BMSCs has an anti-inflammatory effect on inflammatory cells derived from RA patients. Consequently, this study uncovered that TL1A promoted BMSCs immunomodulatory potency through IHH signaling pathway. Compatible with our observations, pretreatment of BMSCs with IFN-γ mediated their suppressing power on activated lymphocytes (35) through increased expression of immunosuppressive molecules, TGF-β, HGF, and PGE2 (36). Meanwhile, a core player in aging, P53, may have an effective interaction with hedgehog signaling (37) and this may have direct contributions to the senescence process and stemness of BMSCs. IHH, a hedgehog homolog related to cartilage and bone formation is considered an inducer in chondrogenesis of human MSC (38). Going on, NF-κB-mediated IL-4 increased the anti-inflammatory property of BMSCs (39), and an engineered MSCs was reported to have a good influence in the treatment of autoimmune inner ear inflammation through overexpressing of IL-4 (40). Besides, IL-7 promoted renal regeneration by enhancing the fusion ability of rat BMSCs (41). Moreover, IL-17-stimulated MSCs increased survival of skin transplant from allogenic source (42), and IL-17a-pretreated MSCs characteristically increased the therapeutic efficiency in renal diseases (43), whereas IL-17b promoted the immunoregulatory potency of MSCs in the treatment of gastric diseases (44). IL-22 promoted MSCs migration, proliferation, and osteogenic differentiation (45). Moreover, IL-23 may have a role in promoting MSCs immunoregulatory ability in the treatment of autoimmune disease through up-regulation of IL-10 and down-regulation of IL-17 in an inflammatory niche (46).
Conditioned medium derived from MSCs is used in clinical applications due to its anti-inflammatory, antioxidant, and regenerative ability. It is believed that the therapeutic potential of MSCs’ conditioned medium is attributed to the secretion of exosomes which contain regulatory molecules of MSCs, cytokines and another paracrine secretome (47). In this study, we found increased expressions of IL-1β, IL-8, RANKL, TNFα, MMP1, and COX2 genes in H9 T cells treated with TL1A- BMMSC-IHH-CM when compared to Control-CM control groups. Moreover, incubation of RA-FLSs with TL1A- BMMSC-IHH-CM revealed increased expressions of IL-6, RANKL, C-fos, and Sp-1 genes more than in RA-FLSs treated with Control-CM. Our results explored that knockdown of IHH led to attenuation of BMSCs immunoregulatory ability despite stimulation by TL1A. Consistently, it has been reported that interrupting cellular signaling may decline the MSCs immunomodulatory potency and accelerate cellular senescence. For example, Mateos et al reported that knockdown of lamin A in MSCs mediates impairments in their differentiation potential (48). Also, MSCs osteogenic differentiation was inhibited through beta-catenin deacetylation inhibition after deletion of SIRT1 (49). Besides, FGF2 interruption in MSCs from bone marrow accelerating aging signs such as increased adipogenic differentiation and decreased osteogenic differentiation (50). Taken together, the activity of TL1A on signaling, differentiation, paracrine secretions, migration, and proliferation of BMSCs is maybe regulated through IHH signaling.
Go to :
Conclusion
In summary, TL1A/TNFR2 interaction induced paracrine activity, migration, and proliferation in BMSCs. In addition, TL1A promoted BMSCs osteogenic differentia-tion at the cost of adipogenic differentiation. Moreover, this study suggests a possible role for IHH signaling pathway in anti-inflammatory ability of TL1A-stimulated BMSCs. These findings introduce a novel approach in cellular therapy to enhance the therapeutic potency of MSCs.
Go to :
 XML Download
XML Download