

 PDF
PDF Citation
Citation Print
Print


Introduction
Hypoxic culture conditions enhance the proliferation or survival of several cell types such as tumor cells (1, 2), endothelial cells (3), fibroblasts (4), neural stem cells (5), and mesenchymal stem cells (6). Moreover, cellular proliferation is maintained at a high level in long-term hypoxic culture (7).
Although a signal pathway dependent on hypoxia-inducible factor (HIF)-1α or HIF-2α has been reported as the major mechanism for high proliferation in hypoxic conditions (8, 9), identification of other involved signal pathways or molecules is required to understand the response and function of cells under hypoxic condition. The secretome from mesenchymal stem cells in hypoxic culture condition shows beneficial effects on the cells themselves or neighboring cells through autocrine or paracrine signaling (10–12).
Previous studies have reported that fibroblast growth factor (FGF)-17 is expressed in the embryonic brain (13). Moreover, FGF17 increased the proliferation of carcinoma cells (14) and leukemic cells (15), and inhibited the differentiation of oligodendrocyte progenitor cells (16). However, the role of FGF-17 in human mesenchymal stem cells cultured in hypoxic conditions has not yet been investigated.
In this study, we aimed to investigate the role of FGF-17 secreted by human Wharton’s Jelly-derived mesenchymal stem cells (hWJ-MSCs) cultured in hypoxic conditions at late passages based on protein profiling of conditioned medium (CM) of hypoxic hWJ-MSCs.
Materials and Methods
Cell cultures
This study was approved by the Institutional Review Board of Samsung Medical Center and informed consent was obtained from pregnant mothers (IRB. No.2016-07-102). hWJ-MSCs were isolated according to the procedure specified in a previous report (17) and cultured in Alpha Minimum Essential Medium (α-MEM, Gibco, Carlsbad, CA, USA) supplemented with 10% FBS (Gibco) and 0.5% antibiotic/antimycotic (Gibco) at 37°C in normoxic (21% O2) or hypoxic (1% O2) culture conditions with 5% CO2. Media changes and passaging for both conditions were performed in a normoxic environment. When the culture reached 90% confluency, the cells were washed twice with phosphate buffered saline (PBS) and dissociated with 0.25% trypsin-EDTA (Gibco) for 2 minutes at 37°C. Dissociated cells were treated with medium to block trypsin-EDTA activity. After centrifugation, cell pellets were resuspended with medium and seeded at 3,000 cells/cm2. Cell images were acquired with a model CKX41 inverted microscope (Olympus Corporation, Tokyo, Japan). For the supplementary experiments on the role of FGF-17 in hWJ-MSCs, 2 of hWJ-MSCs were purchased from PromoCell (Heidelberg, Germany) and named as hWJ-MSCs-1 and hWJ-MSCs-2.
Protein antibody array of CM from hWJ-MSCs
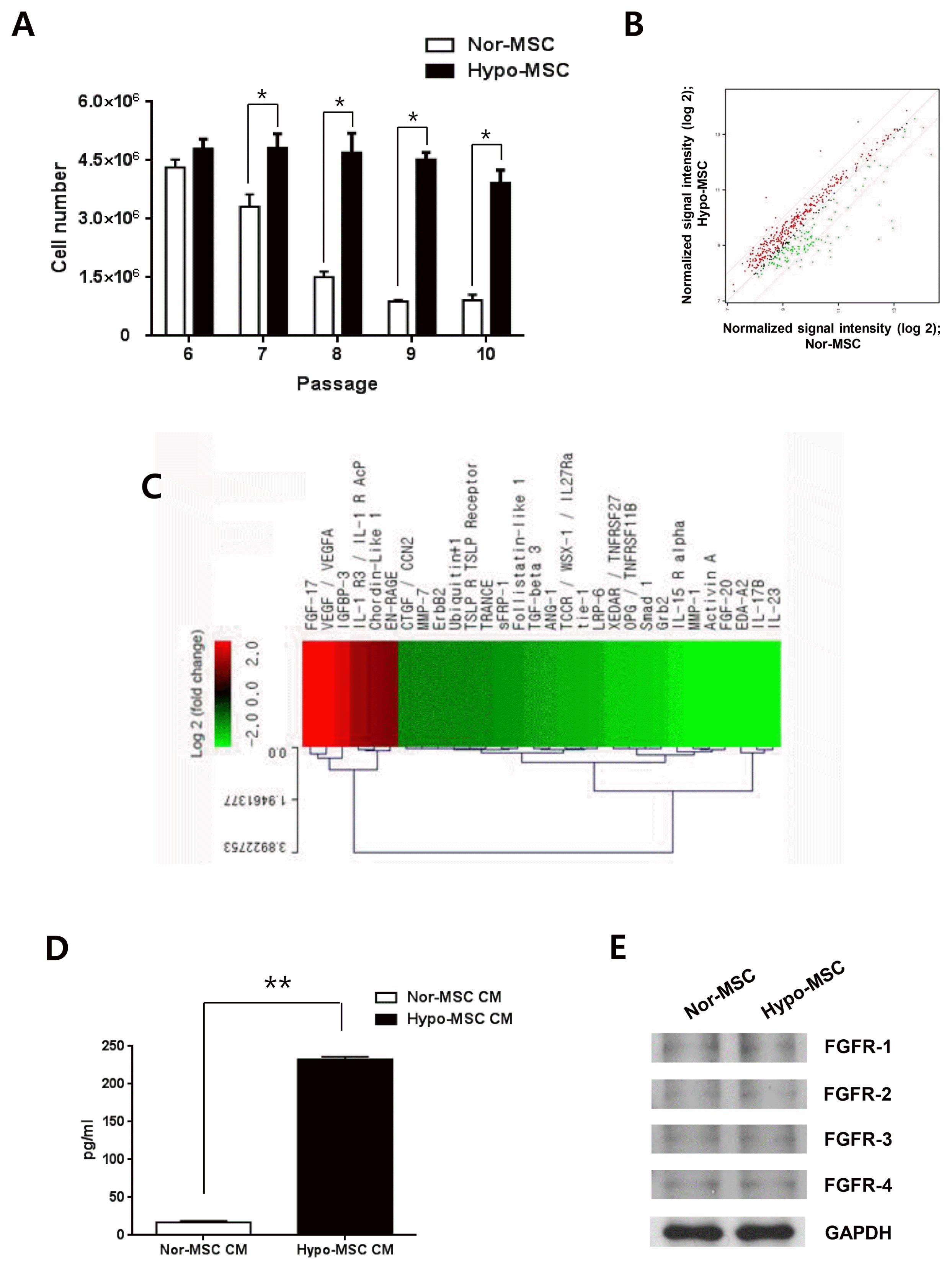
Normoxic or hypoxic hWJ-MSCs at passage 10 were seeded in 100-mm diameter culture plates containing complete medium at a density of 3,000 cells/cm2. When the cells reached 90% confluency the medium was replaced with 12 ml of serum-free medium. After incubation for 18 h, CM was concentrated by centrifugation at 5,000×g for 1 h using an Amicon Ultra-15 centrifugal filter (Millipore, Billerica, MA, USA) with a size cut-off of 3,000 Dalton. Expression of proteins in CM from hWJ-MSCs was measured with a RayBio® Label-based (L-Series) Human Antibody Array (Raybiotech, Inc., Norcross, GA, USA) from E-biogen (Kyung Hee Business Center, Kyung Hee University, Seoul, Korea). The result was acquired from a single sample analysis. Upregulated and downregulated proteins are listed in Fig. 1B, 1C and Table 1.
ELISA
Expression of FGF-17 in CM from normoxic hWJ-MSCs or hypoxic hWJ-MSCs at passage 10 was measured using Human FGF-17 ELISA Kit, Cusabio Technology, LLC, College Park, MD, USA) by manufacture’s manual. Briefly, 100 ul of 1:3 diluted CM from normoxic or hypoxic hWJ-MSCs at passage 10 were added to each well of 96 well plate coated with anti-FGF-17 antibody, and incubated for 2 h at 37°C. In the next step, biotin-antibodies were added to each well for 1 h at 37°C. After washing wells with washing buffer, HRP-avidins were added to each well for 1 h at 37°C. TMB substrates were added to each well for 15 min and stop solutions were added. The optical density of each well was determined using a microplate reader xMark microplate spectrophotometer (BioRad, Hercules, CA, USA) set to 450 nm.
Treatment with siFGF-17 and rFGF-17
For transfection with siRNA, 20 pmol of siFGF-17 (sense, CGACUGAAGGCCUUGCAGA; antisense, UCUGCAAGGCCUUCAGUCG; Bioneer, Daejeon, Korea) was mixed with 1 ul of Lipofectamin 2000 (Invitrogen, Carlsbad, CA, USA) for 20 minutes at room temperature, and then added to normoxic hWJ-MSCs at passage 7 and hypoxic hWJ-MSCs at passage 10. After 6 h, the medium was replaced with fresh complete medium, and the cells were cultured for 48h. Negative control siRNA was used for the control group (SN-1013, Bioneer, Daejeon, Korea).
For rFGF-17 treatment, normoxic hWJ-MSCs at passage 7 and hypoxic hWJ-MSCs at passage 10 were treated with 250, 500, 1,000 ng/ml of rFGF-17 (PeproTech Inc., Rocky Hill, NJ, USA) for 48 h.
For both treatments, cell counting, BrdU proliferation assay, quantitative real-time polymerase chain reaction (QRT-PCR), western blot, and flow cytometry analysis were performed after incubation for 48 h, as described below.
Cell proliferation assay
Cells from passages 6 to 10 were seeded at 3,000 cells/cm2 in normoxic or hypoxic culture conditions. Cells were removed by trypsinization at 90% confluency for each passage and cell numbers were counted using Trypan blue 0.5% solution (Biowest, Riverside, MO, USA) and a hemocytometer. A BrdU cell proliferation assay kit (Millipore) was used to analyze cell proliferation according to the manufacturer’s manuals. Briefly, normoxic or hypoxic hWJ-MSCs were seeded in 96-well plates at 3,000 cells/cm2, incubated overnight, and treated with 1×BrdU solution for 24 h. The medium was removed and the cells were treated with Fixing/Denaturing Solution (Millipore) for 30 min. Cells were incubated with 1×detection antibody and 1×HRP-conjugated secondary antibody (Millipore) for 1.5 h and washed with 1×Wash Buffer (Millipore) After incubation with TMB Substrate (Millipore) for 30 min, STOP solution (Millipore) was added and the absorbance of BrdU was measured at 450 nm using xMark Microplate Spectrophotometer (Bio-Rad, Hercules, CA, USA).
Cell viability assay
3,000 cells/cm2 of normoxic hWJ-MSCs at passage 7 or hypoxic hWJ-MSCs at passage 10 were seeded into the well of 96-well white plates. After 48 h, culture medium was removed and serum free medium was added to the wells. After 1 day incubation, 500 ng/ml of rFGF-17 was treated to the cells for 48 h. For the inhibition of secretory FGF-17 from hypoxic hWJ-MSCs at passage 10, 6,000 cells/cm2 of hypoxic hWJ-MSCs were incubated in 96-well white plates for 48 h. Then, 30 ug/ml of blocking antibody of FGF-17 (R&D Systems, Minneapolis, MN, USA) in serum free medium was treated to the cells for 48 h. In the next step, 100 ul of CellTiter-Glo assay 2.0 reagents (Promega, Madison, WI, USA) was treated to the cells. After 10 min incubation, luminescence from cells was measured with GLOMAX Multi Detection System (Promega, Sunnyvale, CA, USA).
Quantitative real-time polymerase chain reaction (QRT-PCR)
Total RNA was extracted from cells using a RNeasy Plus Mini Kit (Qiagen GmbH, Hilden, Germany) and 200 ng of total RNA was used for reverse transcription with a PrimeScript RT reagent kit (Takara, Bio, Shiga, Japan) according to the manufacturer’s manuals. Real-time PCR was conducted using SYBR Green Realtime PCR Master Mix (Toyobo Co., LTD, Osaka, Japan) with 100 ng template DNA in a 7900 HT Fast Real-time PCR system (Applied Biosystems, Foster City, CA, USA) using reaction conditions of denaturation at 95°C for 15 s and extension at 60°C for 34 s for 40 cycles. The primer sequences are listed in Table 2.
Western blotting
For the analysis of FGF-17 receptors on normoxic hWJ-MSCs and hypoxic hWJ-MSCs at passage 10, cell lysates were harvested from both kinds of cells. For the analysis of intracellular signaling related with FGF-17, cell lysates were harvested from normoxic hWJ-MSCs treated with rFGF-17 and transfected with siRNA against FGF-17 at passage 7 or hypoxic hWJ-MSCs treated with rFGF-17 and transfected with siRNA against FGF-17 at passage 10 using lysis buffer (20 mM HEPES pH 7.6, 20% Glycerol, 250 mM NaCl, 1.5 mM MgCl2, 0.1% Triton X-100, 2 mM PMSF, 1mM DTT, 1 mM NaF and 1 mM Na3VO4) with protease inhibitor cocktail (Roche Diagnostics GmbH, Mannheim, Germany). Quantification of proteins in lysates was performed with Quick Start Bradford 1×Dye Reagent (Bio-Rad), and absorbance was measured at 450 nm using xMark Microplate Spectrophotometer. Protein samples were boiled at 95°C for 15 min and 20 ug of protein from each sample was subjected to SDS–PAGE. Separated proteins in the gel were transferred to a nitrocellulose membrane, which was incubated for 1 h with 5% bovine serum albumin (Abcam, Cambridge, UK) in 1×TBS solution (Intron Biotechnology, Seoul, Korea) with 0.1% Tween 20 (Sigma-Aldrich, St Louis, MO, USA). The membrane was washed with 1×TBST and incubated overnight at 4°C with the following primary antibodies: anti-FGFR-1, FGFR-2, FGFR-3 and FGFR-4 (1:1,000; Cusabio Technology, LLC, College Park, MD, USA), anti-phospho AKT (S473) (1:2,000; Cell Signaling Technology, MA, USA), anti-phospho ERK1/2 (Thr202/Tyr204) (1:500; Santa Cruz Biotechnology, Santa Cruz, CA, USA), anti-ERK1/2 (1:1000; Abcam, Cambridge, UK), anti-phospho STAT3 (Y705) (1:1,000; Cell Signaling Technology), anti-GAPDH (1:10,000; Abcam, Cambridge, UK), anti-P21 (1:1,000; Cell Signaling Technology), anti-P27 rabbit antibody (1:1,000; Cell Signaling Technology), anti-P53 (1:500; Santa Cruz Biotechnology), and anti-FGF-17 mouse (1:500; Santa Cruz Biotechnology) antibody. After incubation with goat anti-mouse or -rabbit HRP-conjugated antibody (1:10,000; Bethyl, Montgomery, TX, USA) for 1 h at room temperature, the expression of proteins was visualized using WESTSAVE UP (Abfrontier, Seoul, Korea) and developed with Automatic X-RAY Film Processor (JPI Healthcare Co, Ltd., Seoul, Korea).
Flow cytometry
Normoxic hWJ-MSCs treated with rFGF-17 and transfected with siFGF-17 at passage 7 or hypoxic hWJ-MSCs treated with rFGF-17 and transfected with siFGF-17 at passage 10 were harvested and washed with 1×PBS (Intron Biotechnology, Seoul, Korea). Normoxic hWJ-MSCs not treated with rFGF-17 and transfected with negative control siRNA or hypoxic hWJ-MSCs not treated with rFGF-17 and transfected with negative control siRNA were used as respective control groups. Cells were fixed with BD Cytofix Fixation Buffer (BD Biosciences, Piscataway, NJ, USA) and stained with V450 mouse anti-human CD31 (1:20), fluorescein isothiocyanate (FITC) mouse anti-human CD34 (1:20), phycoerythrin (PE)-Cy™7 mouse anti-human CD44 (1:20), V500 mouse anti-human CD45 (1:20), PerCP-Cy™5.5 mouse anti-human CD73 (1:20), PE mouse anti-human CD90 (1:20), APC mouse anti-human CD105 (1 : 20), and V450 mouse anti-human CD144 (1:20) (BD Biosciences) antibodies for 30 min at room temperature. Cells were washed twice with Stain Buffer (BD Biosciences) and analyzed with a FACSVerse™ flow cytometer (BD Biosciences) and Flowjo software (Treestar, San Carlos, CA, USA).
Results
hWJ-MSCs cultured in hypoxic condition showed high proliferation and secretion of FGF-17 at late passages
In this study, we defined passage 6 as early passage, passages 7 and 8 as middle passages, and passages 9 and 10 as late passages for normoxic and hypoxic hWJ-MSCs. When normoxic and hypoxic hWJ-MSCs were cultured from passage 6 to 10, cell numbers of hypoxic hWJ-MSCs were highly maintained from passage 7 to 10 compared with normoxic hWJ-MSCs (Fig. 1A). To investigate the factors related to maintenance of high proliferation of hypoxic hWJ-MSCs, we compared proteins in CM from hypoxic hWJ-MSCs and normoxic hWJ-MSCs at passage 10 using protein antibody array. The signal intensities of proteins that were upregulated or downregulated by at least 2-fold were analyzed in a scatter plot (Fig. 1B). FGF-17 (+5.008 fold), VEGF/VEGFA (+4.117 fold), IGFBP3 (+ 3.340 fold), IL-1 R3/IL-1 R AcP (+2.382 fold), Chordin-Like 1 (+2.220 fold), and EN-RAGE (+2.051 fold) were highly expressed in CM of hypoxic hWJ-MSCs (Fig. 1C, Table 1). We focused on the highest expression of FGF-17 in CM from hypoxic hWJ-MSCs, and confirmed that the absolute amount of FGF-17 in CM from hypoxic hWJ-MSCs was significantly higher than in CM from normoxic hWJ-MSCs (Fig. 1D). Furthermore, expression of FGF-17 in hypoxic hWJ-MSCs-1 and −2 was higher than normoxic hWJ-MSCs-1 and −2 (Supplemental Fig. S1A, S2A). Expression of the receptors of FGF-17 such as FGFR-1, −2, −3 and −4 was confirmed without different expression level in both kinds of cells (Fig. 1E).
FGF-17 of hypoxic hWJ-MSCs had a role in the increase of cell proliferation
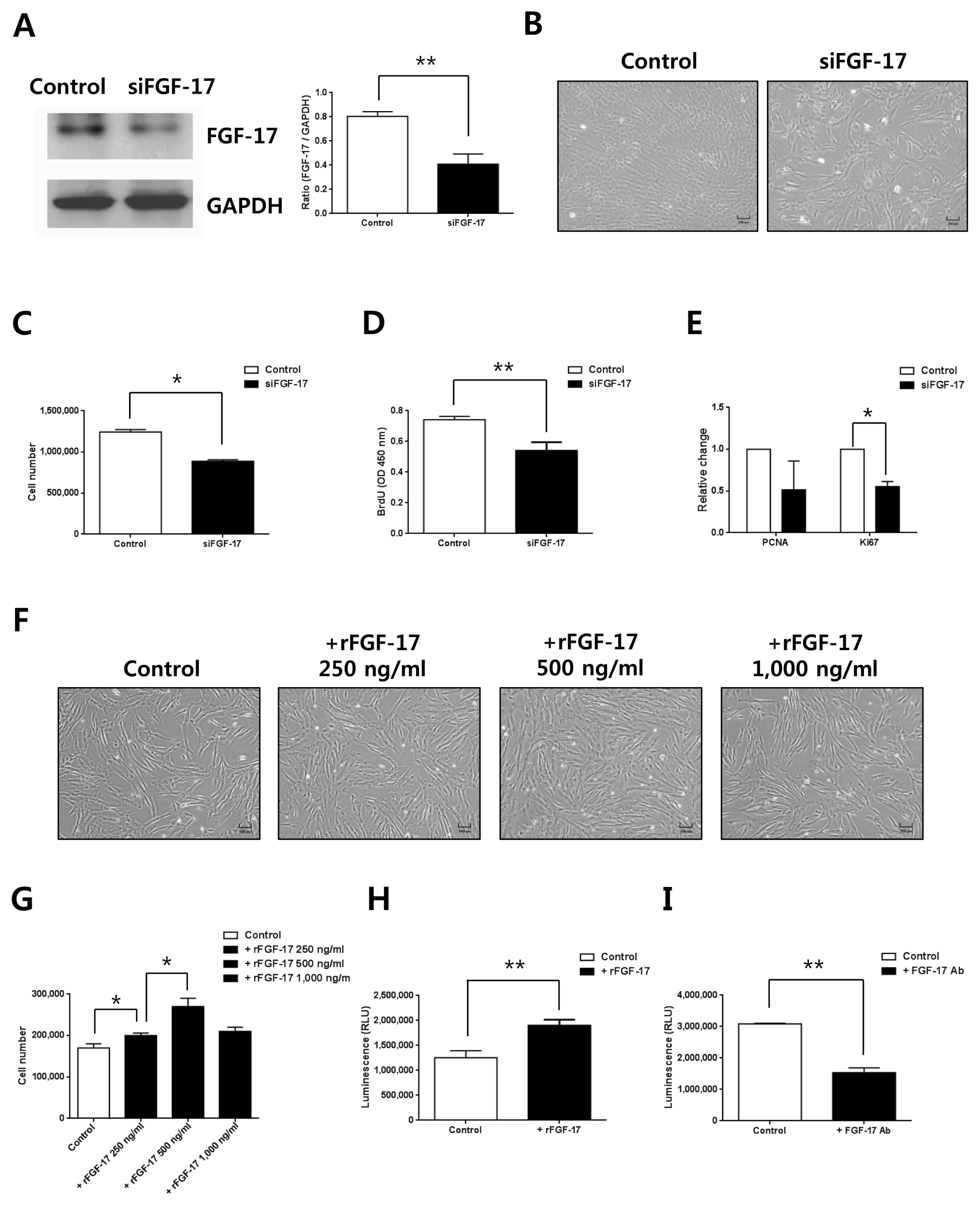
Based on results showing the highest expression of FGF-17 in CM from hypoxic hWJ-MSCs and maintenance of high proliferation of hypoxic hWJ-MSCs at passage 10, we investigated the relationship between FGF-17 and proliferation of hypoxic hWJ-MSCs using siRNA knockdown. Expression of FGF-17 was efficiently decreased when FGF-17 was knocked down in hypoxic hWJ-MSCs by siRNA treatment (Fig. 2A). Furthermore, cell numbers (Fig. 2B, 2C), BrdU incorporation (Fig. 2D), and expression of proliferation-related gene Ki67 (Fig. 2E) were significantly reduced in hypoxic hWJ-MSCs transfected with siRNA of FGF-17. When hypoxic hWJ-MSCs were treated with rFGF-17 for 48 h, cell proliferation (Fig. 2F, 2G) and cell viability (Fig. 2H) were increased in a dose-dependent manner up to 500 ng/ml. Furthermore, there were consistent results on the role of FGF-17 in hypoxic hWJ-MSCs-1 and −2 (Supplementary Fig. S1B~G, S2B~G). When secretory FGF-17 from hypoxic hWJ-MSCs was inhibited by blocking antibody of FGF-17, cell viability was significantly decreased (Fig. 2I).
FGF-17 of normoxic hWJ-MSCs had a role in the increase of cell proliferation
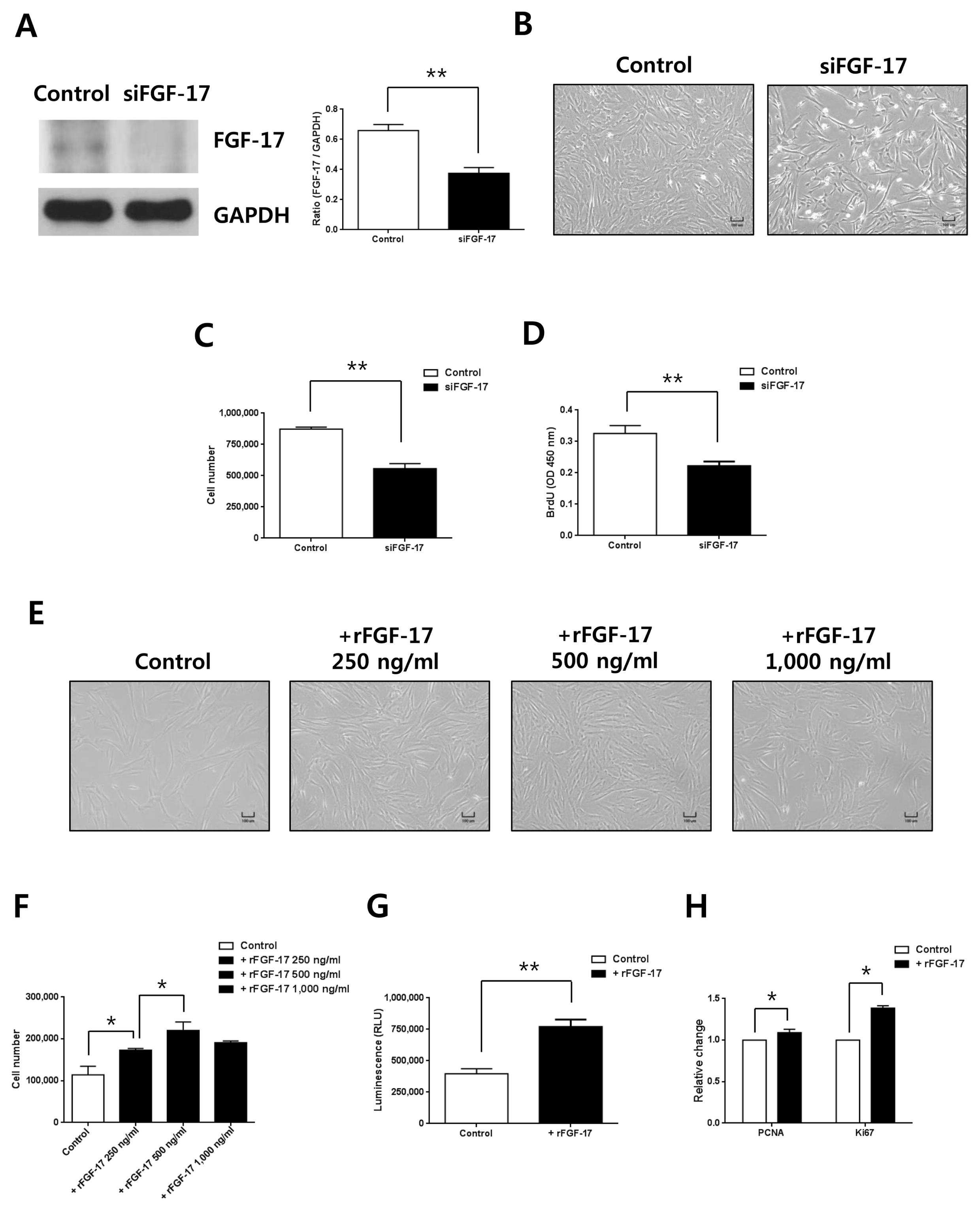
Based on the high levels of FGF-17 in conditioned medium from hypoxic hWJ-MSCs, we investigated the effects of FGF-17 on normoxic hWJ-MSCs at passage 7. Expression of FGF-17 was efficiently decreased when FGF-17 was knocked down in normoxic hWJ-MSCs (Fig. 3A). Cell numbers (Fig. 3B, 3C) and BrdU incorporation (Fig. 3D) of normoxic hWJ-MSCs with transfected with siRNA of FGF-17 were significantly reduced. Treatment of normoxic hWJ-MSCs with rFGF-17 for 48 h increased cell proliferation in a dose-dependent manner up to 500 ng/ml (Fig. 3E, 3F). At 1,000 ng/ml rFGF-17, the proliferation of normoxic hWJ-MSCs was decreased, but without significance (Fig. 3E, 3F). Cell viability and expression of PCNA and Ki67 increased after treatment with 500 ng/ml rFGF-17 (Fig. 3G, 3H). Furthermore, there were consistent results on the role of FGF-17 in normoxic hWJ-MSCs-1 and −2 (Supplementary Fig. S1H~M, S2H ~M).
FGF-17 affects the ERK signaling pathway in hypoxic and normoxic hWJ-MSCs
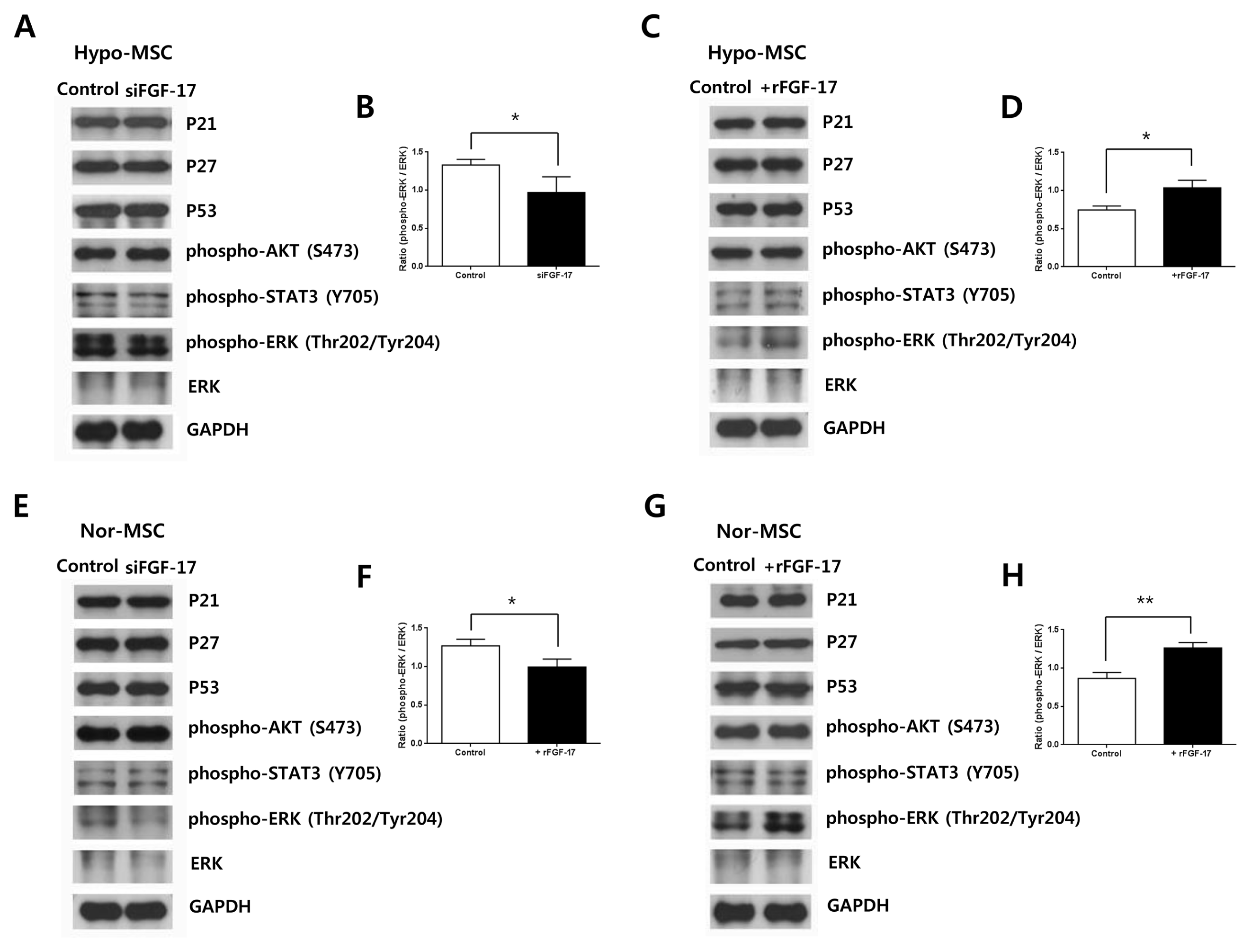
To investigate signal transduction pathways of FGF-17 in normoxic or hypoxic hWJ-MSCs, we analyzed the expression of the following proteins related to cell proliferation or survival: phospho-AKT (S473), phospho-ERK1/2 (Thr202/Tyr204), and phospho-STAT3 (Y705). Knockdown of FGF-17 in hypoxic (Fig. 4A, 4B) or nomoxic hWJ-MSCs (Fig. 4E, 4F) decreased expression of phospho-ERK1/2 (Thr202/Tyr204), but had no effect on levels of phospho-AKT (S473) and phospho-STAT3 (Y705).
Treatment of hypoxic (Fig. 4C, 4D) or normoxic hWJ-MSCs (Fig. 4G, 4H) with 500 ng/ml rFGF-17 for 48 h resulted in increased expression of phospho-ERK1/2 (Thr202/Tyr204), whereas phospho-AKT (S473) and phospho-STAT3 (Y705) were not changed. Based on the relationship between high proliferation and expression of FGF-17 from hypoxic hWJ-MSCs at late passages, we investigated whether FGF-17 was associated with senescence of normoxic and hypoxic hWJ-MSCs. However, expression of senescence-related proteins p21, p27, and p53 was not changed in hypoxic hWJ-MSCs transfected with siFGF-17 (Fig. 4A) and treated with rFGF-17 (Fig. 4C) or in normoxic hWJ-MSCs treated with siFGF-17 (Fig. 4E) and transfected with rFGF-17 (Fig. 4G).
Effects of FGF-17 on characterization of hWJ-MSCs
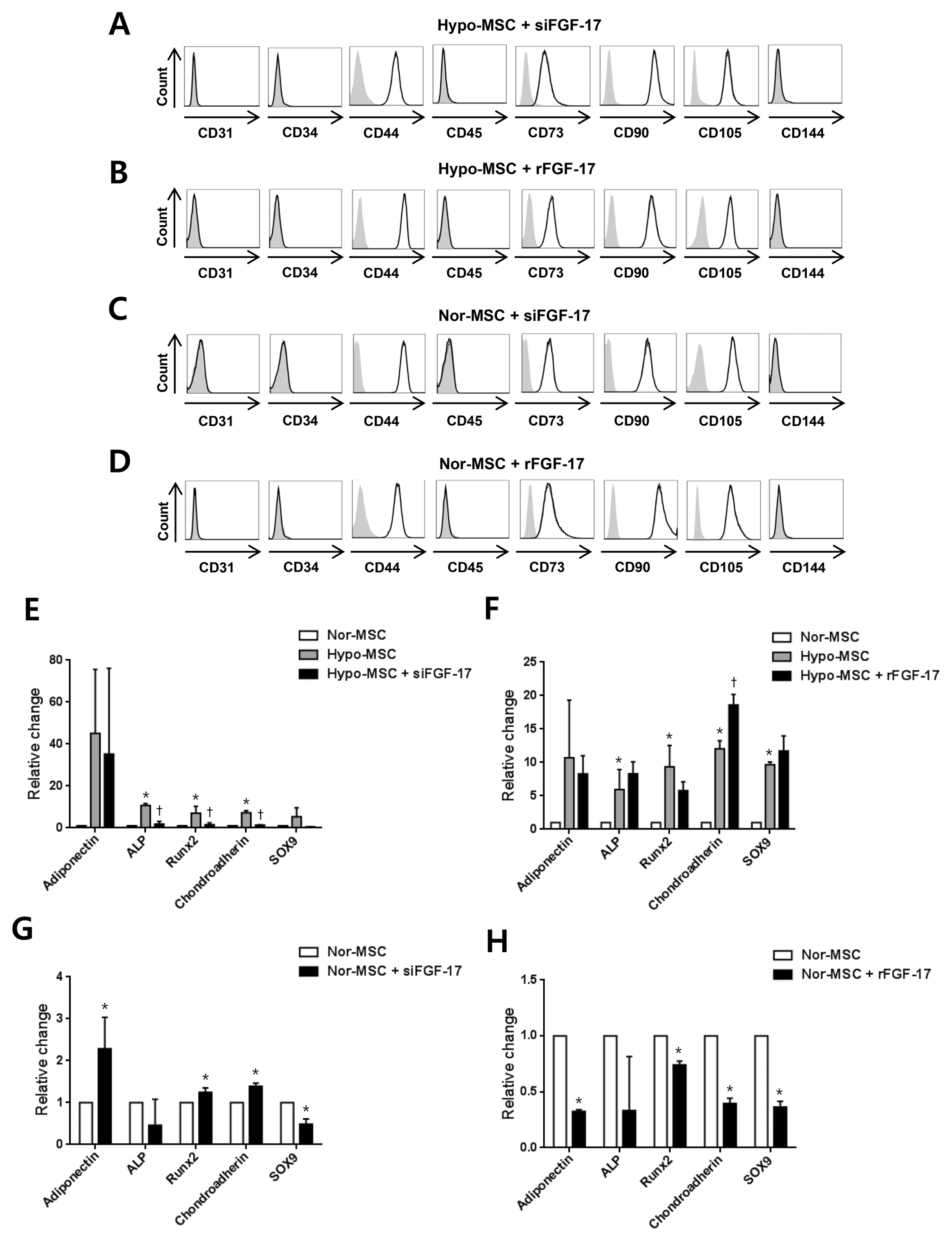
To investigate the effect of FGF-17 on phenotypic characterization of hWJ-MSCs, we analyzed phenotype markers of not only hypoxic hWJ-MSCs transfected with siFGF-17 and treated with rFGF-17 (Fig. 5A, 5B) but also normoxic hWJ-MSCs transfected with siFGF-17 and treated with rFGF-17 (Fig. 5C, 5D). Under both conditions, phenotypes were not changed by FGF-17. The increased expression of osteogenesis-related gene ALP, Runx2, and chondrogenesis-related genes Chondroadherin in hypoxic hWJ-MSCs was significantly decreased by knockdown of FGF-17 (Fig. 5E), and expression of Adiponectin and SOX9 was decreased but without significance (Fig. 5E). When rFGF-17 was treated to hypoxic hWJ-MSCs, Chondroadherin was increased (Fig. 5F). When FGF-17 was knocked down in normoxic hWJ-MSCs, expression of adipogenesis-related gene Adiponectin, osteogenesis-related gene Runx2, and chondrogenesis-related genes Chondroadherin were increased, whereas chondrogenesis-related genes SOX9 was decreased (Fig. 5G). Expression of the adipogenesis-related gene adiponectin, osteogenesis-related gene Runx2, and chondrogenesis-related genes chondroadherin and SOX9 was significantly decreased in normoxic hWJ-MSCs treated with rFGF-17 and expression of the osteogenesis-related gene ALP was decreased without significance (Fig. 5H).
Discussion
In this study we show that increased expression of FGF-17 in CM from hypoxic hWJ-MSCs plays a role in maintenance of high proliferation of human mesenchymal stem cells at late passages.
In general, it is well known that the proliferation and survival of cells decreases with increased passage numbers under normal cell culture conditions (18). The mechanisms related to decreased cell proliferation have been reported to be senescence (18), apoptosis (18), DNA mutation (19), and cell transformation (20). In contrast, it has been reported that hypoxic cell culture conditions not only enhance proliferation and survival of cells, but also influence the secretome of mesenchymal stem cells (10–12). According to our protein antibody array, secretory FGF-17 from hypoxic hWJ-MSCs was highly expressed at passage 10 (late passage), and knockdown of FGF-17 decreased cell proliferation of hypoxic hWJ-MSCs. These findings indicate that FGF-17 from hypoxic hWJ-MSCs plays a role in maintaining high proliferation at late passages. In addition, treatment of normoxic hWJ-MSCs at passage 7 (middle passage) with 250 ng/ml or 500 ng/ml rFGF-17 increased cell proliferation, whereas 1,000 ng/ml rFGF-17 did not further increase proliferation. This may suggest that there is a threshold in the working concentration of FGF-17 for effects on normoxic hWJ-MSCs at the cellular level.
According to protein antibody array, VEGF, IGFBP-3, IL-1 R3, Chordin-Like 1, and EN-RAGE were upregulated at least 2-fold in CM from hypoxic hWJ-MSCs at passage 10 compared with normoxic hWJ-MSCs. It has been reported that VEGF and Chordin-Like 1 are related to increased cell proliferation (21, 22) and these may be additional candidate molecules for the maintenance of high proliferation of hypoxic hWJ-MSCs at late passages. Interestingly, previous studies showed that IGFBP-3 and EN-RAGE play a role in inhibiting cell proliferation (23, 24). It is possible that the positive effect of FGF-17, VEGF, and Chordin-Like 1 overcome the negative effect of IGFBP-3 and EN-RAGE for maintenance of high proliferation in late passages of hypoxic hWJ-MSCs. We will further investigate the roles of these molecules.
In a previous study, FGF-4 and associated activation of the ERK1/2 pathway triggered a switch from self-renewal to lineage commitment in embryonic stem cells (25). In contrast to FGF-4, even though treatment with FGF-17 increased the expression of phospho-ERK1/2 in normoxic hWJ-MSCs, the expression of differentiation genes was decreased. This suggests that FGF-4 and FGF-17 may have different roles in stem cell function even though they are both members of the FGF family. It has been reported that expression of FGF-2, FGF-11 and FGF-23 included in fibroblast growth factor family were regulated by hypoxia-inducible factor (HIF-1α) in hypoxic condition (26–28). For further study, we will investigate the relation between FGF-17 and HIF-1α.
We presumed that the co-relationship between high expression of FGF-17 and maintenance of cell proliferation at late passages might result from inhibition of cell senescence by FGF-17. However, expression of senescence-related proteins (p21, p27, and p53) was not changed in normoxic hWJ-MSCs treated with siFGF-17 and rFGF-17 at passage 7 or hypoxic hWJ-MSCs treated with siFGF-17 and rFGF-17 at passage 10.
Furthermore, under the same conditions the phenotypes of normoxic hWJ-MSCs were not altered and expression levels of differentiation-related genes adiponectin (adipogenesis), Runx2 (osteogenesis), and chondroadherin and SOX9 (chondrogenesis) were decreased by treatment of rFGF-17. These results provide supportive evidence for the use of rFGF-17 as a supplement in culture medium of human mesenchymal stem cells in order to enhance the expansion of cells with stemness.
Interestingly, in hypoxic hWJ-MSCs the expression of differentiation genes was increased (significantly for ALP, Runx2 and chondroadherin; non-significantly for adiponectin and SOX9) compared with normoxic hWJ-MSCs, whereas knockdown of FGF-17 in hypoxic hWJ-MSCs decreased the expression of these molecules (significantly for ALP, Runx2, and chondroadherin; non-significantly for adiponectin and SOX9). This suggests that the effects of FGF-17 on differentiation-related genes are different between normoxic and hypoxic hWJ-MSCs and further studies will be necessary to investigate the mechanisms underlying these differential activities.
In conclusion, FGF-17 from hypoxic hWJ-MSCs plays a role in maintenance of their high proliferation at late passages through the ERK1/2 pathway. And it will be useful as a supplement for the maintenance of proliferation and expansion of hWJ-MSCs under long-term culture conditions.
 XML Download
XML Download