

 PDF
PDF Citation
Citation Print
Print


Introduction
Infertility affects nowadays around 12% of the population worldwide (1). The increased prevalence has been attributed to numerous etiologies such as nutritional behavior, sedentary lifestyle and environmental pollution. Male factor is a major contributor to infertility, being responsible for approximately 50% of these cases, either solely or in combination with a female factor (2).
Several studies support the hypothesis that oxidative stress (OS), defined as an imbalance between reactive oxygen species (ROS) and antioxidant defenses, is strongly associated to male infertility, and may have a subsequent deleterious effect on fertilization, embryonic development, pregnancy and IVF outcomes (3).
Recently, cell therapy has been emerging as a new promising treatment option in the human reproduction field. The immunomodulatory properties of mesenchymal stem cells (MSCs) and their beneficial role in the management and treatment of variable diseases have been already validated by various studies (4). Despite their different origins, all MSCs show plastic adherence, share the following similar phenotypic pattern: CD73+ CD44+ CD90+ CD29+ CD105+/CD45− CD31− CD106− CD34−, and are immunologically inert (5). In addition, studies have recently characterized the antioxidant effect of MSCs, but without clearly defining their mechanism of action (6, 7).
Human MSCs are isolated from bone marrow, adipose tissue (AT), umbilical cord and less frequently from other adult tissues (8, 9). Their potential therapeutic effect is not only due to their differentiation capacity but also to their paracrine and trophic effect; MSCs are reported to secrete various growth factors and cytokines such as interleukin (IL)-6, IL-8, IL-10, insulin-like growth factor 1 (IGF-1), vascular endothelial growth factor (VEGF), hepatocyte growth factor (HGF), interferon gamma, plate-let- derived growth factor, nerve growth factor (NGF) and fibroblast growth factor (FGF) (10).
Due to the numerous advantages of AD-MSCs, mainly their high immune-modulatory effect, great proliferative capability and easy accessibility to body fat (11, 12), the use of their secretome has been widely studied. As a matter of fact, the use of the conditioned medium (CM) has various advantages when compared to the use of stem cells themselves, as CM can be easily obtained, freeze-dried, packed and transported. Moreover, the cellular compatibility between the donor and the recipient is a crucial issue that might lead to therapy failure; hence in order to avoid rejection, CM, devoid of cells, can be the answer (10).
As a promising therapy that warrants further investigation, the biological effects of CM of AD-MSCs on sperm quality were recently evaluated. However, methods of isolation and culture of AD-MSCs, and procedures of preparation of CM were not standardized, thus leading sometimes to inconclusive results (12, 13). Therefore, the aim of this study was to investigate the effect of AD-MSCs-CM on human sperm parameters, where oxidative stress levels and sperm vacuolization are being investigated for the first time.
Materials and Methods
Ethics statement
The study was conducted according to the Declaration of Helsinki and approved by the Ethical Committee of Saint-Joseph University and Hotel-Dieu de France Hospital, Beirut, Lebanon (Ref: HDF 917). Samples (sperm and AT) were collected after obtaining a written informed consent from all participants.
Study population
A total of 30 healthy men attending Azoury IVF Clinic (fertility center) at Mount Lebanon hospital were assessed in this study. All participants had a normal semen analysis based on sperm parameters evaluation. In-person interviews were conducted to complete a questionnaire about age, current or previous disease status including urogenital pathologies, and habits such as smoking and alcohol intake. The mean age of participants was 35±5 years.
Semen samples, obtained by masturbation and collected in sterile plastic containers, were incubated at 37°C for 30 minutes to allow liquefaction. Each sample was then evaluated for sperm concentration, motility, morphology, viability, DNA fragmentation and OS levels. The conventional semen profile for all samples was constructed according to the World Health Organization (WHO) manual for the examination of human semen (14).
Participants with a sperm concentration less than 14×106 cells/ml were not included in the study. All patients presenting with abnormal sperm parameters, suffering from andrological disorders, known to have inflammatory or immune diseases or a recent fever, or undertaking any treatment that may alter spermatogenesis, were excluded from the study.
Preparation of semen samples
Sperm samples were prepared using density gradient technique according to manufacturer’s protocol (Sperm Gradient kit, COOK medical, Australia). Briefly, density gradient media was prepared by layering the 40% medium over the 80% medium, and two ml of the liquefied semen sample were added over the upper layer. After centrifugation (380 g for 10 minutes), most of the supernatant was gently removed and the pellet was resuspended in two ml of sperm medium (Sperm Medium, COOK medical, Australia) to remove the density gradient medium. At the end of the centrifugation (670 g for 5 minutes), the final pellet was resuspended in the sperm medium.
Induction of OS using hydrogen peroxide (H2O2)
The resulting pellets (n=5) were cultured in duplicate in the presence of several concentrations of H2O2 ranging between 0 and 40 μM with different incubation time. Upon establishment of a dose-response curve for the effects of H2O2 on sperm parameters, the suitable concentration and incubation period for the induction of sperm OS were determined. Samples prepared were then left untreated or treated with 10 μM H2O2 for 1 hour (Supplementary Fig. S1).
Isolation and primary culture of AD-MSCs
Adipose tissue samples were obtained from healthy donors undergoing an elective liposuction procedure of the abdominal, hip or thigh areas at the department of plastic surgery, Hôtel-Dieu de France hospital, Beirut, Lebanon. Lipoaspirates were washed extensively with sterile Hank’s buffer (Sigma-Aldrich, USA) to remove red blood cells, and digested with a type I collagenase solution (Sigma-Aldrich, USA) at a concentration of 0.075% in Hank’s buffer for 2~3 hours at 37°C with gentle stirring. The collagenase was then inactivated with an equal volume of DMEM (Dulbecco’s modified eagle’s medium) containing 10% FBS (Sigma-Aldrich, USA), and the mixture was centrifuged for 10 min at 600 g at room temperature. The pellet obtained was resuspended in one volume of Hank’s buffer, and filtered through a 100 μm sieve to remove the debris. After centrifuging the filtrate for 10 min at 600 g, pellet was resuspended in one ml of red blood cell lysis buffer (Sigma-Aldrich, USA) for 1 min, then incubated with 19 ml of Hank’s buffer for 10 min at room temperature. Then, a centrifugation for 7 minutes at 300 g at room temperature was performed to remove lysed red cell debris as well as traces of the lysis buffer. The resulting stromal vascular fraction was plated in cell culture flasks at a density of 10,000 cells/cm2 in DMEM-F12 containing 10% FBS and 1% penicillin-streptomycin-amphotericin (PSA) (Sigma-Aldrich, USA), then incubated at 37°C in a humidified atmosphere containing 5% CO2. After 48 hours, the non-adherent cells were removed and a fresh volume of DMEM-F12 was added and replaced every 2~3 days. Cells adherent to the plastic, and having a fibroblastic morphology were regarded as AT-MSCs. After 21 days, cells were detached by trypsinization, collected, and resuspended in a complete DMEM-F12 medium containing 20% FBS and 1% PS (Sigma-Aldrich, USA). These cells, considered in first-passage (P1), were used in subsequent experiments.
Multi-lineage differentiation capacity of AD-MSCs
AD-MSCs were differentiated into adipocytes, osteocytes and chondrocytes using respectively the Human Mesenchymal Stem Cell (hMSC) Adipogenic Differentiation Medium and hMSC Adipogenic Maintenance Medium (Lonza, Walkersville, MD, USA), the hMSC Osteogenic Differentiation Medium (Lonza, Walkersville, MD, USA), and the hMSC Chondrogenic Differentiation Medium (Lonza, Walkersville, MD, USA) according to manufacturer’s instructions. Adipose cells, osteocytes and chondrocytes were identified using Oil Red O staining, Alizarin Red staining and Alcian Blue staining (Sigma-Aldrich, St. Louis, MO, USA) respectively.
Immunophenotyping of AD-MSCs
AD-MSCs in P1 were characterized by flow cytometric quantification of the surface markers CD73, CD44, CD90, CD29, CD105, CD45, CD31, CD106 and CD34 by specific antibodies (Becton, Dickinson, USA). Cells in culture were detached by trypsin, washed with a DMEM-F12 medium containing 10% FBS and 1% PS, and incubated in the presence of the appropriate monoclonal antibodies in the absence of light for 20 min at 4°C. After incubation, cells were washed twice with cold PBS containing 0.5% bovine serum albumin, centrifuged at 300 g for 5 min, and fixed with a 1% formaldehyde solution. Labeled and unlabeled cells were subsequently analyzed by CellQuest software (Becton Dickinson, USA). A minimum of 10,000 events has been characterized and reported.
Generation of conditioned medium
One million AD-MSCs in P1, cultured in 75 cm2 flasks (Sarstedt, USA) at 37°C using DMEM-F12 medium containing 10% FBS and 1% PS, were used to obtain the CM. At 70% confluence, the media was removed, cells were washed twice with PBS, and re-incubated with serum-free medium. After 24 h, 48 h and 72 h, the CM containing AD-MSCs secretions was collected and centrifuged at 380 g for 10 min to remove cell debris. The CM was stored at −80°C until further use.
Sperm treatment with CM
The effect of CM on sperm parameters was assessed by incubating stressed and non-stressed sperm in 24-well plates in the presence or absence of 100% CM collected after 24 h, 48 h and 72 h of AD-MSCs confluence. Sperm cultured with DMEM-F12 were considered as controls. Sperm parameters were evaluated, after 1, 2, 4, 8, 12, 24 and 48-hour incubation at 37°C.
Measurement of intracellular ROS
Sperm H2O2-induced OS was evaluated through measurement of ROS in sperm cells. ROS status was assessed using nitroblue tetrazolium (NBT) test (Sigma-Aldrich, Japan) as described previously (15). Briefly, NBT staining was done by mixing equal volumes of 0.1% of NBT solution and sample (10 μl) and incubation for 30 minutes at 37°C. A smear was prepared from the mix, then let to dry and finally counter stained with Wright stain solution (Sigma-Aldrich, Japan). A total of 200 sperm cells were evaluated under 100× oil immersion with a bright field microscope. NBT, an electron acceptor, reacts with cellular superoxide ions to form a blue-black compound formazan. ROS positive sperm cells were scored as NBT+ when solid deposits of formazan were found in cytoplasm and/or head.
Sperm motility
Sperm motility was determined under light microscopy using 40× magnification by counting sperm cells in 6 microscopic fields to determine the average percentage, and noted as progressive, non-progressive and immotile, according to WHO 2010 criteria (14). Each sample was evaluated by two technicians, in duplicate, and readings were averaged between the two evaluations after calculation of error.
Sperm DNA fragmentation
Halosperm kit (halotech DNA, Spain) was used as an improved sperm chromatin dispersion test to assess sperm DNA fragmentation (SDF) according to manufacturer’s instructions. A minimum of 300 sperm cells per sample were scored under microscope, using the objective 100. Spermatozoa with large or medium sized halos, characteristic of dispersed DNA loops, were considered to have intact DNA. Sperm cells with small, degraded or absent halos were considered to have fragmented DNA. SDF was determined for each specimen, using the following formula: SDF (%)=100×(number of spermatozoa with fragmented DNA)/(number of spermatozoa counted).
Sperm morphology and vacuolization
Sperm morphology and vacuolization were evaluated using the Spermoscan Kit (RAL diagnostics, Australia) according to manufacturer’s instruction. Briefly, the smears were fixed in the spermo FIX-RAL solution. The sperm cytoplasm and nucleus were then stained using the spermo EOSIN-RAL and the spermo BLUE-RAL respectively. After drying, a minimum of 200 sperm were counted and assessed for morphology and vacuolization using the optic microscope according to Kruger’s strict criteria: head defects: large or small, tapered, amorphous, vacuolated (more than two vacuoles or >20% of the head occupied by unstained vacuolar areas), small or large acrosomal areas; mid-piece defects: bent, cytoplasmic residues; tail defects: coiled, multiple (14).
Sperm viability
Sperm viability was evaluated using eosin-nigrosin staining. Ten microliters of each sample was mixed with equal volume of eosin-nigrosin solution. After 30 seconds, the samples were smeared and let to air dry. At least 200 sperm were evaluated under 100× oil immersion with a bright field microscope. Unstained sperm were classified as alive, whereas dead ones with altered membranes were stained pink or red (eosin) with a dark background (high concentration of nigrosin) (14).
Statistical analysis
Means and standard deviations were calculated for all variables using the GraphPad Prism software version 6 (GraphPad Software, Inc., USA). The analysis of variance (ANOVA) for nonparametric data was used to assess the variables between groups, followed by Tukey’s post hoc test. p-values less than 0.05 were considered statistically significant.
Results
Immunophenotyping of AD-MSCs
Flow cytometry was used to characterize MSCs isolated from adipose tissues. Analysis of results showed that these cells expressed the specific surface markers of mesenchymal stem cells, namely CD73, CD29, CD44, CD90 and CD105, and lacked the expression of endothelial and hematopoietic cell markers CD34, CD31, CD45 and CD106 (Fig. 1).
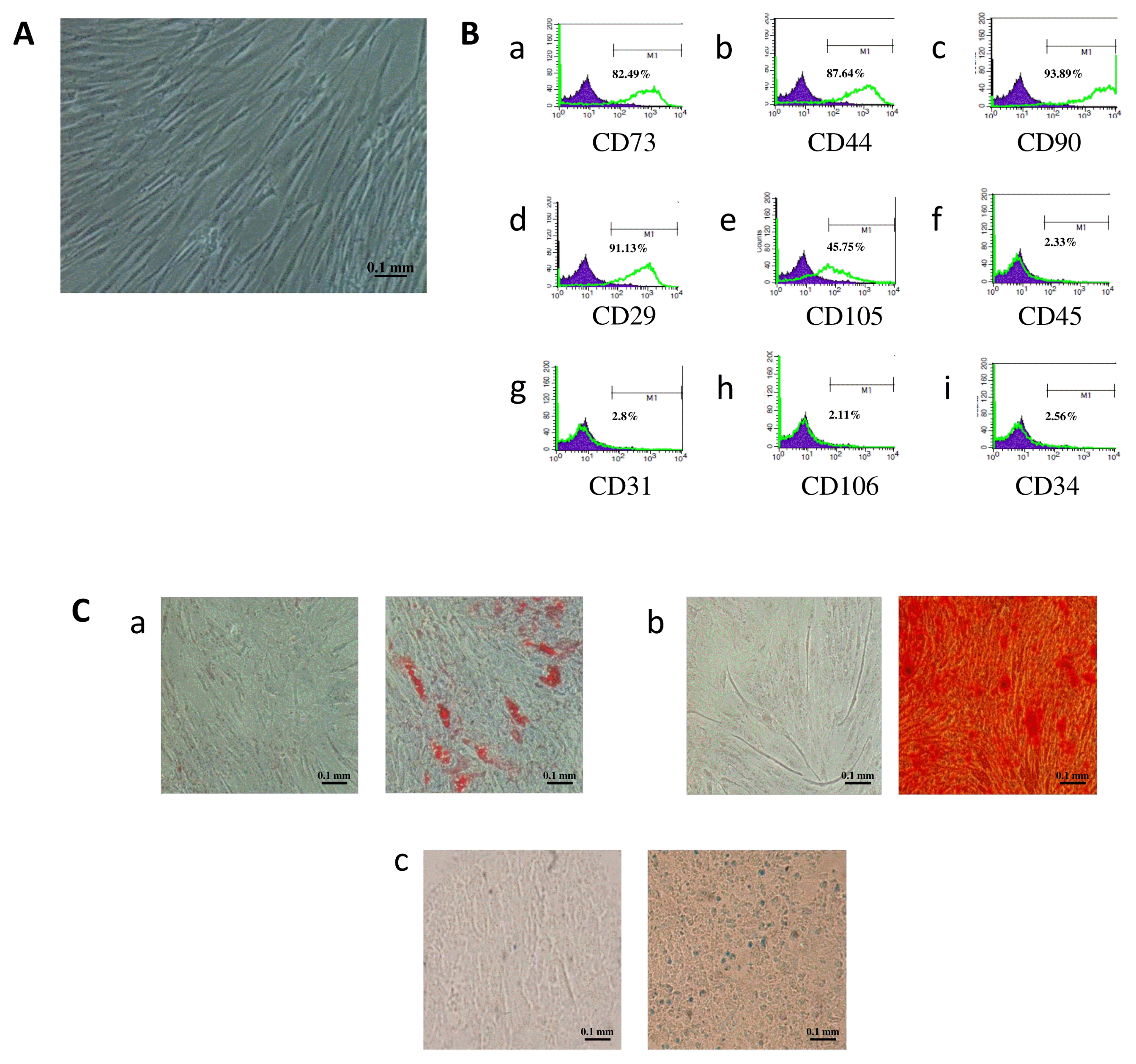
Fig. 1
Immunophenotype and differentiation capacity of human adipose-derived mesenchymal stem cells (AD-MSCs). (A) AD-MSCs have fibroblastic features at passage 1 (P1) (magnification 100×). (B) The flow cytometry histograms for a representative donor are displayed at P1. The percentage of cells stained positive is indicated in the upper right corner of each panel. The green line indicates the positively stained cells, whereas the purple line indicates the isotype-matched monoclonal antibody control. (C) Cells were incubated for 21 days in adipogenic, osteogenic and chondrogenic media. Representative images of AD-MSCs differentiation into (a) adipocytes identified by the presence of intracellular lipid droplets that were confirmed by Oil-Red-Staining, (b) osteocytes characterized by the formation of calcium deposits stained by red alizarin, and (c) chondrocytes which presence was revealed by Acian Blue staining. Scale bar=100 μm.
![]()
Multilineage differentiation capacity of AD-MSCs
Undifferentiated AD-MSCs have a fibroblast-like morphology. To test their multilineage differentiation capacity, cells were incubated for 21 days in adipogenic, osteogenic and chondrogenic media. The formation of intra-cellular lipid droplets revealed the differentiation of AD-MSCs into adipocytes, which was further confirmed by the presence of a red orange color using “Oil Red O” staining. The presence of calcium deposits, characterizing AD-MSCs differentiation into osteocytes, was visualized by red alizarin stain. AD-MSCs differentiated into chondrocyte-like cells were identified by the development of a blue color using Alcian Blue staining (Fig. 1).
Establishment of the optimum incubation period for sperm parameters improvement
The optimum incubation period to improve sperm quality and function was determined by culturing sperm cells for two days with or without AD-MSCs-CM collected at 24, 48 and 72 h of conditioning, and assessing sperm parameters at different time points of culture (1 h, 2 h, 4 h, 8 h, 12 h, 24 h and 48 h). Results revealed that the total motility (%) at 24 h was still stable in stressed and non-stressed sperm treated with AD-MSCs-CM compared to the starting point (T=0 h) while it decreased significantly and in a steady fashion in the control group of sperm cultured with DMEM-F12 medium as of the starting point at T=0 h, hence showing the ability of AD-MSCs-CM to maintain the motility for 24 h before dropping thereafter (Fig. 2A, 2B). Regarding dead sperm, the percentage started to increase progressively in all conditions after 1 h of culture, with levels reaching their maximum at 48 h incubation time. However, in both stressed and non-stressed sperm treated with CM, the percentage of dead sperm was always lower when compared to the control group at all-time points, showing the ability of the CM to withhold sperm death and keep cell viability. Moreover, it is important to note that the turning point at which the curve of dead sperm starts increasing rapidly appears after 24 h incubation time in both group (Fig. 2C, 2D). As for sperm vacuolization (Fig. 2E, 2F), sperm DNA fragmentation (Fig. 2G, 2H) and ROS positive cells (Fig. 2I, 2J), the most significant improvement was noted at 24 h in comparison to other culture conditions and all other incubation timings. Finally, stressed and non-stressed sperm showed no difference in morphology at all-time points of culture, and irrespective of culture conditions (Fig. 2K, 2L). Taking all tested parameters into consideration, the 24 h incubation time was established for having the best effect on sperm vacuolization, DNA fragmentation and oxidative stress while motility and viability were preserved, and thus regarded in our study as the optimum period to study sperm parameters.
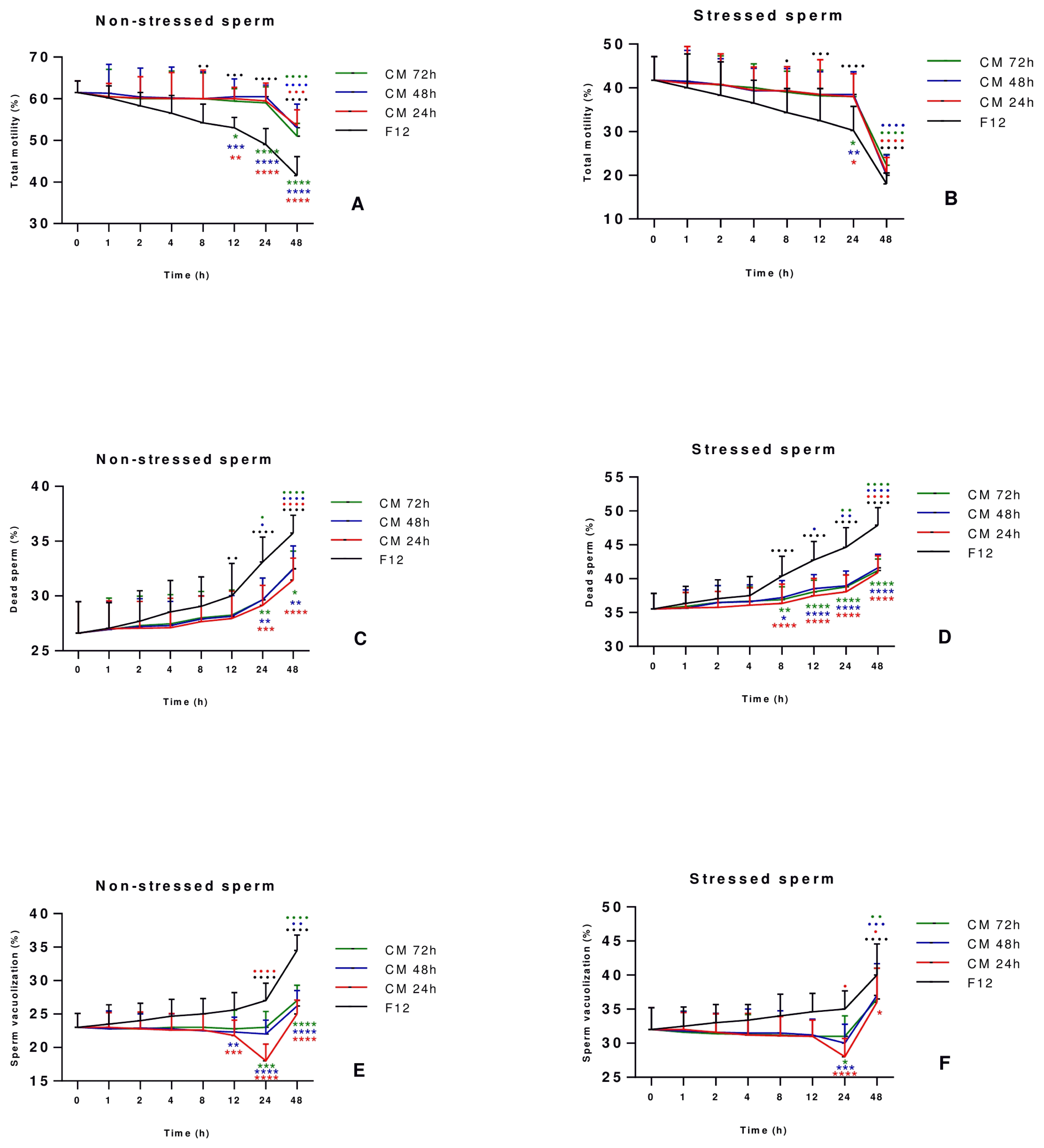
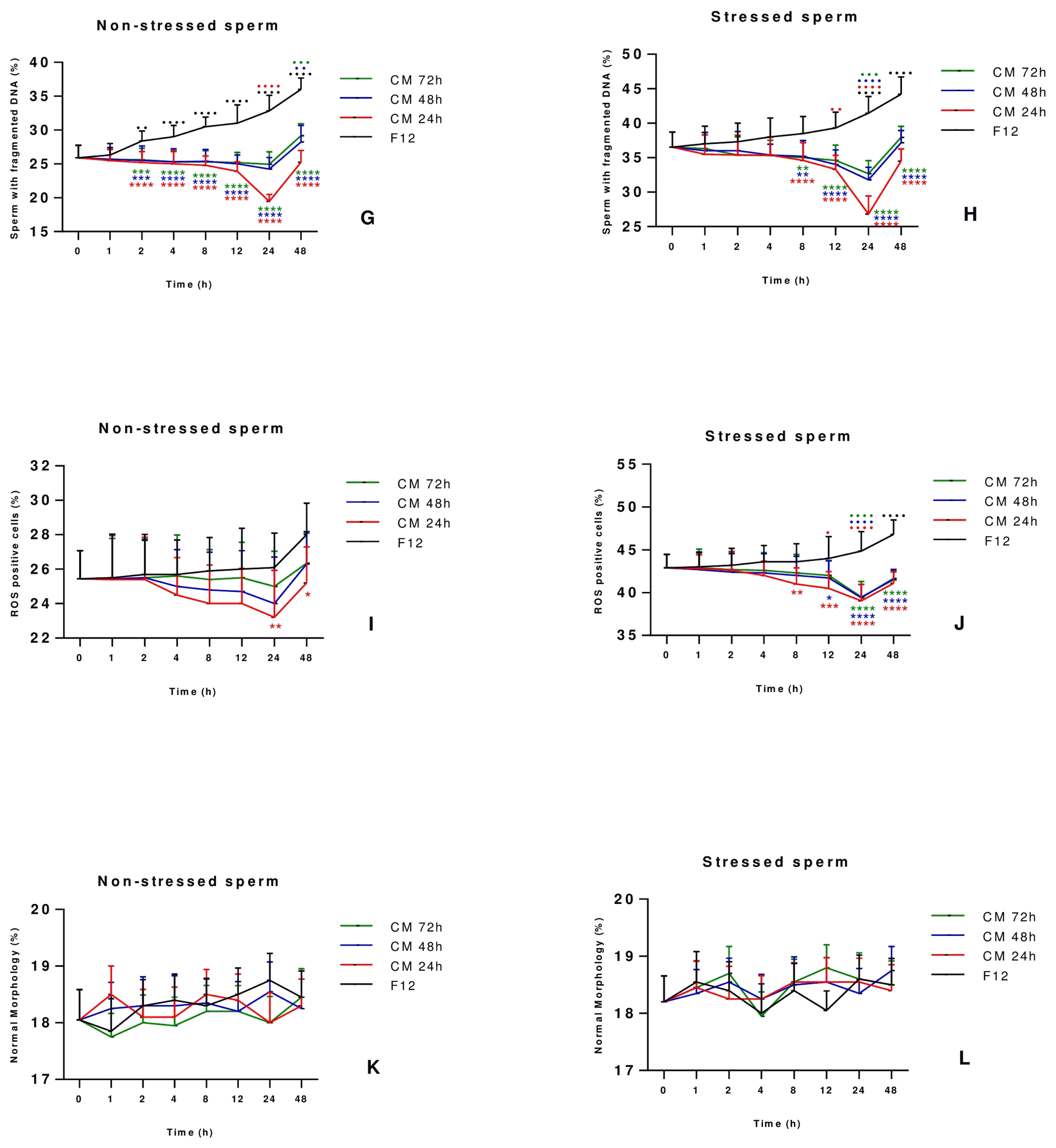
Fig. 2
Establishment of the optimum incubation period for sperm parameters improvement. Sperm collected from 30 healthy individuals were incubated for 1 hour in the presence or absence of 10 μM H2O2. After removal of H2O2, stressed and non-stressed sperm were treated with CM collected at 24, 48 and 72 h of conditioning for different time periods (1 h, 2 h, 4 h, 8 h, 12 h, 24 h and 48 h). Sperm motility (A, B), vitality (C, D), vacuolization (E, F), DNA fragmentation (G, H), ROS levels (I, J), and morphology (K, L) were assessed and compared to those of sperm cultured with DMEM-F12 medium, considered as the control group. Data are represented as mean±SD. Differences among groups were evaluated using the non-parametric Two Way ANOVA test further evaluated by Tukey test for post hoc pairwise comparisons. *p<0.05; **p<0.01; ***p<0.001; ****p<0.0001 for significance between sperm treated with CM 24, 48 or 72 hours and the control group (F12) at a same time-point. ●p<0.05; ●●p<0.01; ●●●p<0.001; ●●●●p<0.0001 represent differences in results observed in sperm treated with CM for different incubation periods as compared to sperm immediately analyzed after collection (0 h incubation time).
![]()
![]()
Study of the temporal effect of CM on sperm parameters
In order to determine how the temporal profiles of CM collection affect sperm quality, stressed and non-stressed sperm were cultured with DMEM-F12 medium or AD-MSCs-CM collected at 24, 48 and 72 h, and parameters were compared after 24 hours of incubation time, defined previously as the optimum period of incubation. Interestingly, incubation with CM significantly improved motility (Fig. 3A) and vitality (Fig. 3B) of stressed and non-stressed sperm for all the time points, but without noticing any difference in the impact with regards to the time of CM collection. Furthermore, a significant decrease in sperm DNA fragmentation (Fig. 3C), sperm vacuolization (Fig. 3D) and ROS positive cells (Fig. 3E) was detected in stressed and non-stressed sperm treated with CM compared to controls. Interestingly, both non-stressed and stressed sperm showed better DNA fragmentation and vacuolization rates when treated with CM 24 h as compared to CM 48 or 72 h; the percentage of ROS positive cells being on the other hand comparable in all CM treated groups, irrespectively of the collection time. Finally, no significant difference in sperm morphology was observed between the treated and control groups, hence indicating no effect of CM on the appearance of the spermatozoa (Fig. 3F).
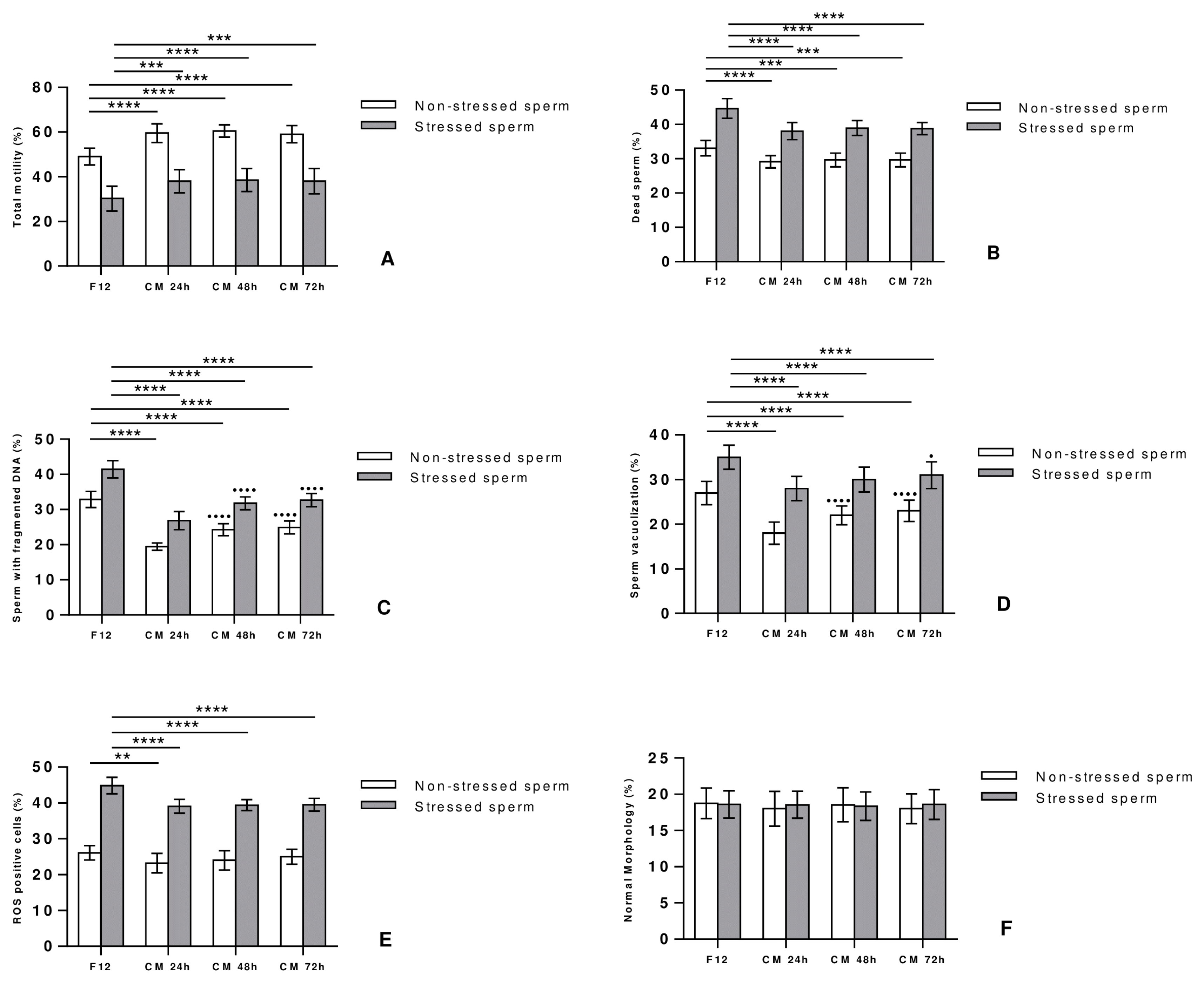
Fig. 3
Temporal effect of conditioned medium on human sperm motility (A), vitality (B), vacuolization (C), DNA fragmentation (D), ROS levels (E), and morphology (F). Sperm collected from 30 healthy individuals were incubated for 1 hour in the presence or absence of 10 μM H2O2. After removal of H2O2, stressed and non-stressed sperm were incubated in the presence or absence of CM collected at 24, 48 and 72 hours. After 24-hour incubation at 37°C, sperm parameters were assessed and compared to the control group (sperm cultured with DMEM-F12 medium). Differences among groups were evaluated using the non-parametric One Way ANOVA test further tested by Tukey test for post hoc pairwise comparisons. Data are represented as mean±SD. **p<0.01; ***p<0.001; ****p<0.0001 for differences between CM-treated groups and F12. ●p<0.05; ●●●●p<0.0001 for differences between sperm treated with CM 48 h or CM 72 h and sperm treated with CM 24 h.
![]()
Effect of CM 24 h treatment on fresh sperm (T0)
After establishing the CM 24 h as the most suitable condition to improve sperm quality and function, parameters were compared to those of fresh semen (T0) usually collected immediately after processing in couples undergoing fertility treatment.
Stressed and non-stressed sperm showed comparable motility (Fig. 4A) and vitality (Fig. 4B) profiles in CM-treated and fresh sperm, thus pointing out the ability of CM to relatively maintain these two parameters over time for about 24 hours. In contrast, CM treatment significantly reduced the percentage of sperm DNA fragmentation (Fig. 4C), sperm vacuolization (Fig. 4D) and ROS positive cells (Fig. 4E) in comparison to the non-treated fresh sperm. These results show that infertile patients with a high degree of sperm vacuolization and DNA fragmentation can benefit from a 24-hour CM treatment of their fresh semen while keeping the other parameters stable, which can improve sperm quality and subsequently IVF outcome.
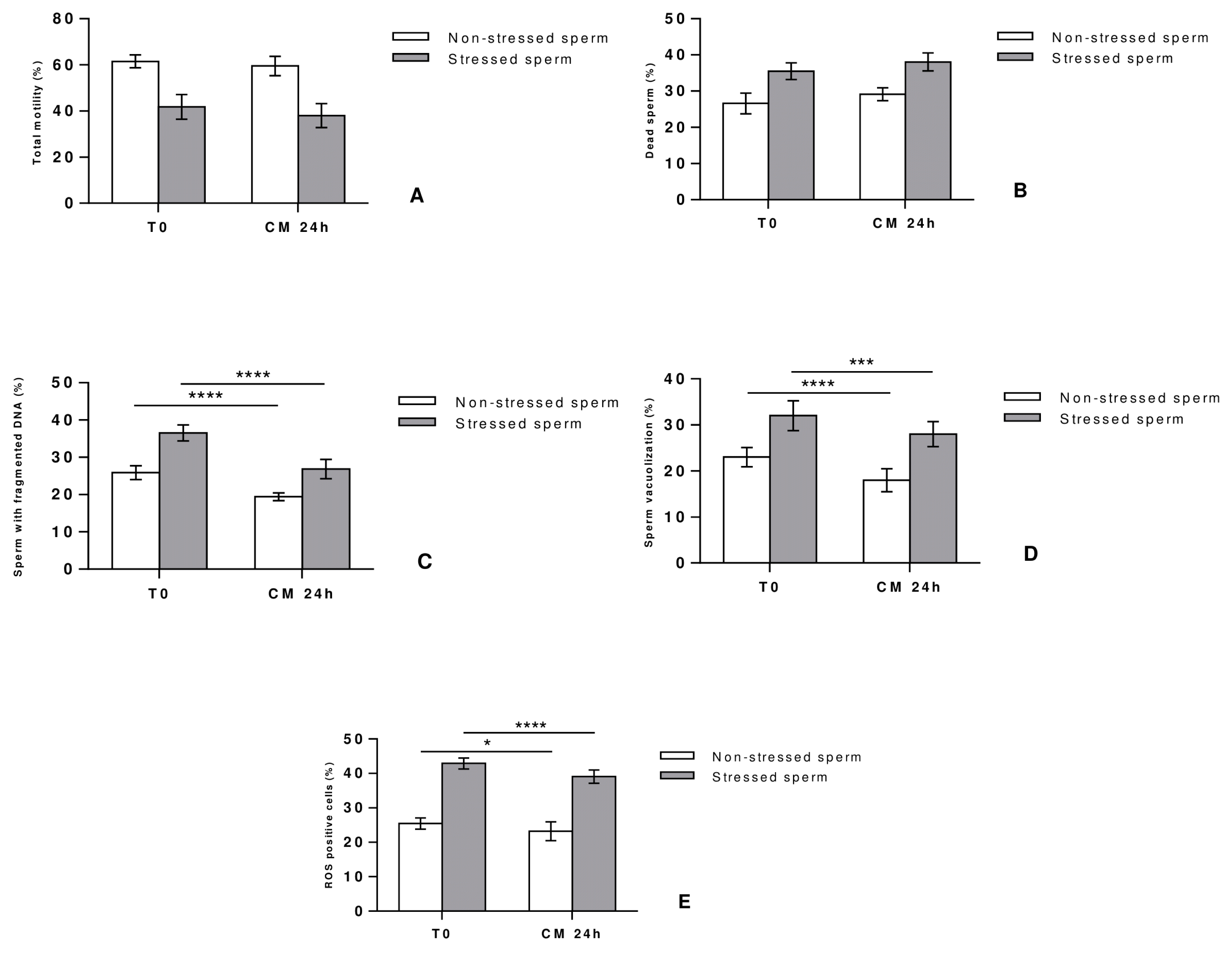
Fig. 4
Effect of CM 24 h on human sperm motility (A), vitality (B), DNA fragmentation (C), vacuolization (D), and ROS levels (E). Parameters of stressed and non-stressed samples treated with CM 24 h were assessed and compared to those of fresh semen immediately tested after collection. Differences among groups were evaluated using the non-parametric One Way ANOVA test further tested by Tukey test for post hoc pairwise comparisons. Data are represented as mean±SD. *p<0.05; ***p<0.001; ****p<0.0001.
![]()
Discussion
OS has been identified as one of the main contributory factors to male infertility, thus affecting fertilization, pregnancy and live birth rates (3). Oral antioxidants (16) and testicular sperm (17) were suggested among possible approaches to overcome OS damage. While testicular sperm is considered to be an invasive procedure, disadvantages of using oral antioxidants include their cost, the lack of agreement on the dose, the duration of treatment, and whether individual or combined oral antioxidants should be used (18). Therefore, the search for a quick, affordable, safe and non-invasive procedure to improve sperm quality can be a game changer in fertility treatments.
Cell therapy using stem cells has been lately shown to have therapeutic effects in many diseases such as spinal cord injury (19) multiple sclerosis (20), as well as in in vivo model of infertile male rats (21). However, it has been reported that the beneficial effect of MSCs is not due to their differentiation capacity but to their paracrine and trophic effects, thus referring to stem cells as “the medicinal signaling cells” (22), a more accurate term reflecting their immunomodulatory and regenerative effect.
In this regard, the idea of the present study was to create a co-culture system in which sperm cells are incubated with AD-MSCs secretome containing a cocktail of growth factors and cytokines. In fact, such a culture system could reduce adverse effects that may result from sperm preparation techniques (23), and hence might represent a new promising approach to restore sperm cells damage. On the other hand, the beneficial effect of secretome was reported in various degenerative diseases such as skin wound healing (24) and muscle degeneration (25). Moreover, the use of CM rather than MSCs directly was based on the easy manipulation, storage and packing of CM, unlike stem cells that are vulnerable to many environmental conditions, making the latter unpractical as a treatment option. MSCs were successfully isolated from human adult adipose tissues, characterized, and subjected to several culture conditions to optimize on one hand the temporal effect of CM, defined as the time of collection of AD-MSCs secretome, and on the other hand the best incubation period required to improve sperm quality parameters.
In agreement with previous data, we observed in the present study a beneficial effect of AD-MSCs secretome on sperm parameters. Our results have shown that 24-hour incubation of stressed and non-stressed human sperm with AD-MSCs-CM collected at 24 hours of conditioning reduce sperm vacuolization and DNA fragmentation rates while preserving motility and viability. Moreover, the most significant result was noted with CM collected after 24 hours of incubation, in comparison to 48 and 72 hours, with comparable results noted in both stressed and non-stressed sperm. In line with these results, a study conducted by Prihatno et al. (26) showed that high-dose secretome derived from the human fetal umbilical cord was able to increase the number and motility of sperm cells in rats with cisplatin-induced testicular dysfunction, and to promote the regeneration of the seminiferous tubules. On the other hand, Mokarizadeh et al. (27) revealed that stem cells secretome can improve viability, motility and antioxidant capacity parameters of cryopreserved adult male Wistar rats sperm.
The significant improvement in sperm parameters may be attributed to the numerous secreted bioactive factors mainly growth factors VEGF, granulocyte-colony stimulating factor (G-CSF), IGF-1, HGF, NGF, and FGF (13), anti- inflammatory cytokines IL-10 and transforming Growth Factor-β (TGF-β) (13), and antioxidant defense constituents, more particularly superoxide dismutase (SOD) and catalase (20). Actually it was reported that VEGF, HGF, FGF2, NGF and IGF-1 were capable of maintaining and improving sperm motility by stabilizing sperm membrane proteins (28–31). In addition, NGF was proven to decrease sperm DNA fragmentation in both normal spermatozoa (32) and asthenozoospermic samples (33), whereas G-CSF was found to have a protective effect on sperm by decreasing DNA damage and oxidative stress (34). On the other hand, anti-inflammatory cytokines IL-10 and TGF-β were found to play an important role in human spermatogenesis (35, 36). In this regard, various studies have investigated the relationship between levels of these cytokine and sperm parameters, but results were variable and sometimes contradictory (37, 38). Further studies assessing the role of cytokines in testicular function and maintenance of male reproductive, and their contribution to infertility disorders may be of great interest. Finally, the antioxidant activity of MSCs was confirmed in previous studies in vitro and in vivo through the secretion of protective enzymes SOD and catalase (20, 39), hence improving sperm motility and reducing OS and DNA fragmentation levels via inhibition of lipid peroxidation of sperm cells (40).
In conclusion, the CM of AD-MSCs seems to improve sperm parameters in a concentration- and time-dependent manner. To the best to our knowledge, this is the first study to explore the effect of the CM derived from AD-MSCs on ROS levels and sperm vacuolization. The positive effect of AD-MSCs may be attributed to the wide spectrum of the factors found in the CM on sperm parameters. Further studies are needed to determine the molecular mechanisms underlying AD-MSCs effect on sperm physiology, and would provide new insights for the use of this approach in assisted reproductive technologies.
 XML Download
XML Download