

 PDF
PDF Citation
Citation Print
Print


Introduction
The direct differentiation of somatic cells into tissue-specific cells such as induced neurons (1) and neural precursor cells (2–5) may serve as an attractive approach in the field of stem cell research. The first study in this direction was reported by Vierbuchen et al. (1) using neuronal lineage-inducing transcription factors such as Brn2, Ascl1, and Myt1L (BAM). These factors converted mouse fibroblasts into neuronal cells without transitioning into a pluripotent state. Since then, several strategies have been employed to perform direct conversion. Direct reprogramming using non-integrating transcription factors (6, 7), chemical (small molecule) treatment (8, 9), and stromal cell-derived inducing activity (10) has been carried out. The low efficiency of this method is, however, a major limitation. To enhance the conversion efficiency, some studies have suggested that cell topography interactions control cell phenotypes (11, 12) and ameliorate tissue-specific cell generation (13, 14). Nanotopography may substantially encourage cellular behavior from morphological changes to differentiation. Furthermore, the extracellular environment may provide signaling cues to adherent cells (15). Matrix elasticity may direct human mesenchymal stem cells to differentiate into specific lineages. A soft matrix may induce neurogenic phenotype, while stiffer matrices may induce myogenic and osteogenic phenotypes (16). Engineered nanometer-scale scaffolds hold great promises for stem cell differentiation and transplantation (17). Recent reports have shown that mechanical factors such as stress (18–20), adhesion area (21, 22), substrate elasticity (16), and topography on micro- and nano-scale scaffolds (23–25) may induce differentiation of stem cells. The development of lithography-based platforms such as silicon-etched, photocured, and replica-molded patterns has enormously contributed to the biological study evaluating cellular behavior on well-ordered surfaces (26). In this study, we hypothesized that topographical cue combined with our previous conversion research method (4) may influence neural precursor cells (iNPCs)/dopaminergic (iDA) neuron generation. iNPC generation was observed earlier and iNPC-derived DA neurons exhibited high capacity to mature on fiber and dot nanoscale structures.
Go to : 
Materials and Methods
Fabrication of electrospun aligned nanofiber scaffold and nanostructured substrates with UV-assisted capillary force lithography (CFL)
Polycaprolactone (PCL; MW, 80,000; Sigma-Aldrich, MO, USA) was dissolved in a mixture of chloroform (Samchun Pure Chemical, Korea) and dimethylformamide (Sigma-Aldrich, MO, USA) at a volume ratio of 75:25. Electrospinning PCL solution was used at 20% concentration (weight-to-volume ratio), as standardized by our group to obtain a unidirectionally aligned and homogeneous nanofiber scaffold array. The electrospinning process was conducted as follows: PCL solution was infused with a disposable syringe and a metal needle (23-gauge) at a rate of 1.5 ml/h to form polymer drops. A DC voltage of 15 kV was applied to the tip of the needle to induce electrospinning. The aligned nanofiber was harvested with an inclined-gap method, wherein the ejecting electrospun nanofiber was aligned in an electromagnetic field formed by two aluminum metal strips. The repulsive force between nanofibers allowed the formation of a well-oriented nanofiber scaffold. To control the nanofiber density, we repeatedly transferred the nanofiber array to a punched PDMS slab (2-cm diameter). The electrospinning time for each transfer was ~1 min and the transfer was repeated for five times to fabricate a single cell culture scaffold. The schematic diagram of the fabrication procedure is illustrated in Fig. 1.
 | Fig. 1Fabrication process of the hybrid nanofiber scaffold and planar SEM images of the fabricated nanopatterns by UV-assisted capillary force lithography (CFL). Scale bars represent 5 and 1 μm in the panel and inset images: (A) Electrospinning process to secure aligned nanofibers and transfer onto the film, (B) Image of the nanofiber based sieve for cell sheet (transfer number: 6, density: 173 fiber/mm) with PDMS frame (20 mm diameter hole, frame size: 30 mm×30 mm, height: 5 mm) and (C) 400 nm diameter and 400 nm pitch dots (left) and 400 nm width and 400 nm pitch lines (right).
|
A small amount (0.1~0.5 ml) of UV curable PUA prepolymer was drop-dispensed on a silicon master with positive patterns (features sticking out), and a supporting polyethylene terephthalate (PET) film was carefully placed on the top surface to allow conformal contact. The PET film used in this study was surface modified with urethane groups to increase adhesion with an acrylate-containing monomer (Minuta Tech., Korea). The silicon masters were prepared by photolithography or electron-beam lithography. The resin was exposed to UV (wavelength: 250~400 nm) for 17 s at an intensity of 100 mW/cm2, and the cured first replica was peeled-off from the silicon master using a sharp tweezer. The first replica was overnight exposed to UV to remove any uncured active groups on the surface. The second replica was prepared using a capillary molding process on glass coverslips with the overcured first PUA as a mold, resulting in a pattern similar to that of the silicon master. After curing, the first replica was removed from the surface using sharp tweezers. The fabricated PUA nanopatterns were sterilized by rinsing with isopropyl alcohol and distilled water, and coated with 0.1% gelatin for 1 h at room temperature prior to cell culture (27). Two types of nanopatterns that varied in size (from 400 nm) were used, including dots (400 nm in diameter) and lines (400 nm in width). For consistency, the pattern height was fixed to 500 nm, as verified by atomic force microscopy measurements (data not shown).
Fibroblast isolation
Animals were housed and cared according to the institutional animal care and use committee (IACUC 2017-0137A) guidelines of Hanyang University. Rat embryonic fibroblasts (REFs) were isolated from Sprague-Dawley (DaeHan BioLink, Seoul, Korea) rat embryos (embryonic day 14) after removing all organs, including spinal cord. REFs were cultured in Dulbecco’s modified Eagle’s medium (DMEM) with high glucose (Invitrogen, Grand Island, NY, USA) supplemented with 10% fetal bovine serum (FBS; Invitrogen), 1% penicillin-streptomycin (Invitrogen), 0.1 mM nonessential amino acids (Invitrogen), 0.1 mM β-mercaptoethanol (Sigma-Aldrich, St. Louis, MO, USA), and 2 mM l-glutamine (Sigma-Aldrich). REFs were used after at least three passages.
Retroviral production
Retroviral construction and production were performed as previously described (4). In brief, retroviral vectors were transfected into 293GPG packaging cells and supernatants containing viral particles were harvested. Retroviruses expressing Brn2 (B), Ascl1 (A), Myt1l (M), Bcl-xL (X), Nurr1 (N), and Foxa2 (F) were stored at −80°C until use.
iNPC generation and iDA neuron differentiation
To generate iNPCs, REFs were cultured on gelatin-coated dishes. The following day, the cells were transduced with Brn2, Ascl1, Myt1l and Bcl-xL (BAMX) retroviruses. After 16~20 h, the medium was replaced with fresh REF medium containing 100 ng/ml human fibroblast growth factor 8 (FGF8; PeproTech, Rochy Hill, NJ). After 2 days, the cells were placed in neural precursor cell induction/expansion medium (NIM/NEM: N2 medium) supplemented with 20 ng/ml of basic fibroblast growth factor (R&D Systems, Minneapolis, MN, USA), 100 U/ml of recombinant human leukemia inhibitory factor (EMD Millipore, Billerica, MA, USA), 2 μg/ml of doxycycline (Sigma-Aldrich) with or without 100 ng/ml of FGF8 and transferred onto several nano-patterns pre-coated with 15 μg/ml of poly-l-ornithine (Sigma-Aldrich) and 1 μg/ml of fibronectin (Sigma-Aldrich).
Transduced cells showed diverse morphologies in each pattern. In particular, neural precursor-like cells were observed in fiber and dot 1:1 condition. At day 16, iNPCs were transduced with Nurr1 and Foxa2 for the generation of dopaminergic neurons and subjected to differentiation on the following day (differentiation medium including 0.2 mM ascorbic acid [Sigma-Aldrich], 20 ng/ml of brain-derived neurotrophic factor [R&D Systems], 20 ng/ml of glial cell line-derived neurotrophic factor [R&D Systems], and 250 μg/ml of dibutyryl-cAMP [Sigma-Aldrich] in N2 medium).
Immunocytochemistry and antibodies
Cells were fixed with 4% paraformaldehyde in phosphate-buffered saline (PBS) for 20 min at room temperature, permeabilized, and blocked in a blocking solution (10% normal goat serum, Pel-Freez, Rogers, AR, USA), 0.3% Triton X-100 (Sigma-Aldrich in PBS) for 1 h. The samples were stained with primary antibodies for 1 day and then incubated for 1 h with secondary antibodies in the dark. Cells were photographed using an epifluorescence microscope (Leica Micro systems, Wetzlar, Germany). Monoclonal primary antibodies used were as follows: NESTIN (BD Bioscience, Franklin Lakes, NJ), microtubule-associated protein 2 (MAP2; Sigma-Aldrich). Polyclonal primary antibodies used included those against SOX2 (Chemicon, Temecula, CA) and tyrosine hydroxylase (TH; Pel-Freez, Rogers, AR, USA).
Cell counting and statistical analysis
Cell counting was performed on a random selection of 5~10 uniform microscopic fields/wells, with 3~4 wells per experimental condition. All values were confirmed with at least three independent experiments. Data are expressed as mean±SEM. When more than two groups were compared, a paired t-test was performed using SigmaPlot for Windows, version 10.0 (Systat Software GmbH, Erkrath, Germany) (5).
Go to :
Results
Nanoscale structures are suitable for cell survival and direct conversion process
We have previously shown that NPCs may be generated from rodent fibroblasts transduced with the ectopic gene expression method (4). The nanoscale pattern protocol for NPC/DA neuron generation from human pluripotent stem cells and somatic cells has been reported (13, 14, 25). To induce NPCs from fibroblasts, we combined these two protocols, as shown in Fig. 2A. After 3 days of BAMX transduction (DIV 4), cells were transferred on several nanoscale scaffolds, such as fiber, dot, and line. During NPC induction/expansion, cell morphology changed, as evident from the small and slender cells, with no effect on survival and proliferation properties (Fig. 2B).
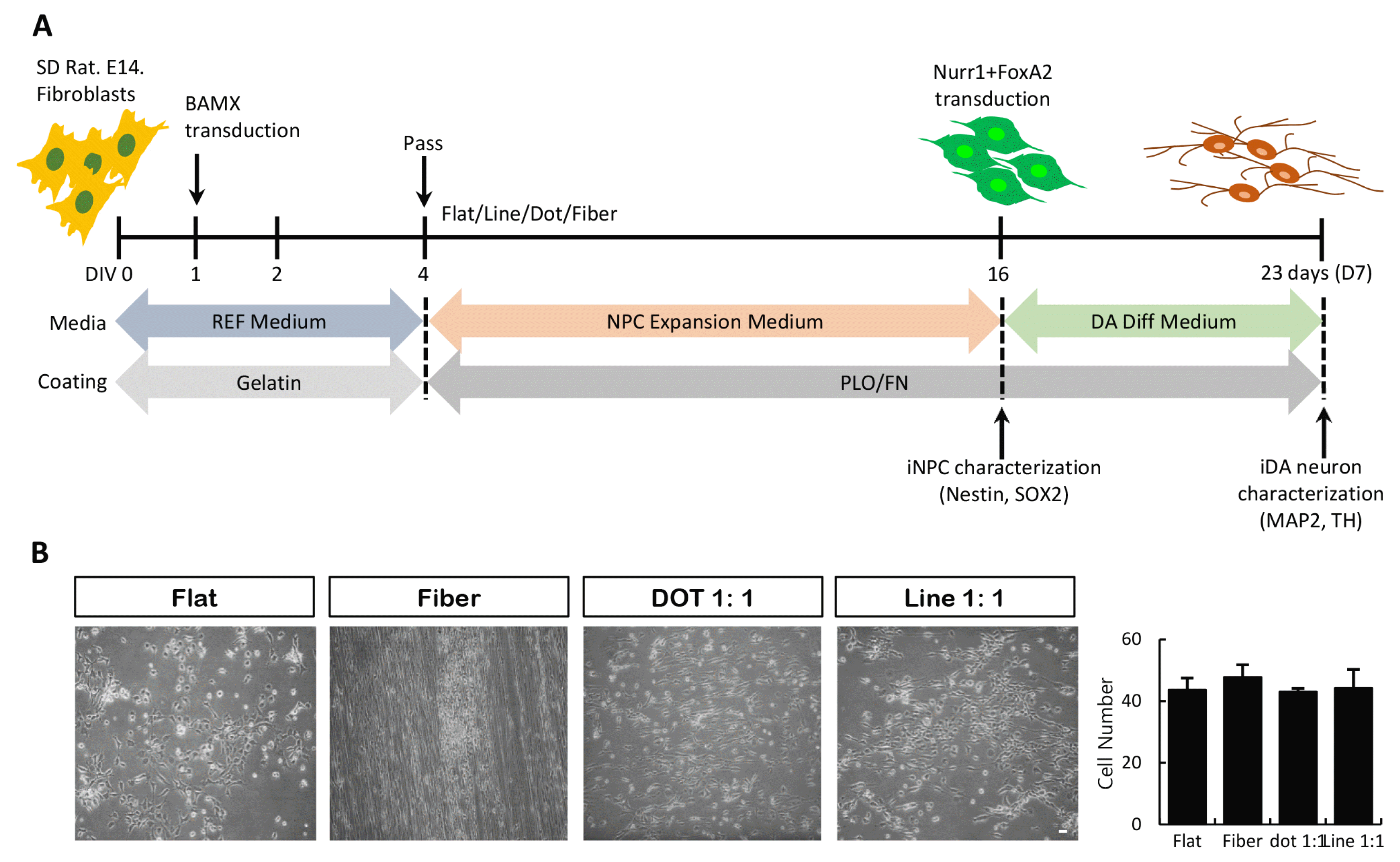
Direct conversion of fibroblasts into iNPCs and iDA neurons on nanoscale pattern substrates
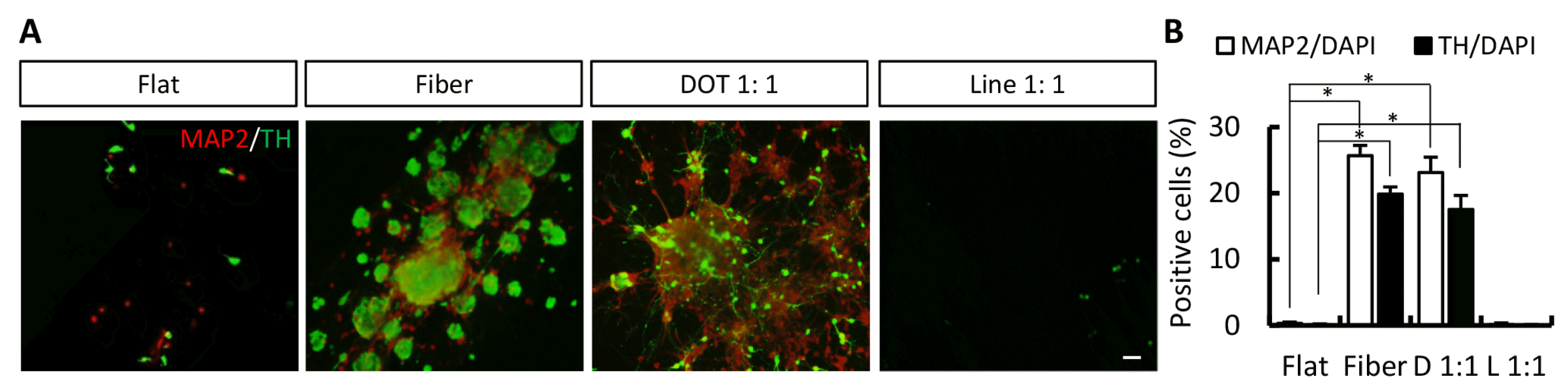
At the end of the NPC induction/expansion stage, the transduced cells exhibited cellular morphology of neural precursors. To confirm iNPCs, cells were immunostained for nestin and SOX2. NESTIN-positive cells were observed on all nanoscale pattern surfaces (Fig. 3A and 3B; NESTIN-positive cells on flat, fiber, dot, and line surfaces were 3.68%±1.68%, 11.95%±0.98%, 9.77%±2.25%, and 3.62%±0.36%, respectively). SOX2-positive cells were also observed on fiber and dot surfaces (Fig. 3A and 3B; SOX2-positive cells on flat, fiber, dot, and line surfaces were 0.97%±0.22%, 8.89%±0.91%, 2.68%±0.97%, and 0.74%±0.23%, respectively). Next, we evaluated the ability of iNPCs to differentiate into iDA neurons on nanoscale pattern surfaces. After differentiation, MAP2-positive and TH-positive cells were expressed on fiber and dot pattern surfaces (Fig. 4A and 4B; MAP2-positive cells on flat, fiber, dot, and line surfaces were 0.32%±0.21%, 25.68%±1.54%, 23.12%±2.32%, and 0.11%±0.26%, respectively; TH-positive cells on flat, fiber, dot, and line surfaces were 0.10%±0.13%, 19.84%±1.11%, 17.53%±2.13%, and 0.02%±0.11%, respectively). Taken together, the nanoscale technique, particularly the fiber and dot scaffold, is useful for the direct conversion of somatic cells.
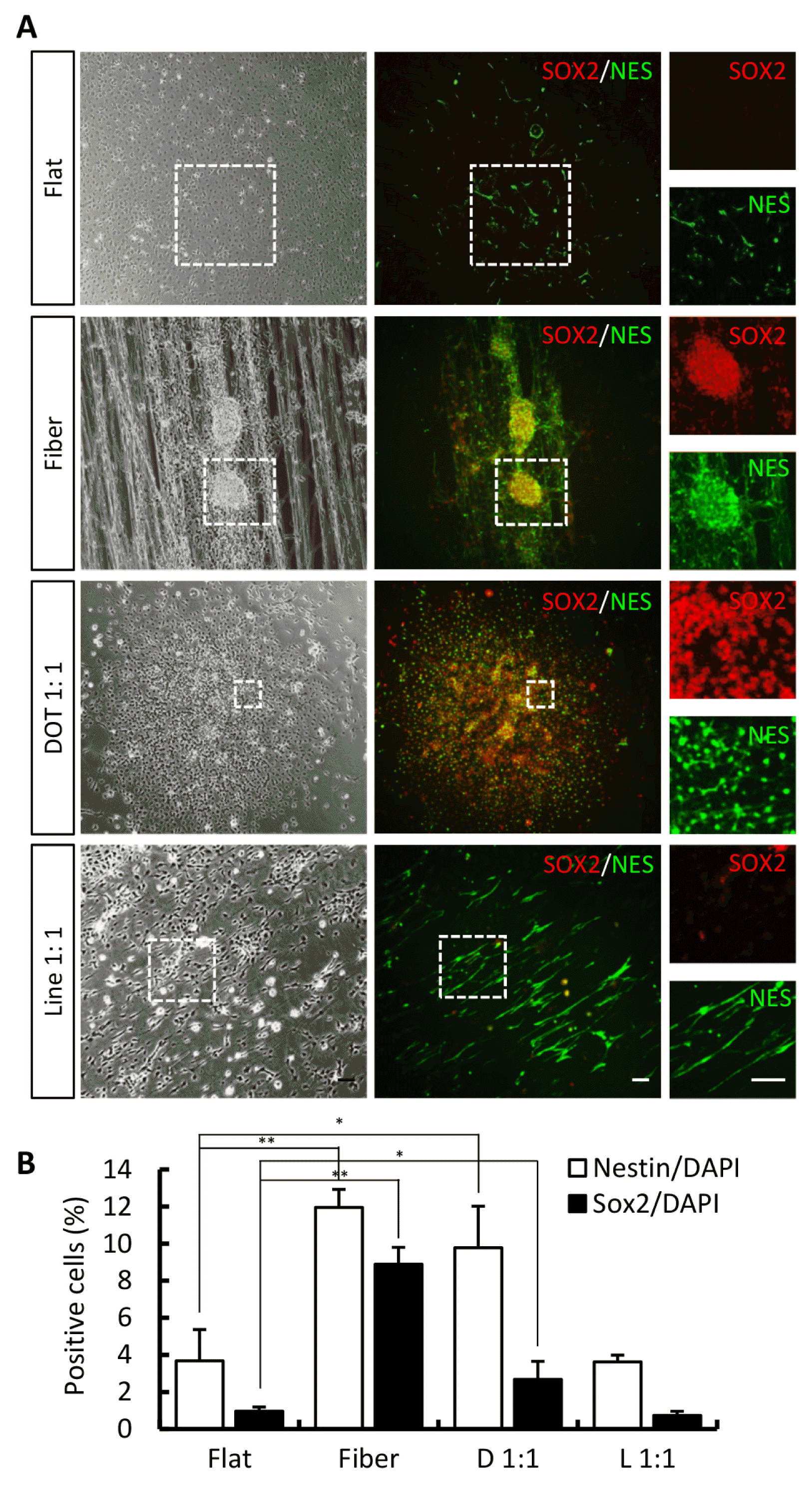 | Fig. 3Characterization of iNPCs among diverse nanoscale pattern surfaces. (A) Phase-contrast images of transduced cells and immunostained image of NPC markers, nestin and SOX2, on each nanoscale pattern. White dot box indicates high magnification images. (B) Calculation graph. Error bars denote the standard error of the mean (SEM, n=3, *p<0.05, **p<0.01). Scale bar=20 μm. D: Dot, L: Line.
|
Go to :
Discussion
Generation of iNPC/iDA neurons from fibroblasts has been the focus of stem cell regeneration research. To obtain iNPC/iDA neurons in vitro and in vivo, various types of cell sources and materials have been developed. We have successfully demonstrated the generation of expandable iNPCs and DA neurons from somatic cells using exogene expression (4) and stromal cell-derived inducing activity (SDIA) methods (10). Various types of experimental procedures have been developed. A recent technique based on nanoscale pattern structure has gathered considerable attention.
The investigation of cell fate modulated using nanoscale pattern environment is one of the active areas in stem cell engineering because of the possible side-effects caused by biochemical signals and biophysical cues (28). In vivo, all adherent cells encounter topographical features from neighboring cells and the extracellular matrix. The functions of cells, in general, and of neurons, in particular, depend on their shape and polarity (13). Cell polarity is controlled by the confinement of focal adhesions on the gratings. Focal adhesions have been shown to play an important role in topography sensing (19). The surfaces can be altered (activated) at will to tip adhesion/cytoskeletal balance and initiate differentiation, hence better informing biological mechanisms of stem cell growth and direct conversion (29, 30). Based on this mechanism, nanotopographical signal exerts synergic effects that may cause biochemical stimulation.
In our work, we combined nanoscale pattern structures (fiber, dot, and line) with the general conversion process. The transduced cells were transferred onto nanoscale pattern substrates, which resulted in the quick transformation of the cells into NPC-like cells without cell death (data not shown). Thus, the nanoscale pattern substrates are nontoxic and may serve as promising tools in conversion material technique. The fiber and dot pattern substrates enhanced the generation of NPCs (NESTIN- and SOX2-positive; Fig. 3) and DA neurons (MAP2- and TH-positive; Fig. 4).
Go to :
 XML Download
XML Download