

 PDF
PDF Citation
Citation Print
Print


Introduction
Most of the colorectal cancer (CRC)-related deaths are caused by recurrence and metastasis of CRC and its chemotherapy resistance (1). Oxaliplatin is a third-generation platinum compound widely used for the treatment of CRC in clinical studies (2). When oxaliplatin combined with 5-flourouracil and folinic acid for the treatment of metastatic CRC, the median progression-free survival of CRC patients prolonged 2 years (3). Despite this improvement, all metastatic CRC finally developed to oxaliplatin resistance and patients succumbed to their disease (4). Therefore, identification of the underlying molecular mechanisms of CRC chemotherapy resistance is worthwhile for its therapy and prevention strategies.
Cancer stem cells (CSCs) have been identified to play a crucial role in a variety of cancers including breast cancer, ovarian, melanoma and CRC (5, 6). CSCs are naturally resistant to chemotherapy due to their ability for DNA repair and multidrug resistance ABC transporters expression (7). Those CSCs confer drug resistance to their progeny, resulting in the tumors eventually become less sensitive to the following chemotherapy (7, 8). An important strategy to overcome drug resistance is to regulate CSCs stemness and thereby to increase the sensitivities of the tumor to chemotherapy.
It has been identified that long non-coding RNA (lncRNA) expression pattern is highly correlated with cancer development and prognosis (9, 10). More recently, some studies have revealed that lncRNAs are capable of regulation of CSCs stemness via different molecular mechanisms (11, 12). It is known that CSCs stemness is a major contributor to cancer chemotherapy resistance (7). Therefore, identification of novel lncRNAs which function as the regulators for CSCs stemness might provide evidence for the development of strategies to overcome cancer chemotherapy resistance. Additionally, abnormal expressions of LINC00525 have been identified in the development of many types of cancer including bladder cancer (13), lung cancer (14) and leukemia (15). However, the roles of LINC00525 in the occurrence and development of CRC are still unknown. Thus, in this study, for the first time, we investigated the relationships between LINC00525 and CRC.
Materials and Methods
Clinical specimens and cell lines
Fresh CRC specimens and normal intestine tissue samples were collected from Yidu Central Hospital of Weifang City between 2017 to 2018. In addition, in Fresh CRC specimens, twenty-eight samples were collected from oxaliplatin sensitive patients and thirty-two samples were collected from oxaliplatin resistant patients. The study was approved by the ethics commitment of Yidu Central Hospital of Weifang City, and followed the ethical guidelines for medical research on Human subjects, the Helsinki Declaration of 1975, as revised in 2000.
Five human CRC cell lines including Caco2, SW480, SW620, HCT116 and HT29 and normal human intestinal epithelial cell line (HIEC) were purchased from the Cell Bank of the Chinese Academic of Sciences (Shanghai, China). The cells were cultured in recommended medium supplemented with 10% fetal bovine serum (FBS) (Life Technologies, Gaithersburg, MD, USA) and 100 U/ml penicillin and 100 μg/ml streptomycin (Beyotime, Shanghai, PR China) at 37°C in the presence of 5% CO2 at constant humidity.
Tumorsphere formation assay
Tumorsphere medium used in this study consists of Dulbecco’s Modified Eagle Medium (DMEM)/F12, epidermal growth factor (20 ng/ml), insulin (5 μg/ml), 2% B-27 and 0.4% FBS. Tumorsphere formation assay was performed according to the previously reported method (16). In brief, the HCT116 and HT29 cells were harvested and suspended in phosphate-buffered saline (PBS). Next, dissociated cells were then seeded in tumorsphere medium. The spheres were observed under an inverted microscope.
Cell viability and apoptosis assay
MTT and colony formation were performed according to previously reported methods (17, 18). In brief, MTT solution was added into each well and the microplate was incubated at 37°C for 4 h. DMSO solution was then added and the optical density was read at 570 nm using a microplate reader (Molecular Devices, Sunnyvale, CA, United States). For colony formation assay, HCT116 and HT29 cells were seeded in 6-well plate and grown for indicated days. Next, the colonies were fixed with 4% paraformaldehyde and then stained with crystal violet (0.05% w/v) overnight. The numbers of colonies were counted using Image J.
DNA fragmentation ELISA kit (Roche, Indianapolis, IN, United States) was used to evaluate cell apoptosis, according to the manufacturer’s instructions.
Construction of stable cell lines
To construct LINC00525 knockdown cell lines, the cells were transfected with lentiviruses expressing either LINC00525-1 shRNA or LINC00525-2 shRNA. The shRNA sequences are in Supplementary Table S1. Twenty-four hours after transfection, total RNA was isolated and expressions of LINC00525 were analyzed by qPCR.
To construct LINC00525 and ELK3 overexpression cell lines, LINC00525 or ELK3 was amplified and subcloned into p-Sin 3×flag vector. The cells were transfected with p-Sin 3×flag vector containing LINC00525 or ELK3 and followed by the selection of antibiotics. Primers for amplification of LINC00525 and ELK3 are shown in Supplementary Table S1. Expressions of LINC00525 and ELK3 were determined by either qPCR or western blotting, to ensure the stable cell lines were successfully established.
Biotin pull-down assay
Biotin pull-down assay was performed under RNase free conditions according to previously reported methods (19). Sense and anti-sense biotin-labeled DNA oligomers were used. Three pairs of LINC00525-DNA oligomers were used in this study. The sequences are shown in Supplementary Table S1. Biotin-labeled DNA oligomers were incubated with cell lysates for 1 h. Next, agarose beads conjugated with streptavidin were added to isolate RNA-RNA complex.
Luciferase assay
Dual-luciferase® reporter assay system was established according to the manufacturer’s instructions (Promega, Madison, MA, United States). LINC00525 wild-type (LINC00525 wt) and the miR-507-binding-site mutated (LINC00525 mt) one was constructed and then integrated into dual luciferase vector. After that, the cells were co-transfected with dual luciferase vector and miR-507 or miR-NC mimics.
Similarly, dual luciferase vector containing wild-type ELK3 3′ UTR or mutant ELK3 3′ UTR was established. Next, the cells were co-transfected with dual luciferase vector and miR-507 or miR-NC mimics. The relative luciferase activities were detected after 48 h.
Isolation of total RNA and quantitative polymerase chain reaction (qPCR)
Trizol reagent was used to extract total RNA from clinical specimens and cell lines, according to the manufacturer’s instructions (Takara, Japan). To remove DNA contamination, RNase-free DNase I was used. Primers for LINC00525, ELK3, CD44, SOX2, OCT4, miR-507, and internal control including GADPH and U6 were used for amplification of these genes. The sequences are presented in Supplementary Table S1. To analyze the accuracy of the PCR reaction, the Melt curves were used. To evaluate the expressions of genes, 2−ΔΔCt values were calculated. The mRNA expression values of LINC00525, CD44, SOX2, and OCT4 were normalized to that of GADPH. The mRNA expression value of miR-507 was normalized to that of U6.
Western blotting
Extraction and qualification of proteins were performed according to the previous methods (20). After the membrane was probed with antibodies against LINC00525, CD44, SOX2, OCT4, ELK3, p53, Caspase-3, Bcl-2, and actin overnight at 4°C, the membrane was then incubated with appropriated secondary antibodies. ChemiDoc MP imaging system (Bio-Rad, Hercules, CA, USA) was used to examine chemiluminescence.
Results
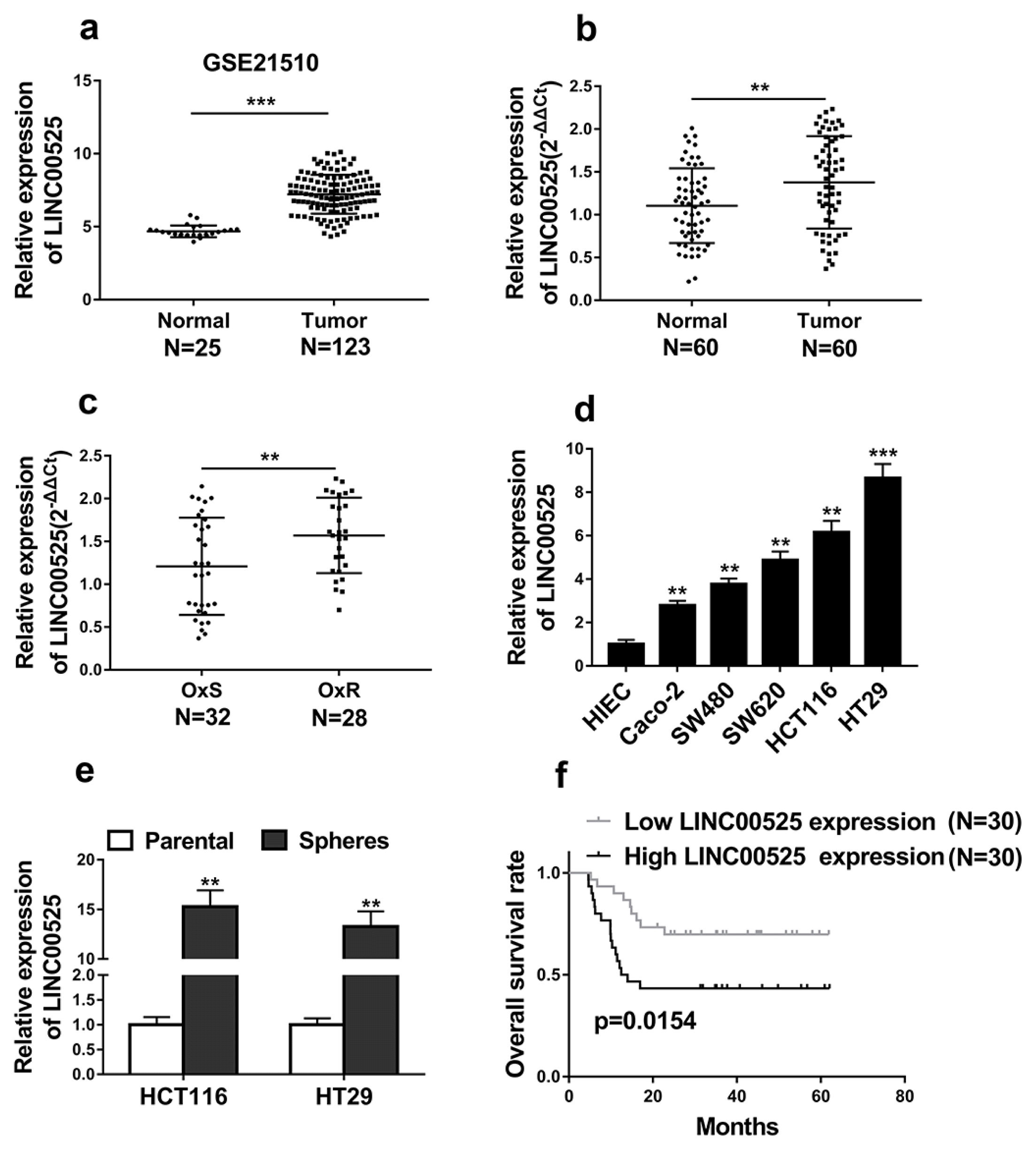
LINC00525 was associated with poor prognosis of CRC
To our knowledge, the roles of LINC00525 in the occurrence and development of CRC are still unknown. Thus, we first explored the relationships between LINC00525 expressions and prognosis of CRC. The relative expressions of LINC00525 were determined in colorectal tumor specimen (n=123) and matched healthy tissues (n=25) from GSE21510 dataset. Expressions of LINC00525 in CRC were significantly increased when compared with that in the healthy tissues (Fig. 1a, p<0.001). In addition, the relative expressions of LINC00525 in colorectal tumor specimen (n=60) and normal intestinal tissues (n=60) were investigated. The expressions of LINC00525 in CRC were significantly increased when compared with that in the normal intestinal tissues (Fig. 1b, p<0.01). We investigated LINC00525 expressions in CRC tissues of oxaliplatin sensitive (OxS) patients (n=32) and oxaliplatin resistant (OxR) patients (n=28). Interestingly, the expressions of LINC00525 in the CRC tissues from OxR patients were higher than those in the CRC tissues from OxS patients (Fig. 1c).
The results further demonstrated that LINC00525 expression levels were higher in CRC cell lines than those in the HIEC (Fig. 1d). We further explored the LINC00525 expression pattern in sphere-forming HTC116 and HT29 cells and their parent cells. Interestingly, LINC00525 expression levels were significantly increased in sphere-forming HTC116 and HT29 cells when compared with those in the parent cells (Fig. 1e, p<0.01).
Furthermore, Kaplan-Meier’s analysis demonstrated that high expressions of LINC00525 were associated with a low survival rate, whereas the low expressions of LINC were associated with high survival rate (Fig. 1f, p=0.0154). Taken together, these results supported that LINC00525 served as a marker for poor prognosis of CRC and could be involved in the regulation of CRC stemness and oxaliplatin resistance.
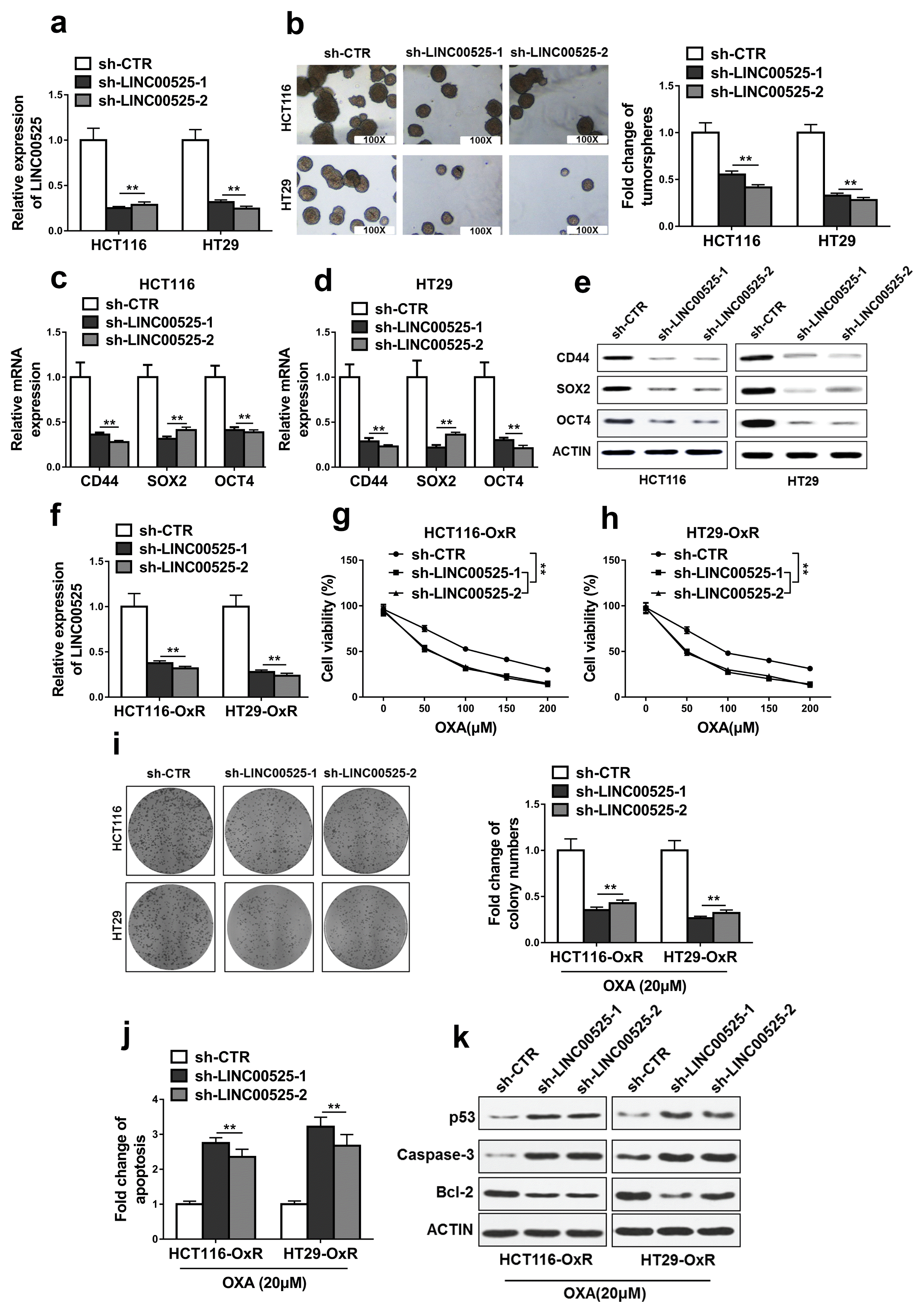
Silencing of LINC00525 inhibited the stemness and oxaliplatin resistance of CRC
First, as shown in Supplementary Fig. S1, we successfully constructed LINC00525 overexpressed HCT116 and HT29 cells (Supplementary Fig. S1a and S1b). Interestingly, the results demonstrated that the mRNA and protein expressions of cancer stem cell markers including CD44, SOX2 and OCT4 were significantly increased in LINC00525 overexpressed HCT116 and HT29 cells when compared with those in the control group (Supplementary Fig. S1c~e). These results supported that that overexpression of LINC00525 was associated with the stemness of colorectal cancer.
We then knocked down LINC00525 in HCT116 and HT29 cells. The results showed that mRNA expressions of LINC00525 in LINC00525 shRNA-transfected cells were significantly decreased when compared with those in the control shRNA-transfected cells, indicating that LINC00525 was successfully knocked down in these cells (Fig. 2a, p<0.01).
The results also demonstrated that LINC00525 knockdown significantly decreased the capacity to form tumorsphere (Fig. 2b, p<0.01) when compared with those in the control group. In addition, the results showed that expressions of cancer stem cell markers including CD44, SOX2, and OCT4 were significantly decreased at both the transcriptional (Fig. 2c and 2d, p<0.01) and post-transcriptional levels (Fig. 2e, p<0.01) in LINC00525 knockdown cells when compared with those in the control group. However, downregulation of protein expressions of SOX2, and OCT4 might be due to the decrease of SOX2, and OCT4 mRNA. Therefore, the regulatory effects of LINC00525 on the protein expressions of SOX2 and OCT4 are directly or indirectly should be clarified in further study.
The results showed that mRNA expressions of LINC00525 in LINC00525 shRNA-transfected cells including HCT116-OxR and HT29-OxR cells were significantly decreased when compared with those in the control shRNA-transfected cells, indicating that LINC00525 was successfully knocked down in these cells (Fig. 2f, p<0.01).
Next, the results demonstrated that LINC00525 knockdown significantly decreased cell viabilities and colony formation abilities of OxR-HCT116 and OxR-HT29 cells when compared with those of the OxR-HCT116 and OxR-HT29 cells transfected with control shRNA (Fig. 2g~ i, p<0.01). LINC00525 knockdown also significantly increased apoptotic cell populations in OxR-HCT116 and OxR-HT29 cells (Fig. 2j, p<0.01). In addition, LINC00525 knockdown increased the expressions of p53 and Caspase-3, decreased the expressions of Bcl-2 when compared with those of the OxR-HCT116 and OxR-HT29 cells transfected with control shRNA (Fig. 2k).
All results suggested that LINC00525 knockdown inhibited the stemness and oxaliplatin resistance of CRC.
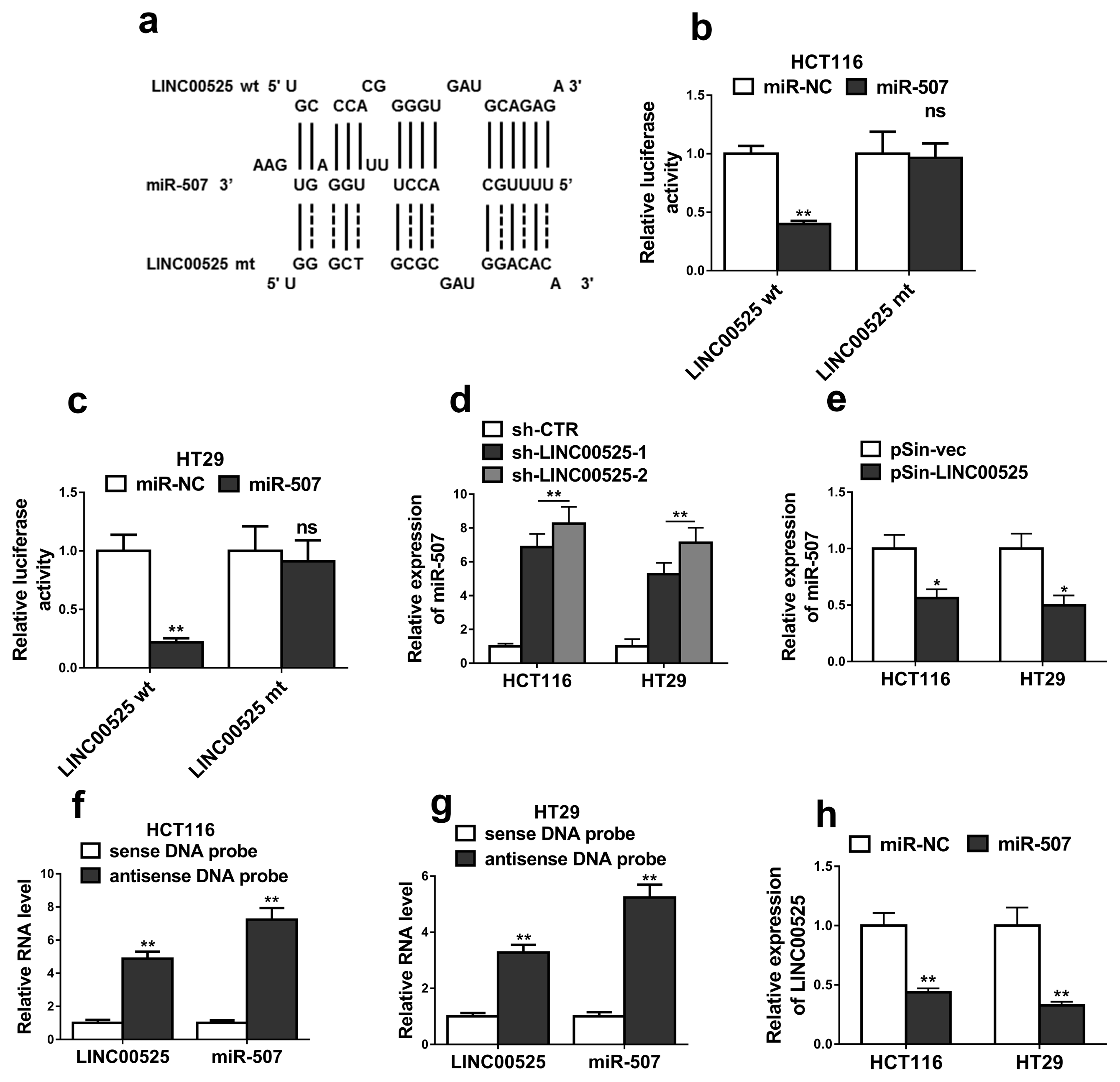
MiR-507 was a direct target of LINC00525
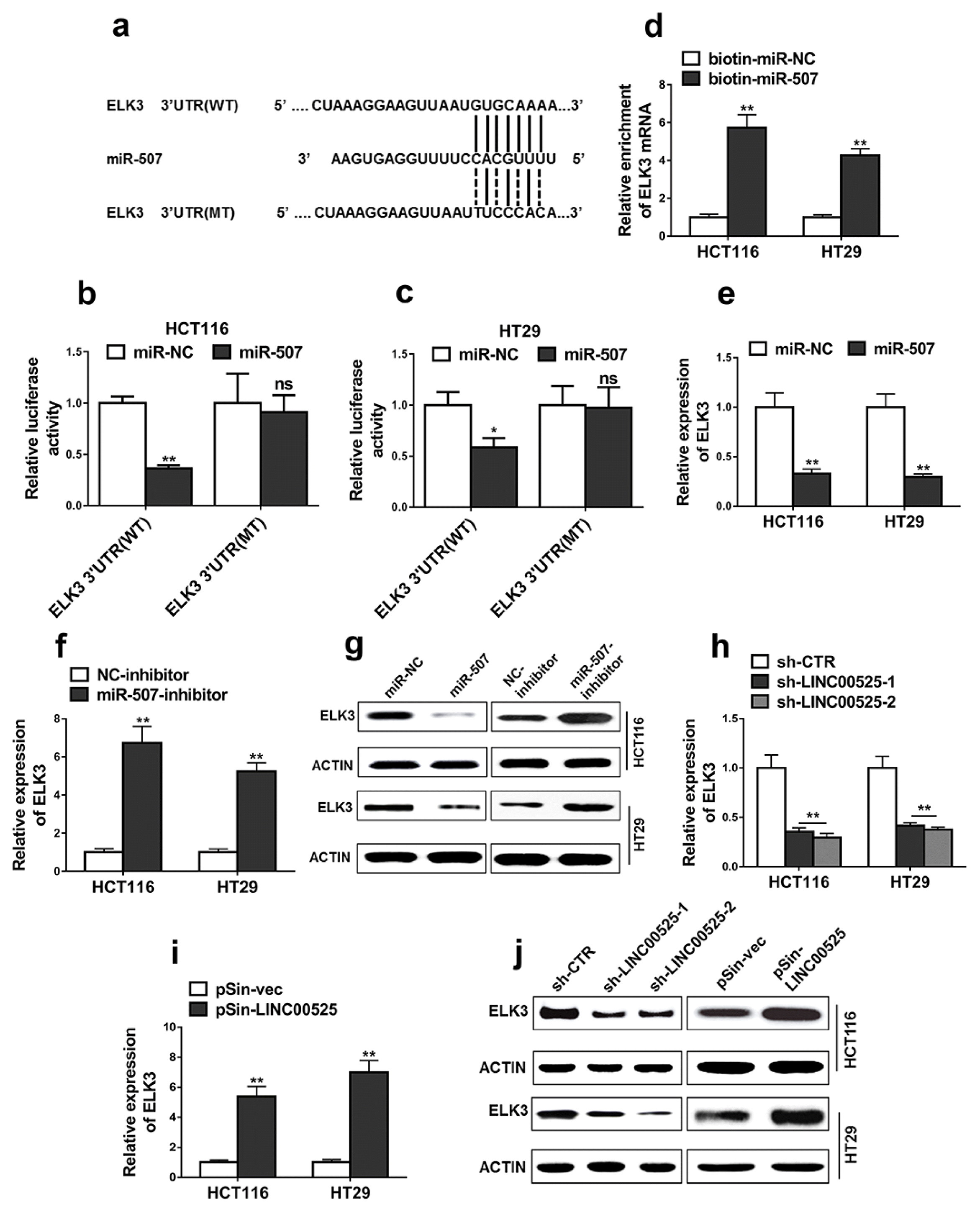
We screened the target miRNAs of LINC00525 using miRcode and identified miR-507 as a target of LINC00525. As shown in Fig. 3a, the binding sites between LINC00525 transcript and miR-507 were predicted using RNAhybrid. Dual luciferase reporter assay showed that miR-507 mimics dramatically decreased the luciferase activities of LINC00525 wt in the HCT116 (Fig. 3b, p< 0.01) and HT29 cells (Fig. 3c, p<0.01). However, luciferase activities between miR-507 mimics and miR-NC mimics in the cells transfected with LINC00525 mt showed no significant difference. These results supported that LINC00525 interacted with miR-507.
Next, we found that LINC00525 knockdown significantly increased the expressions of miR-507 in the HCT116 and HT29 cells (Fig. 3d, p<0.01). Interestingly, when LINC00525 was overexpressed in the cells, the expressions of miR-507 were significantly decreased (Fig. 3e, p<0.01). RNA-pulldown assay was performed to investigate whether LINC00525 directly interacted with miR-507. The results demonstrated that the relative expressions of LINC00525 and miR-507 were dramatically increased in anti-sense DNA probes group, indicating that LINC00525 directly interacted with miR-507 in the HCT116 and HT29 cells (Fig. 3f and 3g, p<0.01). Additionally, the relative expressions of LINC00525 in the HCT116 and HT29 cells were significantly decreased when the cells treated with miR-507 mimics (Fig. 3h).
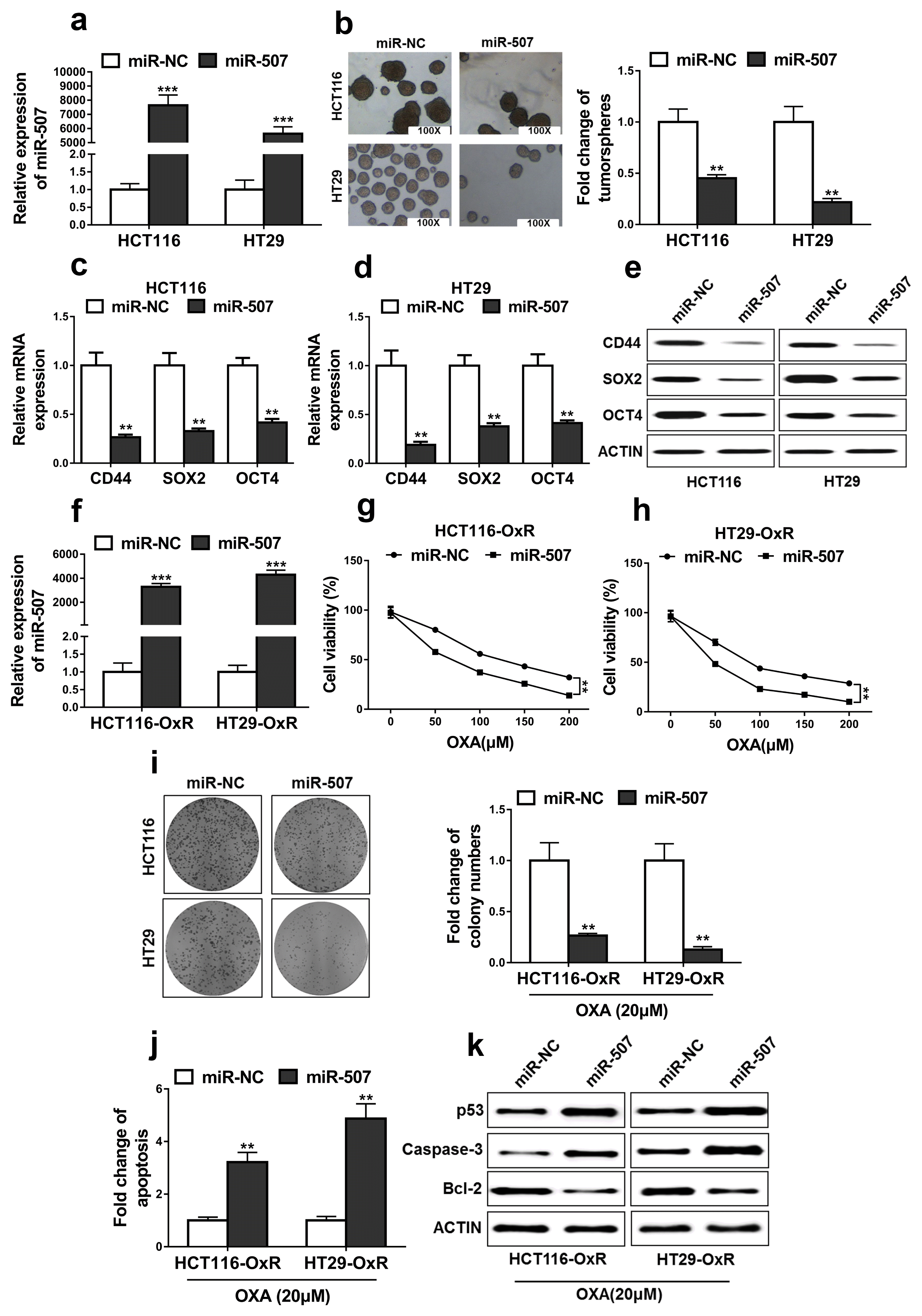
MiR-507 inhibited the stemness and oxaliplatin resistance of CRC
CRC cells were transfected with miR-507 mimics or mimics negative control (miR-NC). Relative expressions of miR-507 were significantly increased in the cells transfected with miR-507 mimics when compared with that in cells transfected with miR-NC (Fig. 4a, p<0.001). Next, we found that overexpression of miR-507 significantly decreased the capacity to form tumorspheres in HCT116 and HT29 cells (Fig. 4b, p<0.01). Additionally, expressions of cancer stem cell markers were evaluated in the cells transfected with miR-507 mimics or miR-NC. The results demonstrated that overexpression of miR-507 significantly decreased the expressions of cancer stem cell markers including CD44, SOX2, and OCT4 at both the transcriptional (Fig. 4c and 4d, p<0.01) and post-transcriptional levels (Fig. 4e).
HCT116-OxR and HT29-OxR cells with miR-507 overexpression were successfully constructed (Fig. 4f). The results demonstrated that miR-507 overexpression significantly suppressed cell viabilities and colony formations when compared with that of miR-NC transfected cells (Fig. 4g~i, p<0.01). Additionally, miR-507 overexpression significantly increased cell apoptosis rates when compared with that of miR-NC transfected cells (Fig. 4j, p<0.01). miR-507 overexpression also increased the expressions of p53 and Caspase-3, decreased the expressions of Bcl-2 when compared with those of the OxR-HCT116 and OxR-HT29 cells transfected with miR-NC (Fig. 4k). Taken together, these data demonstrated that miR-507 inhibited the CRC stemness and oxaliplatin resistance in HCT116 and HT29 cells.
ELK3 was a direct target of miR-507
Targetscan was used to predict the possible targets of miR-507 and ELK3 was identified. The binding sites between miR-507 and ELK3 were shown in Fig. 5a. Additionally, the dual luciferase reporter assay demonstrated that miR-507 mimics specifically decreased the luciferase activities of wide-type ELK 3′-UTR in the HCT116 (Fig. 5b, p<0.01) and HT29 cells (Fig. 5c, p<0.05), indicating that effects of miR-507 on ELK3 expression.
The results demonstrated that ELK3 mRNA levels were significantly increased in bio-miR-507 pulldown group (Fig. 5d, p<0.05), indicating that miR-507 interacted with ELK3. In addition, overexpression of miR-507 significantly decreased the relative expressions of ELK (Fig. 5e, p<0.05). Interestingly, the results demonstrated that inhibition of miR-507 significantly decreased the relative expressions of ELK in the HCT116 (Fig. 5f, p<0.01) and HT29 cells (Fig. 5g, p<0.01).
We then investigated the effects of LINC00525 on the expression pattern of ELK3. The results demonstrated that LINC00525 knockdown significantly decreased the mRNA and protein expressions of ELK3 (Fig. 5h and 5j, p<0.01), whereas LINC00525 overexpression suppressed the mRNA expressions of ELK3 (Fig. 5i, p<0.01). The effects of LINC00525 on the regulation of ELK are at the transcriptional levels (Fig. 5h~i). However, since the decrease of mRNA of ELK leaded to the decrease of protein expression of ELK. Therefore, the regulatory effects of LINC00525 on the protein expressions of ELK should be clarified in further study.
Restoration of ELK3 expression abrogated the effects of LINC00525 knockdown
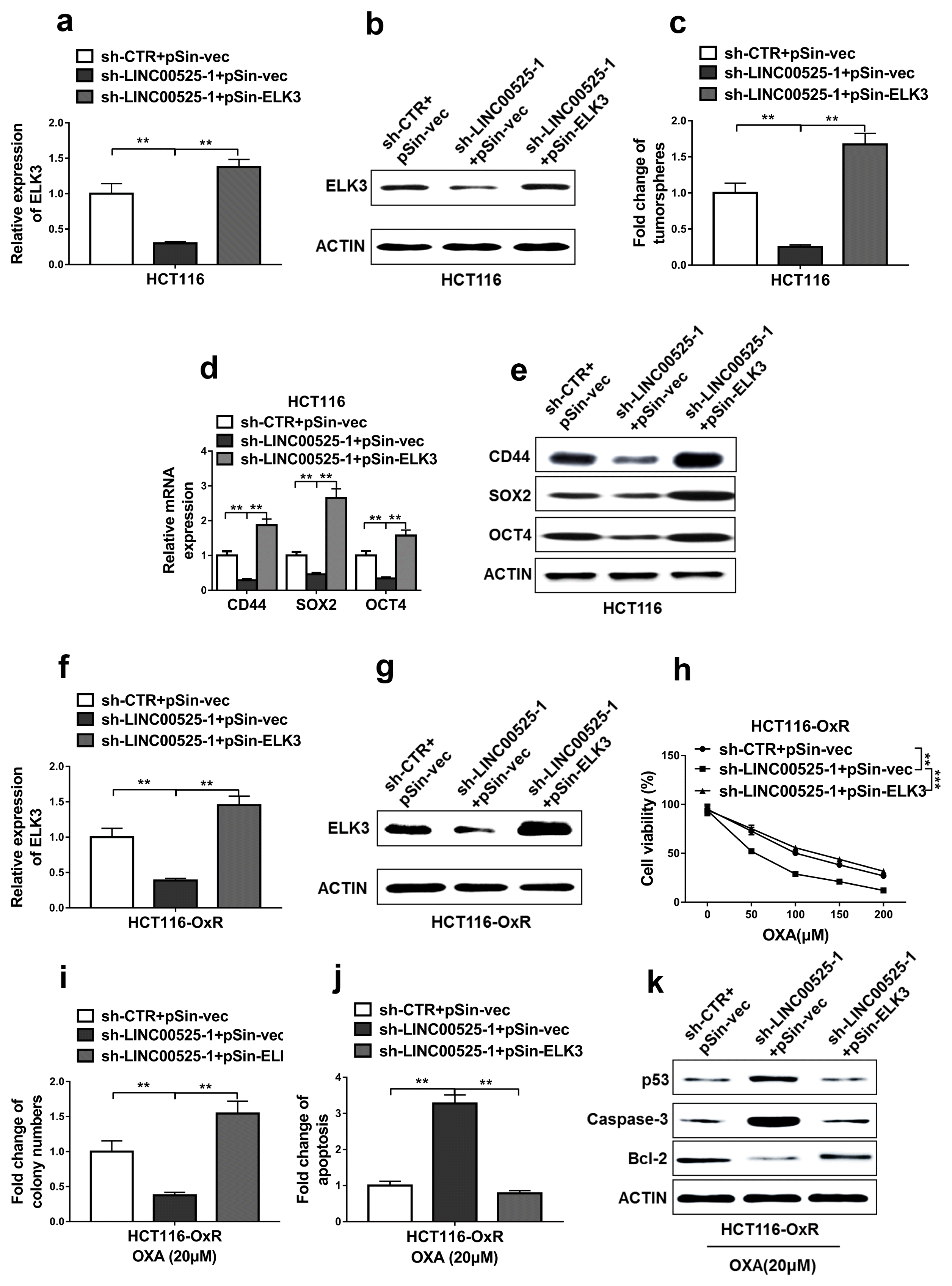
First, we successfully restored the ELK3 expression in the HCT116 cells transfected with LINC00525 shRNA (Fig. 6a and 6b). The results demonstrated that restoration of ELK3 expression significantly increased the capacity to form tumorsphere of LINC00525 knockdown cells (Fig. 6c). Additionally, restoration of ELK3 expression also significantly increased the expressions of cancer stem cell markers including CD44, SOX2, and OCT4 in the LINC00525 knockdown cells (Fig. 6d and 6e).
HCT116-OxR cells were successfully transfected with pSin-ELK3, as shown in Fig. 6f and 6g. Next, cell viabilities and apoptosis were evaluated. The results demonstrated that restoration of ELK3 expression increased the cell viabilities and colony formations in LINC00525 knockdown cells when compared with that of LINC00525 knockdown cells transfected with pSin-vec (Fig. 6h and 6i). Interestingly, restoration of ELK3 expression in HCT116-OxR cells transfected with LINC00525 shRNA significantly decreased the apoptosis rates when compared with HCT116-OxR cells cotransfected with LINC00525 shRNA and pSin-vec (Fig. 6j). Apart from that, restoration of ELK3 expression in HCT116-OxR cells transfected with LINC00525 shRNA also significantly decreased the expressions of p53 and Caspase-3, increased the expressions of Bcl-2 when compared with HCT116-OxR cells cotransfected with LINC00525 shRNA and pSin-vec (Fig. 6k). These results supported that restoration of ELK3 expression abrogated the effects of LINC00525 knockdown in oxaliplatin resistant CRC cells.
Discussion
As the third-generation of platinum compound, oxaliplatin exhibits promising properties in the treatment of CRC (21). In clinical trials, oxaliplatin produces a response rate of 12% to 24% for patients with previously untreated CRC (22). When it combined with 5-flourouracil, more than 50% response rate of metastasis CRC patients is reported (23). Recently, strategies have also been developed to combine oxaliplatin with monoclonal antibodies including panitumumab, cetuximab, and bevacizumab in the treatment of metastasis CRC patients (24). Unfortunately, metastatic CRC patients finally developed to oxaliplatin resistance and succumbed to the disease (4). Therefore, identification of some regulators able to overcome oxaliplatin resistance can be used to improve the response rate of CRC patients.
CSCs are naturally resistant to chemotherapy due to their ability for DNA repair and multidrug resistance ABC transporters expression (7). Those CSCs confer drug resistance to their progeny, resulting in the tumors eventually become less sensitive to the following chemotherapy (7, 8). Therefore, strategies for regulation of CSC stemness can be applied to overcome oxaliplatin resistance. In the current study, we identified a novel lncRNA, LINC00525, as a regulator for CRC stemness. First, we found that expressions of LINC00525 in the CRC tissues from OxR patients were much higher than those in the CRC tissues from OxS patients. Interestingly, we observed that LINC00525 knockdown dramatically decreased cancer stem cell markers including CD44, SOX2, and OCT4 at both the transcriptional and post-transcriptional levels. More importantly, LINC00525 knockdown resulted in the decrease of cell viabilities and colony formation abilities, the increase of cell apoptosis rate in the oxaliplatin resistant cells. These results supported that LINC00525 was associated with CRC stemness. LINC00525 knockdown can be used to regulate CRC stemness, thereby increasing sensitivities of CRC to oxaliplatin.
One important function of lncRNAs is to act as endogenous miRNA sponges (25). LncRNA can regulate miRNA activities by lncRNA-miRNA interactions (26, 27). Therefore, in the current study, we explored the possible target miRNAs of LINC00525 using miRcode and identified miR-507 as a target of LINC00525. One study performed by Jia et al. (28) has demonstrated that miR-507 regulates cell migration and invasion in breast cancer cells. In another study, Wang et al. (29) have reported that miR-507 inhibits the levels of vascular endothelial growth factor C in human chondrosarcoma cells. The current study supported that miR-507 regulated the stemness and oxaliplatin resistance of CRC in part by ELK3 suppression. Our results, along with the previous studies, suggested that miR-507 functions an important marker for the development of different types of cancers. More importantly, we found that a novel lncRNA, LINC00525, regulated the expressions of miR-507, thereby regulating CRC stemness and oxaliplatin resistance.
ELK3 is a transcription factor belonging to Ets family (30). It can be activated by different signaling pathways including PI3K/Akt/mammalian target of rapamycin (mTOR) and ERK signaling pathways (31). Some studies have identified that ELK3 is involved in the incidence and progression of different types of cancers. For instance, it has reported that ELK3 regulates cell migration and invasion in breast cancer and hepatoma cells (32) The current study demonstrated that ELK3 was involved in the regulation of CRC stemness and resistance to oxaliplatin. We found that ELK3 was a direct target of miR-507 and restoration of its expression abrogated the effects of LINC00525 knockdown. Interestingly, another study performed by Park et al. (33) has reported that ELK3 suppression regulates sensitivities of breast cancer cells to doxorubicin. These results suggest that ELK plays an important role in the regulation of cancer cell chemotherapy resistance. More importantly, the results showed that miR-507 regulated expression of ELK3. Inhibition of miR-507 significantly increased the expressions of ELK3. Therefore, apart from PI3K/Akt/mTOR and ERK-mediated signaling pathways, we identified that the inhibition of miR-507 is another pathway triggering ELK3 activation.
This study demonstrated that LINC00525 functioned as an oncogene and its high expression was associated with prognosis of CRC patients. LINC00525 promoted the stemness and chemoresistance of CRC by targeting miR-507/ELK3 axis. Silencing LINC00525 inhibited the stemness and oxaliplatin resistance of CRC. These results supported that LINC00525 might serve as a prognostic biomarker and a potential target for CRC therapy.
 XML Download
XML Download