

 PDF
PDF Citation
Citation Print
Print


Introduction
PSMD10, also called Gankyrin (PSMD10Gankyrin), is a well-established proteasome assembly chaperone and an oncoprotein. Initially identified as p28 component (Nas6/PSMD10) of 19S regulatory subunit of the 26S proteasome (1), PSMD10 later gained importance for its tumorigenic potential in many cancers by regulating multiple signaling pathways (2, 3). From a structural perspective PSMD10Gankyrin belongs to ankyrin repeat family proteins, well-known for their role as mediators of protein-protein interactions that regulate numerous physiological processes (4, 5). Most importantly PSMD10Gankyrin acts as a dual-negative regulator of both pRb and P53 tumor suppressor pathways, by modulating their ubiquitin mediated proteasomal degradation via specific protein-protein interactions (6, 7).
The well-established association of PSMD10Gankyrin with various cancers overshadows its role in normal cells. As a 19S assembly chaperone, PSMD10Gankyrin is central to the building of the base complex and aids in its association with the 20S core particles (8). Ability of PSMD10Gankyrin to retain NF-κ B in the cytoplasm (9) and regulate its transcriptional activity at the basal level, may be a critical checkpoint in normal cells. PSMD10Gankyrin enhances the transcriptional activity of β-catenin/TCF3 complex (10) that drives Wnt signaling, a major player in cellular differentiation process. Our recent study reported number of proteins that interact with endogenous PSMD10Gankyrin in mammalian cells (11). PSMD10Gankyrin interactions with Hsp70 and Hsp90 chaperones that help in the turnover of proteins may be critical during heat shock response or ER stress. These limited and yet important interactions bring forth the role of PSMD10Gankyrin in cellular homeostasis.
To obtain deeper insights on the biological role of PSMD10Gankyrin, we generated microarray gene expression profiles by overexpressing PSMD10Gankyrin in HEK293 cells used as a model system in our previous studies. The data were analyzed using a combination of topological investigation of networks, gene-set enrichment and pathway analysis. This strategy led us to a remarkable prediction, namely – the potential involvement of PSMD10Gankyrin in the biogenesis of neurons and neuronal functions. Although such networks can have wider biological implications, this analysis focused our attention on a human neural progenitor cells (hNPCs) model system for differentiation. Here we provide preliminary and yet compelling evidence for the role of PSMD10Gankyrin in neurogenesis. We show that both the mRNA and protein levels of PSMD10Gankyrin increase in differentiated cells compared to the hNPCs. This increase in PSMD10Gankyrin parallel the time-dependent increase in β-tubulin, a marker of the differentiated neurons. Overexpression of PSMD10Gankyrin in hNPCs cell line induces the expression of Ngn1 and alters the ratio of neurons to astrocytes favoring the process of neurogenesis.
Materials and Methods
Plasmids and cloning
PSMD10Gankyrin gene was cloned in pCMV10-3XFlag vector to generate stable cell-line in HEK293 cells. PSMD10Gankyrin with N-term 3XFlag tag was cloned into AgeI and EcoRI sites of pTRIPZ vector under minimal CMV promotor with a KOZAC sequence (CGCCACCATG) replacing the RFP gene from the vector. PAX2 and pMD2G packaging vectors were used for virus production.
Cell culture, generation of stable cells and transduction
HEK293 and HEK293-FT cells were grown in DMEM with 10%FBS supplement, 5% CO2 at 37°C. Stable cells for PSMD10Gankyrin overexpression were generated by transfecting HEK293 cells with pCMV10-3XFlag-PSMD10Gankyrin construct and selected with (800 μg/mL) G418 (Sigma). For PSMD10Gankyrin silencing HEK293 cells were transfected with siRNA using Lipofectamine RNAiMax (Invitrogen) and incubated for 48 hrs. Human Neural progenitor cells (ReNcell VM, purchased from Millipore) were grown on laminin (10 μg/mL: Sigma) coated plates or cover slips in Neural Stem Cells maintenance media (Millipore) supplemented with EGF and FGF (20 ng/mL each: Sigma). hNPCs grow rapidly as a monolayer on laminin with a doubling time of 20~30 hours (Supplementary Fig. S1). For differentiation studies cells were differentiated into astrocytes, oligodendrocytes and neurons (Supplementary Fig. S1B~E) in 10~14 days of incubation period by withdrawing EGF and FGF as per the manufacturers protocol (Millipore). hNPCs were transduced with high titer lentiviral particles along with polybrene for 3XFlag-PSMD10Gankyrin overexpression (More details in Supplementary Methods).
Microarray and data analysis
RNA was isolated from ~1×106 HEK293 stable clone cells overexpressing PSMD10Gankyrin and the Vector Control HEK293 clone by the TRiZol (Invitrogen) method following manufacturers protocol. The isolated RNA was outsourced to Genotypic Technology (Bangalore, India). After their quality control report, cDNA was prepared inhouse and was later analyzed by microarray using the the Agilent platform. The data were analyzed using Agilent expression reference guide. Network reconstruction and extensive topological analysis of networks were carried out using various bioinformatics tools (More details in Supplementary Methods).
Semi quantitative and real-time PCR
mRNA from PSMD10Gankyrin stable cells, control HEK293 cells, hNPCs (UD) and Differentiated (DF) cells were isolated by TRiZol (Invitrogen) reagent and cDNA were prepared using Reverse transcriptase (Invitrogen) PCR. Semi-Q PCR and Real time PCR were carried out for target genes using different PCR primers (Supplementary information).
Western blot and antibodies
PSMD10Gankyrin stable cells, control HEK293 cells, hNPCs (UD) and Differentiated (DF) cell lysates were prepared with NP-40 lysis buffer (50 mM Tris pH7.4, 150 mM NaCl, 1% NP-40, 1 mM DTT and 1× protease inhibitors) and immunoblotting was performed with different antibodies (Suplementary Materials).
Luciferase assay
PSMD10Gankyrin stable cells, control HEK293 cells were transfected with TopFlash and Renilla constructs at a ratio of 10 : 1 respectively using Fugene-HD (Promega). Cells were incubated for 48 hrs post transfection followed by cell lysis using 1× luciferase passive lysis buffer (Dual luciferase reporter assay, Promega). Firefly and renilla luciferase activity were measured using a plate reader (Cytation 5).
Immunofluoroscence
The UD or DF cells were fixed with 4% PFA for 20 min at RT, permeabilized with 0.3% Triton-X100 containing 5% BSA in 1× PBS for 2 h at RT. Primary antibodies and secondary antibodies were duluted with the blocking solution and incubated for 1 h at RT or ON at 4°C. For nuclear staining cells were treated with 1 μg/mL of DAPI for 60 sec. Images were taken by Laser confocal microscope (Zeiss LSM meta 510).
Densitometric and statistical analysis
Densitometric quantification of scanned western blot images was performed using Mac BioPhotonics ImageJ. Statistical analysis was performed using Graph Pad Prism 5. To evaluate the significance of values obtained, unpaired Student’s t test was performed. p<0.05 and p>0.05 are considered as significant and non-significant data respectively. ***represents p-value<0.001 and **represents p-value<0.01.
Results and Discussion
Network and pathway analysis of microarray genes predicted probable role of PSMD10Gankyrin in neuronal differentiation
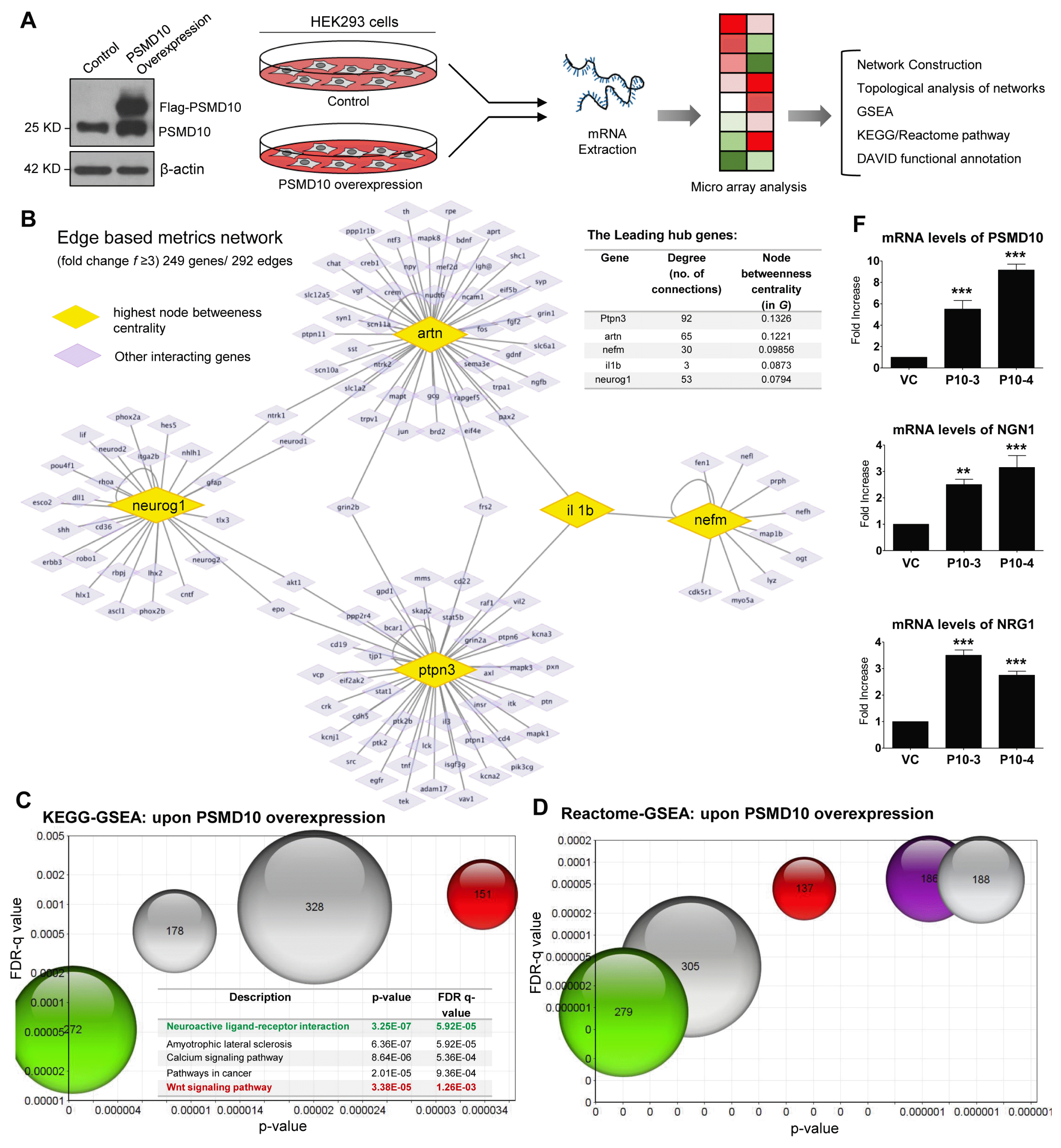
In order to characterize the impact of PSMD10Gankyrin on global gene expression, we carried out a microarray analysis of the total RNA isolated from PSMD10Gankyrin overexpressing HEK293 stable-cells (Fig. 1A) and identified the differentially expressed genes compared to vector control cells. Following an extensive and stringent network reconstruction (Supplementary Methods) we identified 249 genes, 292 edges and 7 hub genes, or the genes whose degree is much higher than the average degree of our network, G (Fig. 1B). The leading genes in G with respective degree mentioned in parenthesis, are: ptpn3 (92), artn (65), neurog1 (53), nefm (30), cebpa (19), galc (9) and tnxb (8). Genes with the highest node betweenness centrality in G are ptpn3 (0.1326), artn (0.1221), nefm (0.09856), il1b (0.0873), neurog1 (0.0794), mapk1 (0.05973), lif (0.0212), akt1 (0.0199) etc. Genes with the highest node closeness centrality in G are: artn (0.2326), il1b (0.2058), bdnf (0.2058), neurog1 (0.1898), ngfb (0.1690). We next turned our attention to edge-based metrics in networks. Edges with the highest values of edge betweenness are (nefm - il1b), (il1b - artn), (mapk1 - nefm), (ptpn3 - mapk1), (lif - artn), (neurog1 - lif) etc. Edges with the highest values of edge proximity (12) consistently have artn, neurog1 and nefm at one of their ends. To extract the biological information from these edges, we used Uniprot to annotate the nodes. This annotation led us to the following biological intuition: the genes and interactions identified as important by various network metrics seem to be involved in neuronal differentiation (Fig. 1B) and therefore we speculated PSMD10Gankyrin could regulate cell fate decisions.
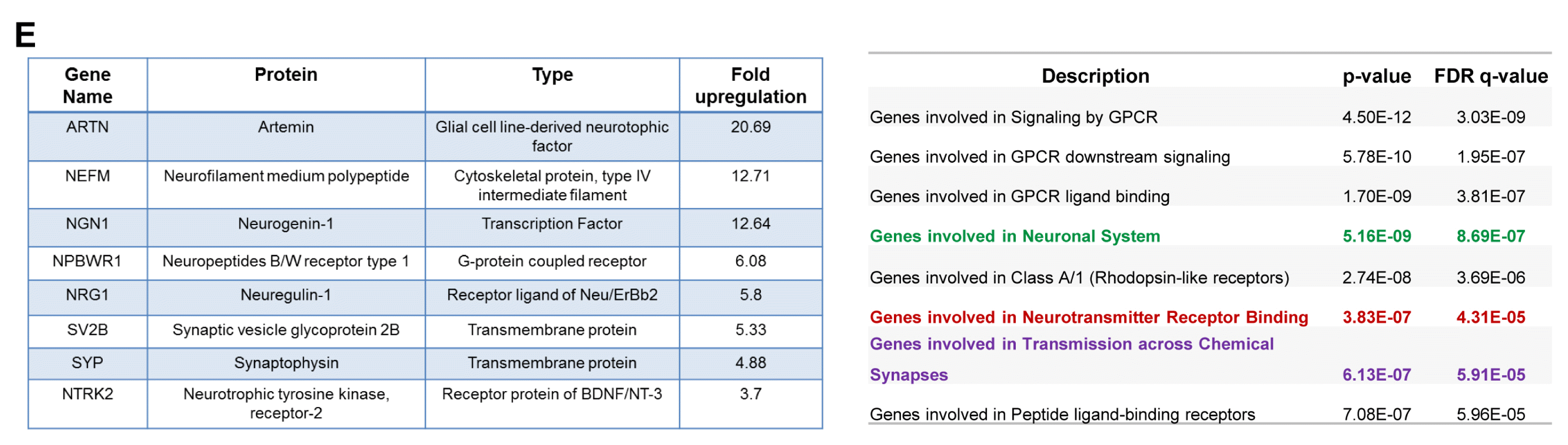
In an independent approach, the cohort of upregulated genes (1.2 log 2 fold changes and above) were analyzed using various web-based bioinformatics tools: GSEA and DAVID functional annotation (13–15). KEGG and Reactome biological pathway analysis sought out statistically significant top hits (with high p-value & FDR q-value) included developmental process, compartmentalization, signaling and functions involving the neurons (Fig. 1C, 1D, Supplementary Table S1, S2). Although less significant by FDR measures but with a p-value of 0.057, one of the hits annotated within DAVID functional analysis was neurogenesis (Supplementary Table S3, S4). Representative genes with the expression values (fold change≥3) and having involvement in neuronal biology are included in Fig. 1E. By Real-time PCR we confirmed mRNA levels of Ngn1 (Neurogenin1), Nrg1 (Neuregulin1) along with PSMD10Gankyrin are higher in PSMD10Gankyrin clones than that of vector control cells (Fig. 1F).
PSMD10Gankyrin levels increase during differentiation of hNPCs
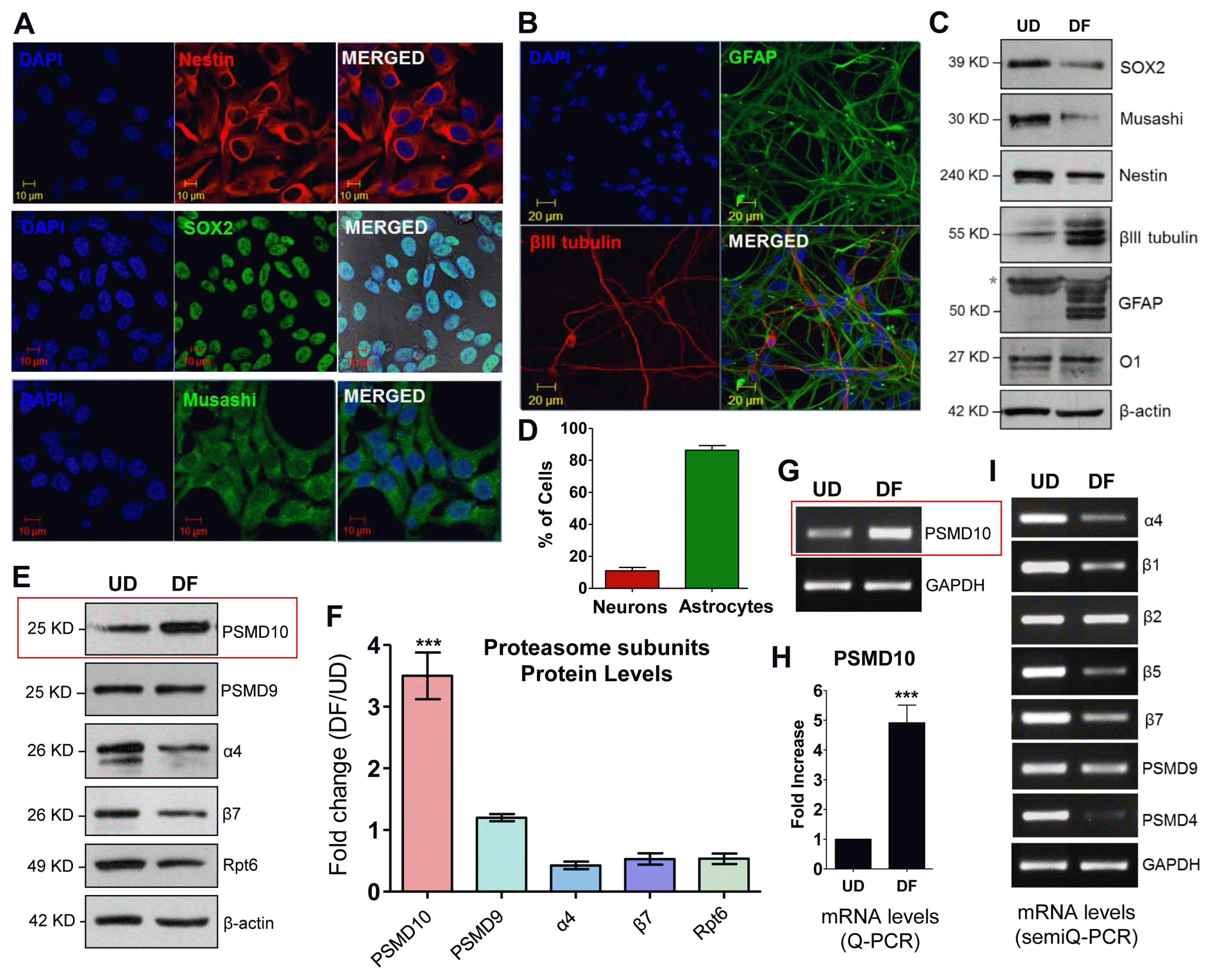
With the above network analyses in place, we infer on the potential role of PSMD10Gankyrin and tested the role of PSMD10 in the differentiation of human neural progenitor cells (hNPCs). First, we checked the differentiating potential of the hNPCs in a monolayer culture (Fig. 2A~C and Supplementary Fig. S2). After 14 days of incubation in media without growth factors (EGF and FGF) the hNPCs differentiated largely to astrocytic cells (~80 to 90%), a small fraction to neurons (~10 to 15%) (Fig. 2D), and the oligodendrocyte cells were sparingly populated (Supplementary Fig. S1A).
We checked PSMD10Gankyrin levels in the undifferentiated progenitor cells (UD) and after 14 days in the terminally differentiated cells (DF). PSMD10Gankyrin protein levels (Fig. 2E, 2F) as well as mRNA levels (Fig. 2G, 2H) are significantly elevated in DF cells as compared to UD cells. In contrast, other proteasome subunits representing each of the sub-complex (α4 and β7 from 20S-CP; Rpt6, an ATPase and PSMD9, a non-ATPase subunit of 19S-RP) either showed a small decrease or remained unaltered in their protein and mRNA levels (Fig. 2E, 2F and 2I). Decreased expression of few proteasome subunits both in protein levels (α4, β7 and Rpt6) and mRNA levels (β1, β5, β7 and PSMD4) may indicate an altered proteasome levels and activity; however, further investigation in the future is needed to understand the significance of these observations.
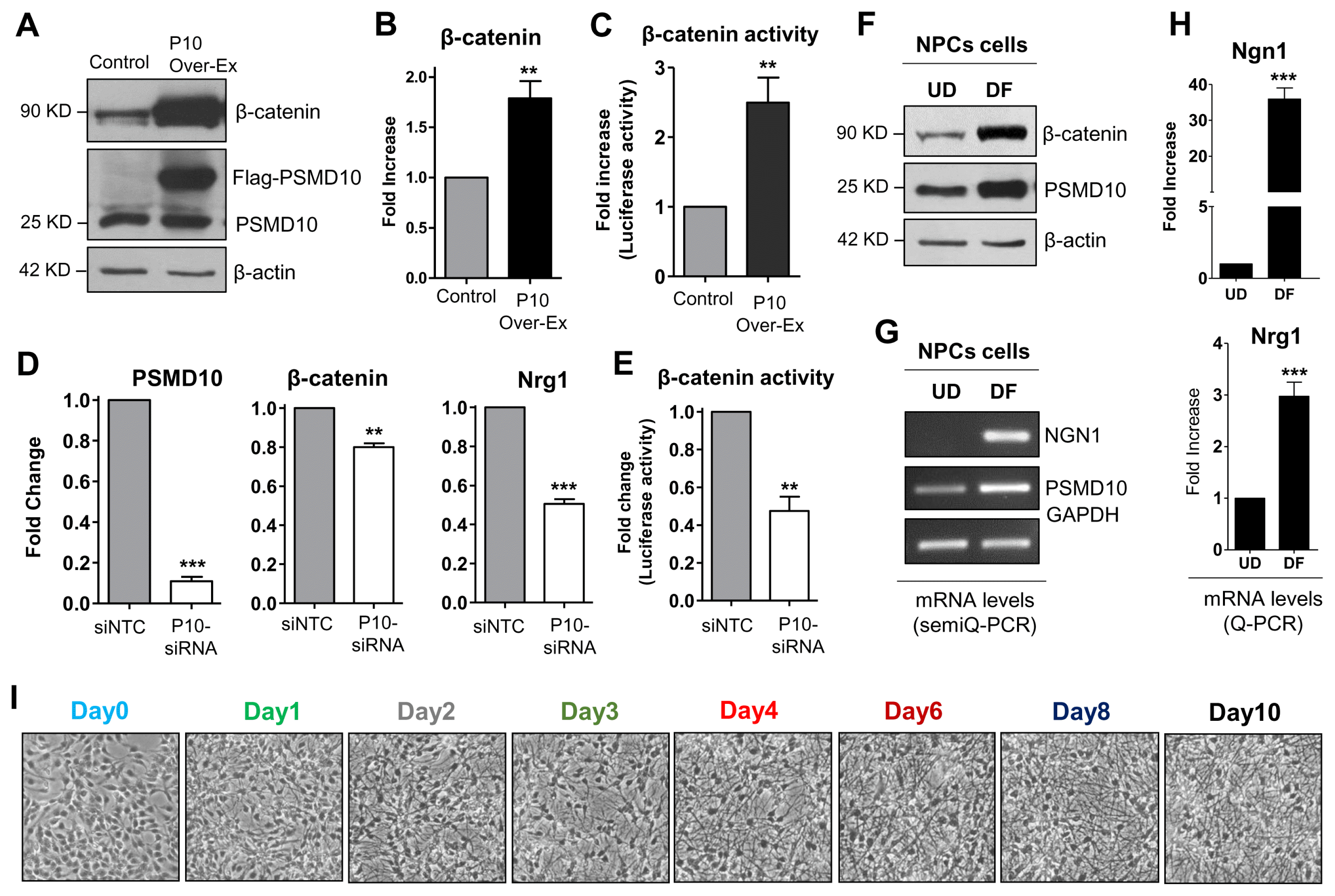
An important outcome of the topological network analyses is that the independent identified nodes are well established for their role in neurogenesis. Among them are Ngn1, a key transcriptional regulator of genes encoding key transcription factors of neurogenesis (16) and β-catenin which directly binds to Ngn1 promotor and regulates its expression during neuronal differentiation of NPC (17). Furthermore, β-catenin and PSMD10Gankyrin mutually regulate expression of each other (10). In support of this, in PSMD10Gankyrin overexpressing HEK293 cells, an increase in β-catenin protein levels (Fig. 3A), mRNA levels (Fig. 3B), and its transcriptional activity (Fig. 3C) were observed. Correspondingly, silencing PSMD10Gankyrin by siRNA reduced the mRNA levels (Fig. 3D) and transcriptional activity (Fig. 3E) of β-catenin. Since hNPCs after differentiation showed higher PSMD10Gankyrin levels, we tested β-catenin levels under undifferentiated (UD) and differentiated conditions (DF). β-catenin protein levels showed a significant increase in the DF cells than in UD cells (Fig. 3F). We also checked the mRNA levels of Ngn1 and Nrg1; both were indeed upregulated in the differentiated populations as compared to undifferentiated hNPCs (Fig. 3G, 3H). Nrg1, one of the upregulated genes in the microarray data, belongs to family of proteins known to induce growth and differentiation of epithelial, glial and neuronal cells (18, 19). Interestingly, Nrg1 was upregulated in PSMD10Gankyrin stable cells (Fig. 1F) and downregulated upon silencing PSMD10Gankyrin in HEK293 cells (Fig. 3D).
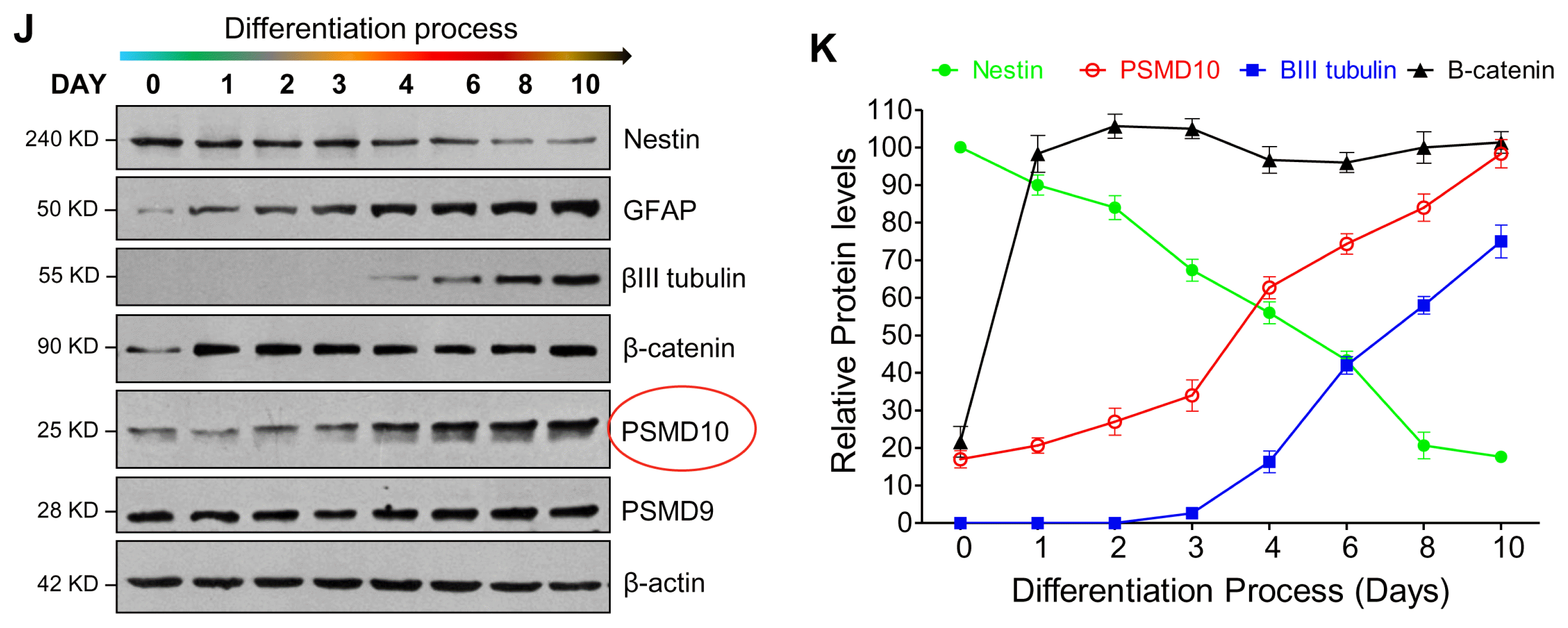
To confirm the observed association between PSMD10Gankyrin levels and cell lineage commitment, we monitored PSMD-10Gankyrin expression pattern during the differentiation process. hNPCs were differentiated for 10 days (Fig. 3I) and cell extracts prepared at different days (total of 8 time points) were analyzed by immunoblotting. Nestin levels steadily decreased indicating that hNPCs are now committed to the process of differentiation (20). The differentiation marker GFAP unique to astrocytes detected on the very first day (Day1) of differentiation and continued to increase until the last day (Day10). The neuronal marker β-III tubulin detected at Day-4, increased gradually until Day-10 of the differentiation process (Fig. 3J, 3K). Concurrently, the levels of PSMD10Gankyrin increased linearly until the end of the differentiation process and paralleled the increase in β-III tubulin during the late stage of differentiation. The quantum increase in PSMD10Gankyrin coincident with the quantum increase in βIII-tubulin suggested a profound role for PSMD10Gankyrin in the differentiation of hNPCs to neurons. In addition, the expression of STAT3 and phosphorylation of STAT3 correlate with the expression of PSMD10Gankyrin during the differentiation process until day 6 (Supplementary Fig. S3A, S3B). However, in the terminally differentiated cells the phosphoSTAT3 levels and mRNA levels of STAT3 (Supplementary Fig. S3C, S3D) went down indicating suppression of STAT3 activity. These results together suggest a probable involvement of PSMD10Gankyrin in glial cell lineage during the initial days of differentiation.
PSMD10Gankyrin overexpression enhances neurogenesis in hNPCs
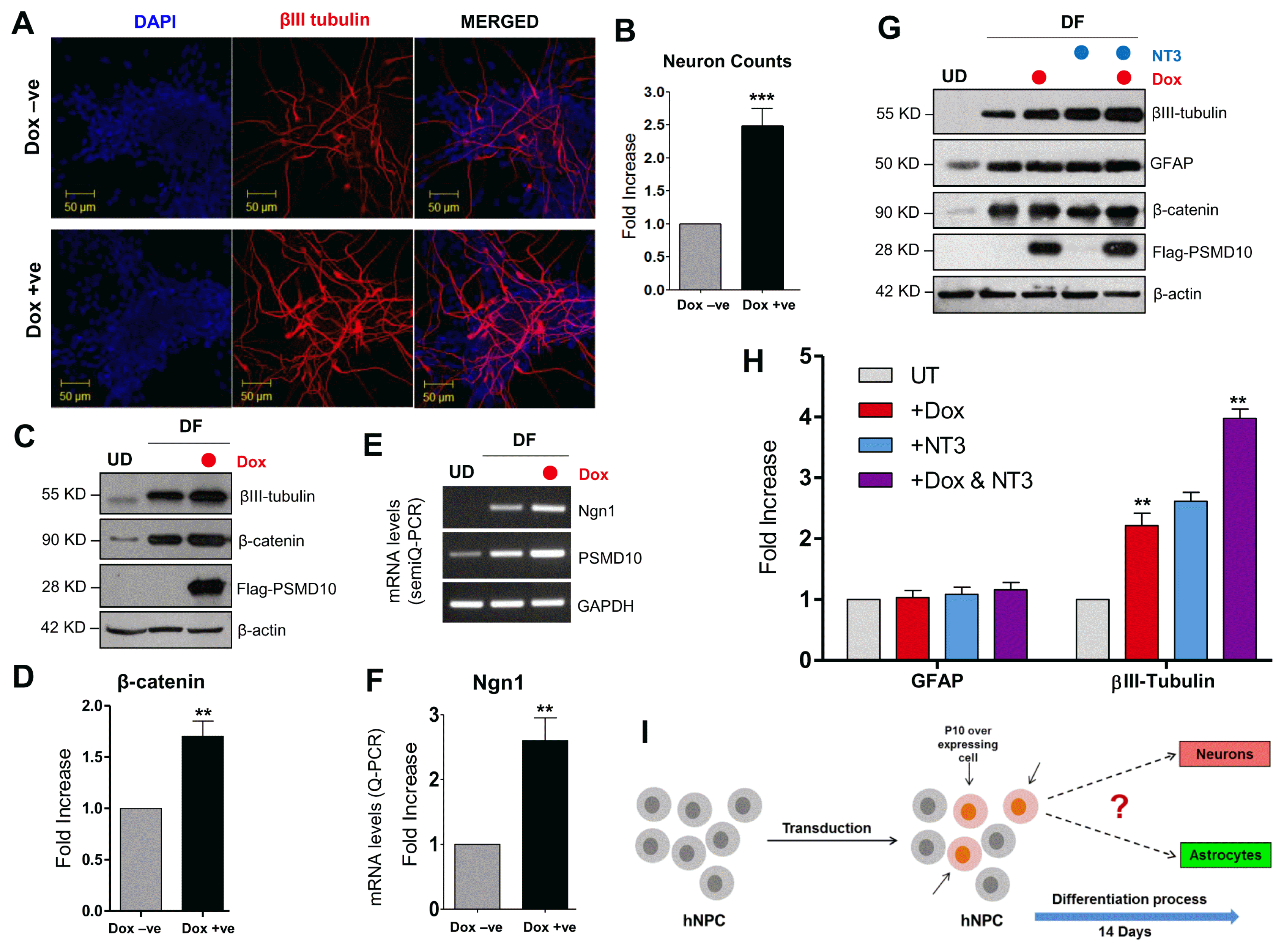
In order to test the aforementioned relationships, we overexpressed doxycycline inducible Flag-PSMD10Gankyrin in hNPCs by viral transduction and checked the induction of β-catenin and Ngn1 and the consequence on cell lineage. Upon Dox-induced PSMD10Gankyrin overexpression in hNPCs and after 12 days differentiation, the neuron count in total differentiated population increased from 10% to ~25% indicating a 2 to 2.5-fold increase in neuronal population (Fig. 4A, 4B). Western blot of the Dox-treated and untreated DF cell lysates showed significant increase in βIII-tubulin (Fig. 4C). Strikingly, both β-catenin and Ngn1 levels increased 2 to 3 fold in the differentiated population when PSMD10Gankyrin was overexpressed (Fig. 4D~F). Furthermore, when PSMD10Gankyrin overexpressing (Dox treated) hNPCs were treated with neurotropin-3 (NT3) they showed higher levels of βIII-tubulin expression than that of the Dox-untreated cells while levels of GFAP remained unaltered (Fig. 4G, 4H). These results are strongly suggestive of the possible functional association between PSMD10Gankyrin and Ngn1 quite likely via β-catenin. PSMD10Gankyrin may be a key player in the network of proteins responsible for neurogenesis.
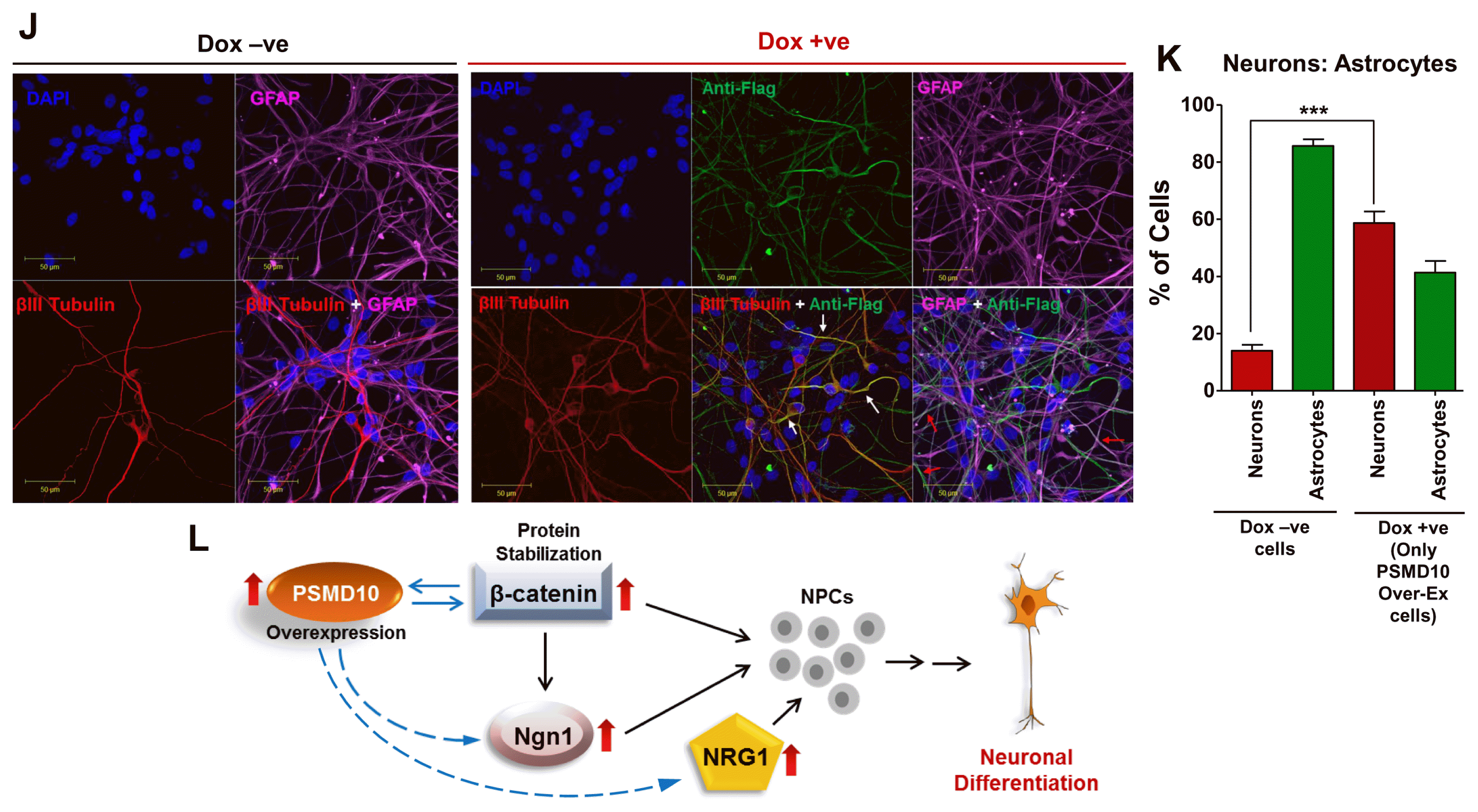
To establish the causal effect of PSMD10Gankyrin in neurogenesis, the hNPCs expressing Flag-PSMD10Gankyrin were analyzed for the composition of the cell types (Fig. 4I). After 12 days of differentiation, immunofluorescence studies indicated that ~60% of Flag-PSMD10Gankyrin expressing cells differentiated into neurons and ~40% Flag-PSMD10Gankyrin expressing cells differentiated into astrocytes (Fig. 4J, 4K). This shift in the ratio of neurons : astrocytes (i.e. from 1 : 9 to 3 : 2) in the untreated cells versus transduced-Dox treated cell population, parallels the overall increase seen in neuron counts upon doxycycline treatment (from 10% to ~25% in Fig. 4A). Significant, but small increase in the overall neuron count may be due to the low transduction efficiency (~20 to 30%, Supplementary Fig. S4). The presence of astrocyte marker in cells that express Flag-PSMD10Gankyrin indicates that PSMD10Gankyrin driven shift in neurogenesis is not absolute which could be due to the following reasons: (1) The current differentiation protocol supports astrocyte formation more than neurons. Therefore, PSMD10Gankyrin overexpression induced signaling events or transcriptional programing may not be enough to execute an exclusive switch to neurogenesis. (2) PSMD10Gankyrin overexpression may trigger “lateral inhibition phenomena” responsible for regulated differentiation of different cell types in a mixed population. (3) PSMD10Gankyrin may favor the formation of both neurons and astrocytes in a spatio-temporal manner. Although less strong than that of βIII-tubulin, correlation between phosphoSTAT3 levels and PSMD10Gankyrin during the early days of differentiation (Supplementary Fig. S3) is suggestive of such a dual role.
The results presented here demonstrate a definite correlation between the levels of PSMD10Gankyrin and the key players such as β-catenin and Ngn1. Microarray analysis of differentially expressed genes induced by PSMD9, a regulator of NF-κB activation (21) did not confer an overwhelming signature for neurogenesis by any of the computational analysis described above (data not shown). In agreement there was no change in the expression of this gene in the course of differentiation of the NPC cells (Fig. 3J). Thus, the gene signature and experimental evidence obtained in the present study is associated specifically with PSMD10Gankyrin. Strengthened by integrated network approach involving topological analysis, gene enrichment and pathway analysis of a microarray data coupled with experimental validation, PSMD10Gankyrin emerges as a new player in cell fate decisions. Here we provide preliminary but compelling evidence for the role of PSMD10Gankyrin in neurogenesis of human neural progenitor cells possibly by stabilizing the β-Catenin and subsequent increase in Ngn1 expression and activity (Fig. 4L).
 XML Download
XML Download