

 PDF
PDF Citation
Citation Print
Print


Introduction
The liver is one of the most complex organs in the body. Its properties of regeneration have intrigued scientists from around the world. It is composed of many cells such as hepatocytes, biliary epithelial cells, Kupfer cells, stellate cells, and hepatic progenitor cells. The most prominent of these cells are hepatocytes, composing nearly 95% of all cells in the liver. The main function of hepatocytes is to regulate metabolism including glycogenesis and glucogenesis. The other main functions of hepatocytes include their ability to synthesize urea as well as lipids (1). These cells are often mononucleated or binucleated. The remaining 5% are biliary epithelium cells also known as cholangiocytes (2). Cholangiocytes line the bile ducts in the intrahepatic biliary tree as well as the extrahepatic biliary tree. Even though these epithelial cells are not present at a high percentage, they retain a very important role in the liver. Cholangiocytes modify the bile produced by the hepatocytes so that it does not cause harm or injury to the liver. It then transports this bile from the liver to the gut through the biliary tree (3). Therefore, it could be hypothesized that the main function of cholangiocytes is to maintain gradient homeostasis within the liver (4).
The interruption of such a function can cause cholangiopathies. The extrahepatic and intrahepatic bile ducts are only a small portion of the liver. Despite this, the most common treatment for cholangiopathies is a full liver transplantation. Other non-invasive, inexpensive, and quicker methods of treatments have yet to be used for the treatment of cholangiopathies. This could be partly due to the limited knowledge obtained on the mechanism of actions in cholangiopathies. Throughout the years, many researchers have conducted studies giving rise to many differentiation protocols of iPSCs cells to cholangiocytes in 3D culturing systems. Other scientists have focused on the making of artificial bile ducts to replace the native bile duct. In this review, the basic features of cholangiocytes, cholangiopathies, and their current treatment methods as well as currently available research findings will be highlighted.
Go to : 
Developmental Biology of the Bile Ducts
The liver develops differently among species. There is a considerable difference between the time of liver development in humans and animals. Studies done on Zebrafish show that gut cells derive from endoderm cells that move to the midline; however, whether the liver stems from these cells as well is unknown. In mouse and chick models, the liver develops from the anterior and posterior epithelial sheet of endodermal cells (5). As early as embryonic day 8.5, the liver bud starts to develop, and hepatic differentiation through the expression of albumin can be detected. The intrahepatic and extrahepatic bile ducts develop differently at different times. The extrahepatic duct includes several different ducts: the common hepatic duct, the cystic duct, and finally the common bile duct. In every species except the rat, the extrahepatic duct also includes the gallbladder (6). Not much is known about the developmental cascade and signals that lead to the extrahepatic bile duct formation. However, its generation is thought to occur through the caudal part of the ventral foregut endoderm. The intrahepatic bile duct forms through the diverticulum of the foregut endoderm and the septum transversum, which sends signals to initiate its formation (7). FGF from the cardiac mesoderm, BMP from the septum transversum mesenchyme, and Wnt signaling are very important for correct formation of the liver bud. Hepatoblasts, the progenitor cells of the liver, detach and migrate to the septem transversum mesenchyme, and the activation of the sonic hedgehog (SHH) pathway allows for these cells to proliferate prior to differentiation. However, after E11.5, at around E13, the SHH levels stop being detected and differentiation to hepatocytes or bile ducts, depending on the location of the hepatoblasts, starts to occur. Hepatoblasts in the parenchyma will differentiate into hepatocytes whereas hepatoblasts near the portal mesenchyme will become cholangiocytes through the help of the mesenchyme signaling. These hepatoblasts will undergo different changes to form the ductular structure of the bile duct. This formation can be characterized in four steps. The first step consists of fibroblast lining up into a single monolayer surrounding the portal veins. This primary structure is called the ductal plate. The second step consists of the duplication of the ductal plate followed by the third step, dilation, which forms the primitive tubular structure. These tubular structures will integrate into the portal mesenchyme. In the third step, some of these hepatoblasts will not undergo the dilation step and therefore need to be removed through apoptosis. When these cells are removed, primitive tubular structures will undergo the fourth and final step by undergoing further morphological changes and branching to form the biliary tree (8). Cholangiocytes will form a 3D structure with a lumen and apicobasal polarity. Cholangiocytes lining the intrahepatic bile duct share the same origin as hepatocytes as they are both a result of hepatoblast differentiation. However, extrahepatic bile ducts’ cholangiocytes share their origin with the duodenum and pancreas as they are both derived from endodermal cells (6). The most important signals that have been reported to be involved in bile duct formation are TGFβ, Notch, Wnt, and FGF signaling. Disruptions in these signals during bile duct formation or biliary differentiation can cause duct malformation, the most common being Alagille’s syndrome, which is caused by mutations in Jag-1 or Notch2 (2). Others will be discussed further in a separate section. Cholangiocytes will differ in size depending on their location post differentiation. Small cholangiocytes will line small bile ducts and are around 9 μm in diameter. These cholangiocytes, similarly to hepatocytes, are diploid cells and have a high nucleus to cytoplasm ratio compared to large cholangiocytes, which as the name suggests, are the opposite of small cholangiocytes. Large cholangiocytes line big bile ducts, commonly the extrahepatic bile duct, are larger than 9 μm in diameter, have a single nucleus, and a low nucleus to cytoplasm ratio. Another difference lies in the shape of these cells. Small cholangiocytes have a cuboidal morphology. In contrast, large cholangiocytes have a columnar morphology (9). While both large cholangiocytes/ducts and small cholangiocyte/ducts are found in all species, the number of cholangiocytes lining the large ducts differs among species. Similarly to humans, rodents have 4 to 5 cholangiocytes lining the small bile ducts found in the intrahepatic part. However, the difference lies in the large hepatic ducts where humans have 40 cholangiocytes lining them whereas rodents have up to 15. This could be due to the difference in liver size between rodents and humans. Furthermore, all cholangiocytes possess a primary cilium. However rat, mouse, and human cilia differ in size. Every cilium has the 9+0 ax-oneme pattern. However, in rats, cilia size and cell size have a positive correlation and are proportionate. In the mouse, the cilia grows to a maximum length of 10 μm, and in humans, the length of the cilia has not yet been characterized in the different bile ducts (6).
Go to :
Signaling of the Bile Duct and Organogenesis Defects
As we previously mentioned, TGFβ, Notch, Wnt, and FGF signaling are important in the formation of the intrahepatic bile duct. Mutations or disruptions in these leads to a malformation of the bile ducts. However, some transcription factors such as Forkhead Box (10) A1 and A2 can also have an effect by inhibiting the initiation of liver development altogether (11). Notch initiates the process of the progenitor cells to differentiate towards a biliary fate for cholangiocyte generation. Notch can also have repair properties on the biliary tree. Notch 1, 2, 3, and 4 are all receptors encoded by the notch genes that interact with ligands such as Jagged-1, 2 and Delta like 1, 3, and 4. When an impairment between the interactions of Notch2/Jagged-1 occurs, Alagille syndrome occurs. This syndrome deals with the branching of the biliary tree, where faults can be seen during the developmental period (7). Notch signaling also activates a number of transcription factors which in turn helps the differentiation of the hepatoblasts into biliary epithelium cells as well as formation of the intrahepatic bile ducts. Hepatic nuclear factor (HNF) 6 is one of the most important transcription factors that has been demonstrated through HNF6 null mice. These mice show different symptoms such as the absence of a gallbladder as well as abnormalities in developing intra and extra hepatic bile ducts. Inactivation of the transcription factor HNF1β causes similar symptoms; however, unlike HNF6, the gallbladder is present, but clear abnormalities in this organ can be seen through the presence of severe jaundice as well as the intrahepatic bile duct (12). HNF6 also controls the delamination of hepatoblasts and their migration to form bile ducts. Inadequate levels of HNF6 can cause inhibition of the migration of the hepatoblasts and a loss of differentiation. Furthermore, Notch1 aids the differentiation of hepatoblasts to cholangiocytes through the repression of hepatic differentiation of the hepatoblasts. However, Notch1 does not regulate the differentiation of hepatoblasts to biliary epithelium cells. In fact, in a study reported by Tchorz et al., it was shown through N2ICD/AlbCre mice, in contrast to control mice, that cells expressing Notch2ICD survive into adulthood showing the important of Notch2 (13, 14). Another study performed with the knockout of the RBP-Jκ gene, which is controlled by the canonical Notch pathway, shows a decline in the number of cells ready to commit to a biliary fate, further highlighting the importance of the Notch pathway (14, 15). The presence and absence of Wnt signaling is also important during liver specification. During initiation of the specification of the liver, Wnt must be switched off. After the initiation process is finished, Wnt can then be switched on. Wnt signaling oversees hepatoblast proliferation and when inhibition occurs, liver hypoplasia (abnormal liver development) can appear. β-catenin is a protein codded by the CTNNB1 gene and works downstream of Wnt signaling to promote proliferation. Studies of Wnt signaling have been reported in chicks and mice models. In chicks, the pro-proliferative action of Wnt signaling was shown, which was further proven in the mice model through the deletion of the CTNNB1 where liver hypoplasia was observed. Not only do they play an important role in regulating the normal developmental stages of the liver, but studies also show that the Wnt/β-catenin pathway also promotes biliary differentiation where a disruption in this pathway has led to the inhibition of this process. Working with the Wnt/β-catenin pathway are hepatocyte growth factor (HGF) and fibroblast growth factor (FGF). These factors are important in normal organogenesis of the liver. When they are deleted, the liver does not grow to its full size. Studies by Hussain et al. also showed the importance of the Wnt pathway by studying the cell reaction to an inhibition or addition of Wnt. The group found out that under Wnt-free conditions, cell viability was inadequate, proliferating cell nuclear factor (PCNA) was negative in over 95% of the cells, and Wnt also ameliorates the proliferation and survival capacity of bile duct cells. These effects show the importance t of the signaling in bile duct morphogenesis (16). TGF-β has a dual role in the liver where it controls its growth as well as the fate of hepatoblasts to turn into biliary epithelium cells. Studies done on mice models that are heterozygous for the SMAD2/3 (signaling mediators) show that mutations or disruptions can cause liver hypoplasia. Other signaling pathways can be involved in ductal formation such as the hippo pathway signaling. However, what the mechanism of action is and whether it primarily controls biliary differentiation is still unknown and research is still ongoing. However, the distal mediator called Yap1 can provide some evidence where the deletion of this gene can cause bile duct formation to fall through and the levels of serum bilirubin to increase (17).
Go to :
Cholangiopathies
In the United States. Around $400 million is spent for liver transplantations, which accounted for 16% of all transplantations between the years 1988 and 2014 due to cholangiopathies (18). Cholangiopathies can be grouped into different groups depending on the cause of the cholangiopathy. However, many of these categories can overlap. Cholangiopathies are often hard to diagnose and treat as they usually involve multiple organs and systems. Therefore, establishing a method to treat just one system is often difficult. This poses a problem due to many cholangiopathies being discovered but only transplantation as the preferred method of treatment. As previously mentioned, Notch signaling is important in many stages during bile duct generation. In the mouse model, mutations in the Jagged 1 ligand and notch2 receptor cause Alagille syndrome (AGS). AGS not only affects the liver through bile duct paucity as well as cholestasis but can also affect the cardiac, renal, skeletal, and ophthalmic systems (19, 20). Cholangiocarcinoma (CCA) accounts for 3% of all gastrointestinal cancer, ranks first for biliary malignancy, and second for hepatic malignancy. With a poor prognosis of 24 months median survival after diagnosis and only 10% of patients surviving five years after diagnosis, the only therapy option is surgery. CCA is divided into three subtypes: intrahepatic cholangiocarcinoma (iCCAs), perihilar cholangiocarcinoma (pCCAs), and distal cholangiocarcinoma (dCCAs) (21). CCA does have etiological evidence. In Asian countries, hepatolithiasis is considered to be a risk factor whereas in the West, primary biliary sclerosis is the most common predisposing factor. Furthermore, choledochal cysts (bile duct cysts) are a higher risk factor in Asian countries than in the West. Among hepatobiliary flues, infections with Opisthorchis viverrini as well as Clonorchis sinensis are known to cause CCA due to their ability to cause inflammation through infections. Interestingly, diseases such as choledocholithiasis, inflammatory bowel disease, and cholelithiasis can promote CCA development. The main reason for this is due to inflammation playing a major role in how CCA arises. During inflammation, inflammatory cytokines are released into their surroundings causing an upregulation in the synthesis of nitric oxide through nitric oxide synthase. This in turn results in breaks in a single strand or both strands of the DNA. Normally, this would not cause a problem. However, a result of this excess leads to DNA repair enzymes being inhibited and therefore the DNA breaks do not get fixed (22). Primary biliary cirrhosis (PBC) is considered to be rare with an end result as severe as liver failure. PBC is an autoimmune disease that is found in women in most cases. PBC can be derived from several extrahepatic immune diseases such as autoimmune thyroid disorders and others. It is unique in that it affects small bile ducts and is a risk factor for cholangiocarcinoma (23). Primary sclerosing cholangitis (PSC), unlike PBC, is idiopathic meaning that there is no known reason for why it occurs. Unlike PBC, it does not primarily affect women but is mostly found in males where 75% of PSC patients also have irritable bowel syndrome (IBD). While the exact relationship between PSC and IBD is not known, it is hypothesized that the infection caused by IBD is what causes PSC as previously mentioned. Another difference between PSC and PBC is that PSC affects large and extrahepatic bile ducts. The only similarity between the two is that they are both a risk factor for cholangiocarcinoma (3). Having discussed adulthood cholangiopathies, childhood biliary atresia (BA) and familial intrahepatic cholestasis (PFIC) are also of importance. BA affects 1 in 10–19,000 in Europe and North America. This number is quite low however, it is the most common cholangiopathy. It is characterized by neonatal jaundice and malfunction of the canalicular-bile duct continuum (24). Ductal plate malformation (DPM) is characterized by necroinflammation and intrahepatic bile duct destruction as well as an accompanying fibrotic reaction. During the 8th week of gestation, the bile ducts start to form. In a previous section, organogenesis was mentioned. However, DPM is the result of a disruption or a failure in one of the aforementioned steps. DPM is also associated with autosomal dominant polycystic disease (ADPKD) and is thought to be a result of PKD1 as well as PKD2 mutations. CFTR holds an important function being a cAMP-activated Cl− channel. A disruption in this function can cause pathological results, and a deficiency in CFTR can cause pulmonary or pancreatic disease. Its other functions include working hand in hand with Na+ channels, the Cl/HCO3 exchanger also known as AE2, and potassium channels. Cystic fibrosis is the result of an accumulation of mutations in the CFTR gene that affects cholangiocytes and is therefore called ‘cystic fibrosis-associated cholangiopathy’. Usually, when an insult is sprung upon bile ducts, proliferation is initiated and can replace injured cholangiocytes. However, during cholangiopathies, cholangiocytes are lost at a faster rate than regeneration happens. When this happens, cholangiopathies are described as late-stage and ‘terminal’ (25).
Go to :
Current Treatment
At present, there are no cures for cholangiopathies except for liver transplantation. However, liver transplantation has many disadvantages, such as being a risk for immune rejection. However, the biggest disadvantage is the shortage of liver donors and the soaring number of patients needing liver transplantation. The only drug therapy in current use is through treatment with Ursodeoxycholical acid (UDCA). It is usually administered at a dose of 15~20 mg/kg/day. However, a misdosage, especially in the higher spectrum, can cause serious and unwanted outcomes. Furthermore, there is insufficient data showing that UDCA can increase the survival time of patients suffering with cholangiopathies. In addition, since cholangiopathies are often the result or a consequence of many systems failing together, targeting all of them together can prove inefficient (26). Glucocorticoids such as budeosinide have been used for the treatment of PBC at a dose of 6~9 mg daily; however, safety issues such as the effects of long term administration have limited their clinical use. T-cell-targeted therapies have also been brought up as an option. While many are still currently in clinical trials, Ustekinumab, a monoclonal antibody that works in a negative manner in the Il-12 and IL-13 shared p40 subunit showed no effects (27). It is therefore of critical importance to find a solution to this issue.
Go to :
Advances in Regenerative Medicine
Many efforts in the present are being put into generating cholangiocytes for drug screening and cell therapy. Artificial bile ducts seeded with or without cells have also been produced and tested in vivo for the treatment of cholangiopathies. However, much research is yet to be done. Human induced pluripotent stem cells are a good cell source, having advantages such as self-renewal capacity and pluripotency. Pluripotency aids in this research as researchers can mimic the way native bile ducts are formed to produce cholangiocyte-like cells. Through the use of 3D bioprinting, scaffolds can be made to resemble the native bile duct with high specificity. The advantage is the ability to control the thickness and length of the artificial bile duct, so it can be tailored for individual patients in different cases. Reprogramming of iPSCs cells have also been done by infected iPSC-derived hepatic progenitor cells using small hairpin RNA (shRNA) against SOX17 (28).
Sampaziotis et al. (29) published two papers on scaffolds and cholangiocyte-like cell induction. Their first paper published in 2015 consisted of adding human induced pluripotent cells (hIPSC) to cholangiocyte-like cells in five steps. Their first step was from hIPSC to definitive endoderm by the addition of Activin A, FGF2, BMP4, Ly294001 and conditioned differentiation media. The second step was to differentiate to foregut progenitor using activin A and RPMI. The third step was hepatoblast differentiation through SB-431542, BMP4, and RPMI. The fourth step was to induce cholangiocyte progenitors using FGF10, retinoic acid, Activin A, and RPMI. Their final step was to produce cholangiocyte-like cells in 3D culture using Matrigel and William’s media supplemented with EGF. These cholangiocyte-like cells were able to express the cholangiocyte marker cytokeratin 19 (CK19) as well as cytokeratin 7 (CK7) and cystic fibrosis transmembrane receptor (CFTR), which are key biliary markers in the induction of cholangiocytes. Of importance, they were able to generate cholangiocyte-like cells from patients with cystic fibrosis. Through these organoids, they tested the function of the experimental drug VX809 and were able to generate positive results that were never seen before (29). In their more recently paper published in 2017 (30), instead of differentiating, they produced organoids called extrahepatic cholangiocyte organoid (ECO) from cholangiocytes extracted from the extrahepatic biliary tree of healthy donors. After organoid formation, they seeded them onto collagen tubes and transplanted them into the extrahepatic biliary injury (EHBI) mouse model. The isolated donors’ cholangiocytes were cultured in a mixture of 66% William’s media and 33% Matrigel with the addition of nicotinamide, sodium bicarbonate, 2-phospho-L-ascorbic acid trisodium salt, sodium pyruvate, glucose, HEPES, ITS+premix, dexamethasone, Glutamax, pencillin/streptomycin, EGF, R-spondin, and DKK-1. These ECOs expressed the biliary markers CK7, CK19, SRY-box 9 (SOX9), and CFTR. Next, due to their inability to induce an immune response, they used polyglycolic acid (PGA) scaffolds as they are biodegradable and easy to use and manipulate. When the ECOs were seeded on the scaffold, they retained their biliary marker expression, and when transplanted into the EHBI mouse model (NSG mice with a compromised biliary tree due to an incision in the gallbladder wall), they were able to show regeneration capacity as all the mice survived and reconstructed the counterpart almost identically to the native one with retained biliary marker expressions. One of the most important observations was that there was no tumour formation, which is advantageous to human induced pluripotent stem cells known for tumour induction (30).
Dianat et al. (31) also developed a differentiation protocol to induce cholangiocytes from human pluripotent stem cells. Both Sampaziotis et al. and Dianat et al. similarly mimicked the organogenesis of the native bile duct by using the same chemicals to go through differentiation from a definitive endoderm to hepatic endoderm to a hepatoblast and finally to cholangiocytes. The media used to differentiate hepatoblasts to cholangiocytes consisted of William’s E/Ham F12 1:1 with linoleic acid-Albumin, 3,3′,5-Triiodo-L—thyronine, IU insulin, Vitamin C, human apo-transferrin, and sodium pyruvate. After 24 hours, HGF and EGF were introduced into the medium for three days following the addition of interleukin-6 (IL6) for another three days. In the final stage of differentiation, the cells expressed biliary markers such as CK19, aquaporin 1 (AQP1), and CFTR as well as CK7 and other markers. They also stained positive for acetylated α-tubulin showing cilia formation in the cholangiocytes generated from human induced pluripotent stem cells (hESC-Chol) (31).
Many studies have been conducted on differentiating human induced pluripotent stem cells through stage-step differentiation as previously mentioned. Ogawa et al. (32) published a paper using a similar differentiation process. However, they co-cultured hPSC derived hepatoblast with OP9 stromal cells at the cholangiocyte differentiation step with the addition of HGF, EGF, and TGFβ in a mixture of 1.2 mg/ml collagen type-I and 40% Matrigel. This resulted in ductal plate or/and cysts forming. They expressed CK19, CFTR, secretin receptor (SCTR), and stained positive for the presence of cilia. To test the function of these cells, they transplanted them into the mammary fat pads of non-obese diabetic–severe combined immunodeficient–interleukin 2rγ−/− (NOD-SCID-IL2rγ −/−; NSG) mice at a cell count of around 106 cells using a Matrigel plug. Eight weeks post-transplantation, the biliary markers were retained, and structures typical of biliary fate were seen. Similarly to Sampaziotis et al., they attempted to induce cholangiocyte cysts from CF patients. They discovered that until the hepatoblast generation step, there was no difference between the normal groups and the CF groups. During cholangiocyte differentiation, however, cysts could not be generated. Hence, they tested the effect of VX-809 on cyst formation. Interestingly, unlike Sampaziotis et al., there was no correction in cyst formation, but there was upregulation in the amount of CFTR that was accumulated on the apical part of the lumen (32).
De Assuncao et al. (33) generated cholangiocytes from human induced pluripotent stem cells following the definitive endoderm-hepatic specification-hepatic progenitor-cholangiocytes method. To generate cholangiocytes from human induced pluripotent stem cell derived hepatoblasts, they used H69 medium supplemented with 100 μg/ml TGFβ and a collagen.-type I and Matrigel mix. This method generated cyst like and branch like formations typical of 3-dimensional (3D) culturing of cholangiocytes. They were also able to express markers such as CK19, CK7, CFTR, and anion exchanger 2 (AE2). They produced a mouse model by distal common bile duct clamping and dissection of the gallbladder. Next, they injected 106 cells. Three days later, bile duct structures emerging from these cells were seen as well as a biliary marker expression showing the regeneration potential of this protocol (33).
Apart from human induced pluripotent stem cells, Kido et al. (34) published a paper in 2015 after using carboxypeptidase M (CPM) as a marker for liver progenitor cells (LPC). They then differentiated these CPM+ LPC into cholangiocytes through the use of a collagen/Matrigel mix, R-spondin 1, and WNT3a. This resulted in the generation of bile duct epithelial cells after seven days of culture expressing aquaporin 1 (AQP1) and F-actin in the apical membrane and CD49f in the basolateral membrane. They also expressed CK7 and CFTR were expressed at a higher level than cholangiocytes derived from human induced pluripotent stem cells (34).
Researchers have also used different materials to produce artificial bile ducts to replace the native bile. Aikawa et al. (35) used a bioabsorbable patch consisting of polycaprolactone and polyglycolic acid. They performed surgery on pigs and checked for results five weeks and four-months post implantation. The results were positive with total bilirubin, aspartate, and alkaline phosphatase levels going back to normal 4 months after surgery. Furthermore, the patch dissolved, and the artificial bile duct was indistinguishable from the native bile duct. The patch site also stained positive for CK19, showing regeneration of the bile duct with a population of bile duct epithelial cells (35).
Perez Alonso et al. (36) also replaced the common bile duct with artificial collagen tubes coated with 2% agarose hydrogel. They implanted these artificial bile ducts in 40 guinea pigs and examined them at 4 weeks, 3 and 6 months post implantation. At 6 months, all the guinea pigs were still alive as well as total bilirubin, ALP, and AST levels identical to those of the control group. Furthermore, reepithelization of the artificial bile duct was noted, although they did not establish what had caused this reepithelization. Their results further proved the regeneration potential of artificial bile ducts (36).
Unlike the previous two methods that included artificial bile ducts that were not seeded with cells, Miyazawa et al. (37) conducted research on the difference between transplanting artificial bile ducts with autologous bone marrow cells (BMC) and without. A scaffold was made of polycaprolactone and polylactic acid, reinforced with polylactic fibers, and transplanted into the common bile duct of hybrid pigs. At 6 months, the pigs had gained weight and acquired normal levels of ALT, AST, and total bilirubin. They also showed no signs of jaundice. Reepithelization was also seen in this case from the hepatic and duodenum ends. However, the degree of reepithelization between the groups with and without cells was not significant. Furthermore, the cells repopulating the artificial bile duct were stained positive for CK19 (37).
Our laboratory published a paper in 2017 (38) about bile duct regeneration using three-dimensional bioprinting through MRI imaging (Fig. 1). 3D and 2D MRCP images were taken of a rabbit’s bile duct and used as a template to produce an almost identical tubular scaffold. The scaffold was made of polyvinyl alcohol (PVA) filaments 1.75 mm in diameter by an in-house printer called the RepRap Mendel 3D printer. It was then coated with polycaprolactone (PCL), which when dried formed a coating on the PVA. The PCL-coated PVA was then immersed in dichloromethane to remove the PVA template. The end result was a tubular structure made from PCL almost identical to the rabbit’s native bile duct obtained from MRI imaging. The artificial bile duct was anastomosed to the proximal and distal ends of the extrahepatic bile duct (EHBD). Three days after implantation, MRI was performed again on the surviving rabbit. The images showed a slight stricture; however, bile flow was intact and flowing normally with a smooth interconnection between the artificial and native bile ducts. H&E of the tissue containing the artificial bile duct was also performed after sacrificing the rabbit showing an appropriate healing process as well as regeneration by the presence of cells morphologically similar to biliary epithelium cells (38).
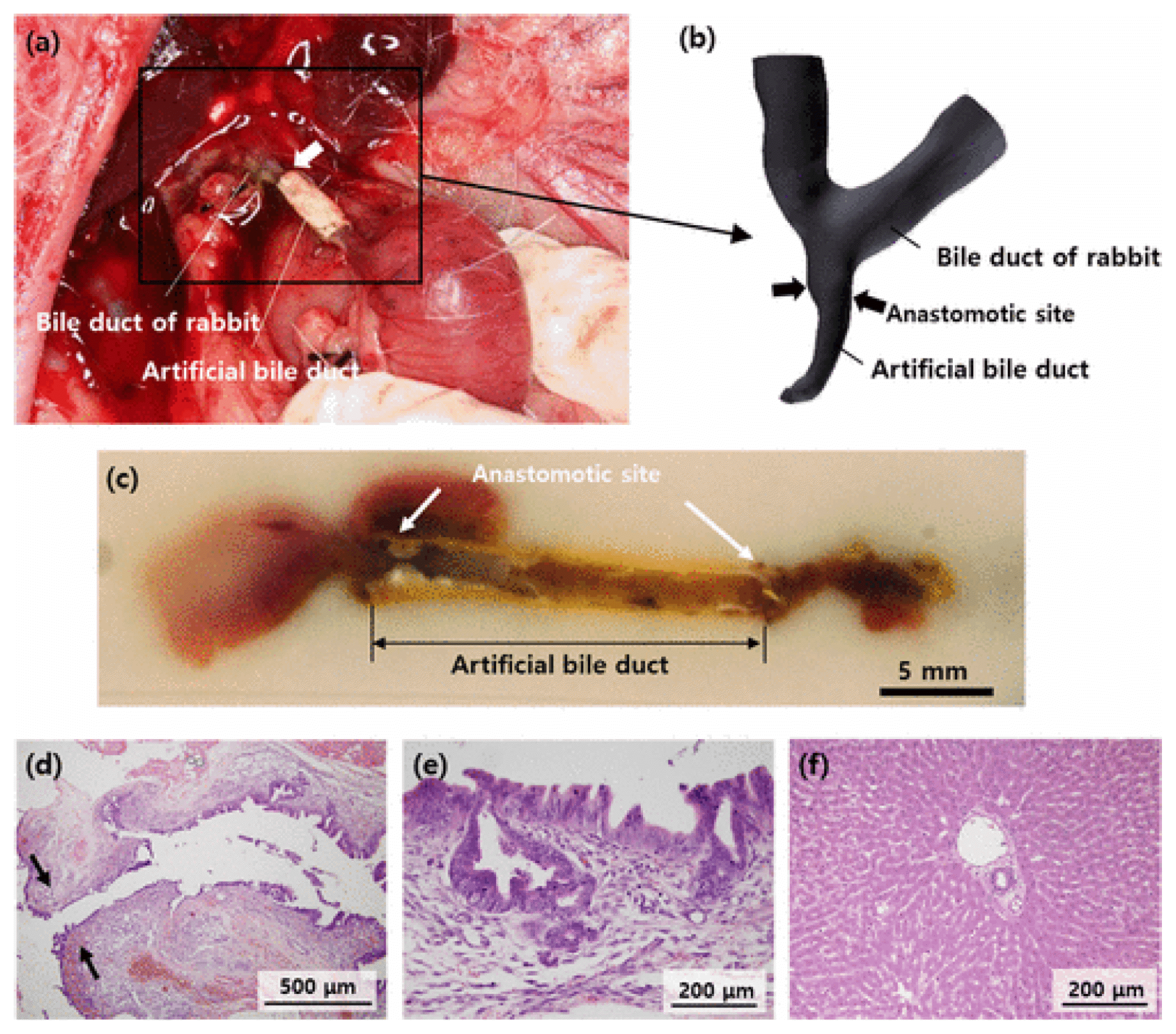 | Fig. 1This figure and legend were adapted courtesy of the authors of Park et al. for use in this manuscript. (a) Photograph of the anastomosed bile duct. (b) The 3D-reconstructed MRCP image shows the anastomotic site and both the dilated intrahepatic bile ducts of the rabbit. (c) Photographs of the sectioned tissue containing the replaced artificial bile duct. Histopathological examination of the rabbit bile duct and liver. (d) Bile duct wall adjacent to the anastomosis site (black arrows). (e) Mucosal epithelium inside the bile duct wall showing normal regenerative atypia. (f) Liver parenchyma with well-preserved hepatic lobules and portal area.
|
We also developed a two-dimensional culture system to differentiate human chemically derived progenitor cells (hCdH) into cholangiocytes. hCdH were derived from hepatocytes obtained from patient livers through the perfusion method. After hepatocyte isolation, hCdH were generated through a cocktail mix of HGF, CHIR, and A83-01, a TGFβ inhibitior. These cells are then plated on 0.1% gelatin coated dishes and differentiated into cholangiocytes using DMEM/F12 supplemented with 10% fetal bovine serum, 1X ITS, 10 mM nicotinamide, 0.1 mM dexamethasone, 75 ng/ml EGF/HGF, 7.5 μM CHIR, and 10 μM sodium taurcholate hydrate on days 10~14.
The medium was changed every day. Two weeks after differentiation, these cells formed clusters or a ductal branch (Fig. 2). They also had a higher expression of the biliary markers CK7, CK19, AE2, CFTR, and AQP1 as seen through RT-qpcr in comparison to their negative control hCdH (Fig. 3). CK7 and CK19 were found three-times higher, AE2 and AQP1 twelve-times higher and CFTR was expressed as much as twenty-times higher in hCdH-Chols. This was further proved through confocal microscopy (Fig. 4). Fluorescein diacetate was also performed to check for metabolic activity. Thirty minutes after FD removal, these cells were able to retain fluorescence showing functional abilities (Fig. 5).
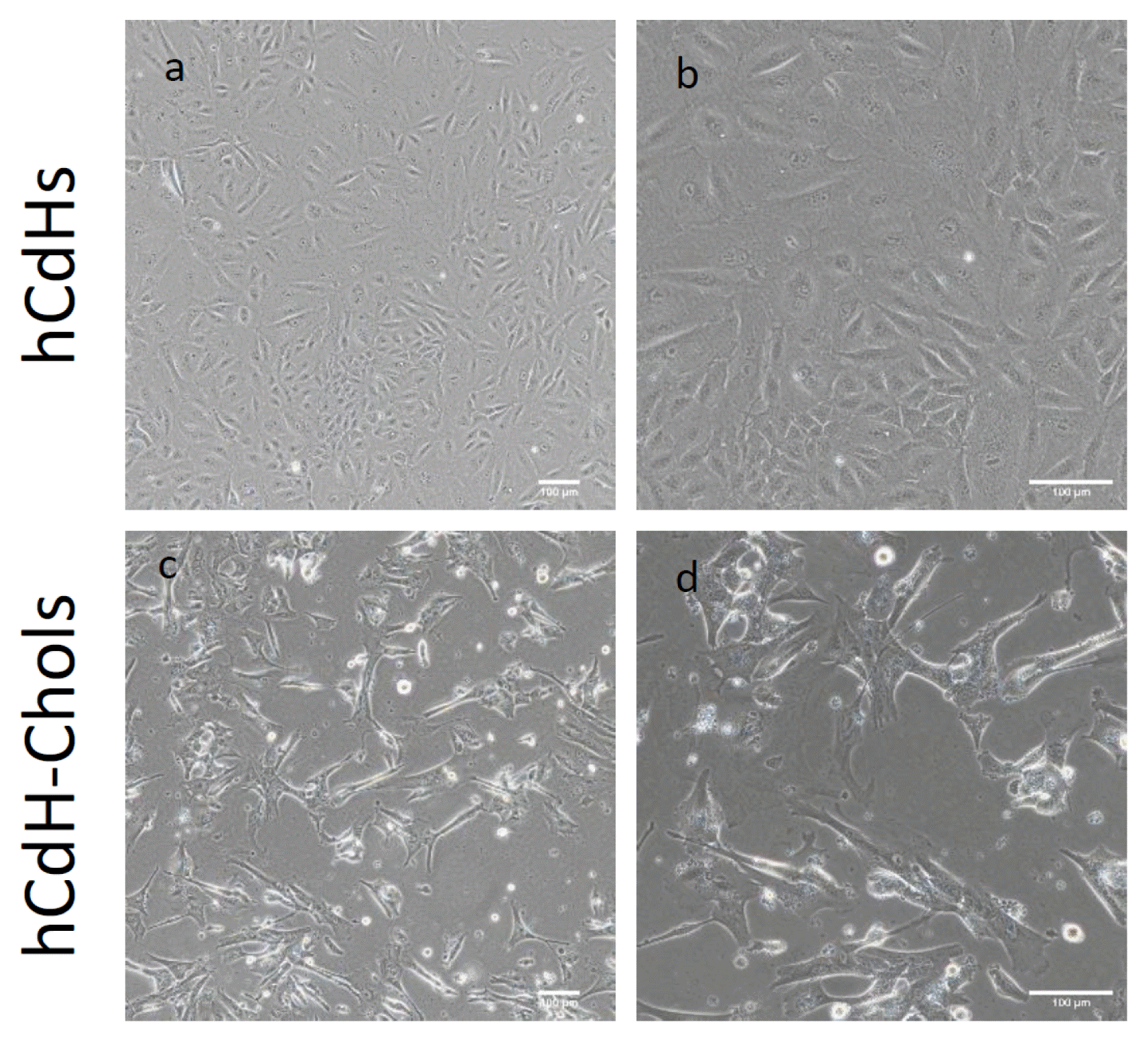 | Fig. 2Microscopy analysis of human CdH and human CdH derived cholangiocytes: (a) Morphological analysis of hCdH on a bright field microscope showing hCdH clusted together in a colony. (b) Magnified view of (a). Scale bars: 100 μm. (c) Bright field images showing hCdH-chol forming some branch-like structures similar to the cholangiocyte characteristics. (d) Magnified view of (c). Scale bars: 100 μm.
|
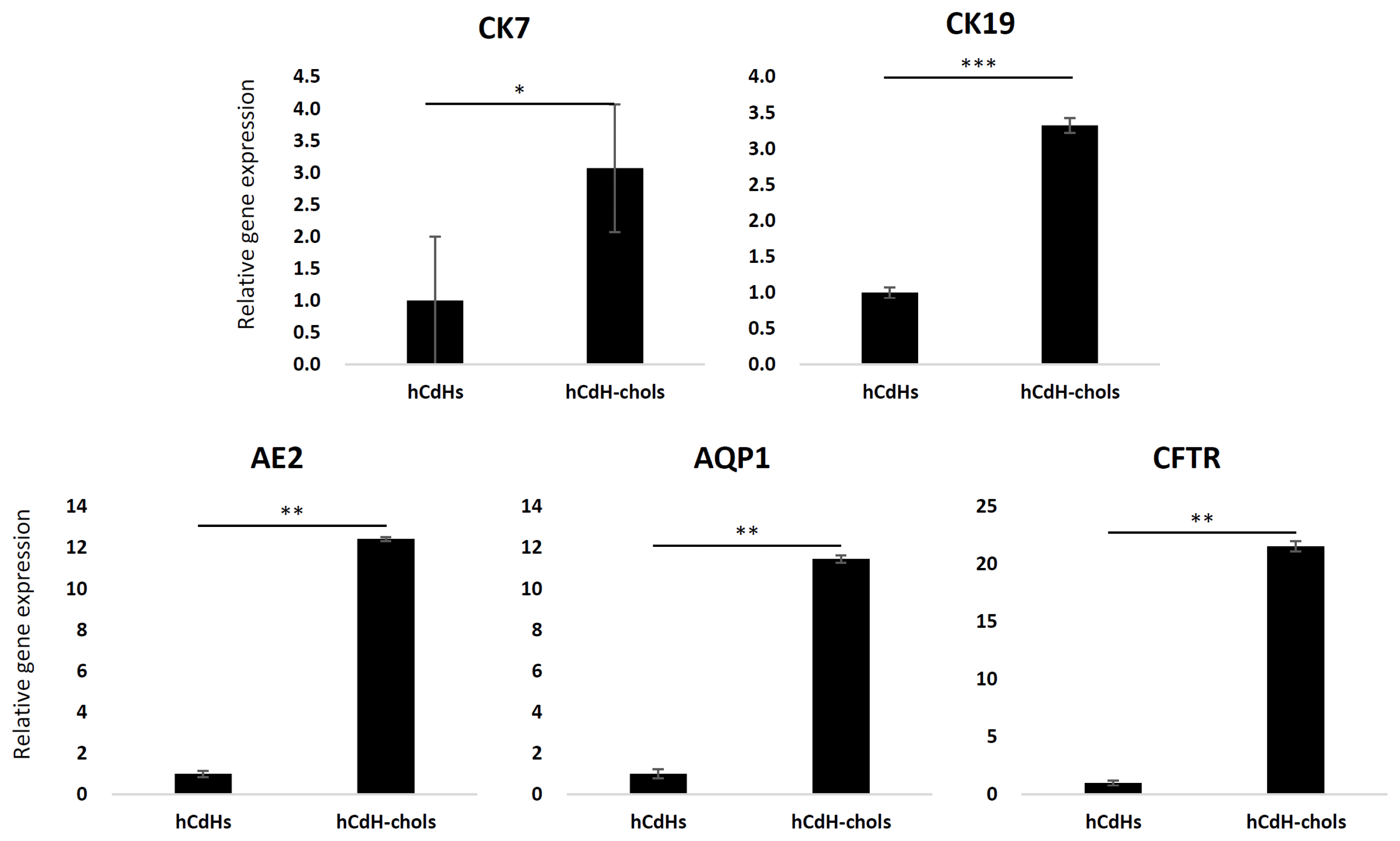 | Fig. 3Cholangiocyte markers of the relative expression between hCdH-chol and their negative control CdH as determined by RT-qpcr. GAPDH was used as the housekeeping gene. The data is shown as the mean value±SD. *p<.05, **p<.01, ***p<.001.
|
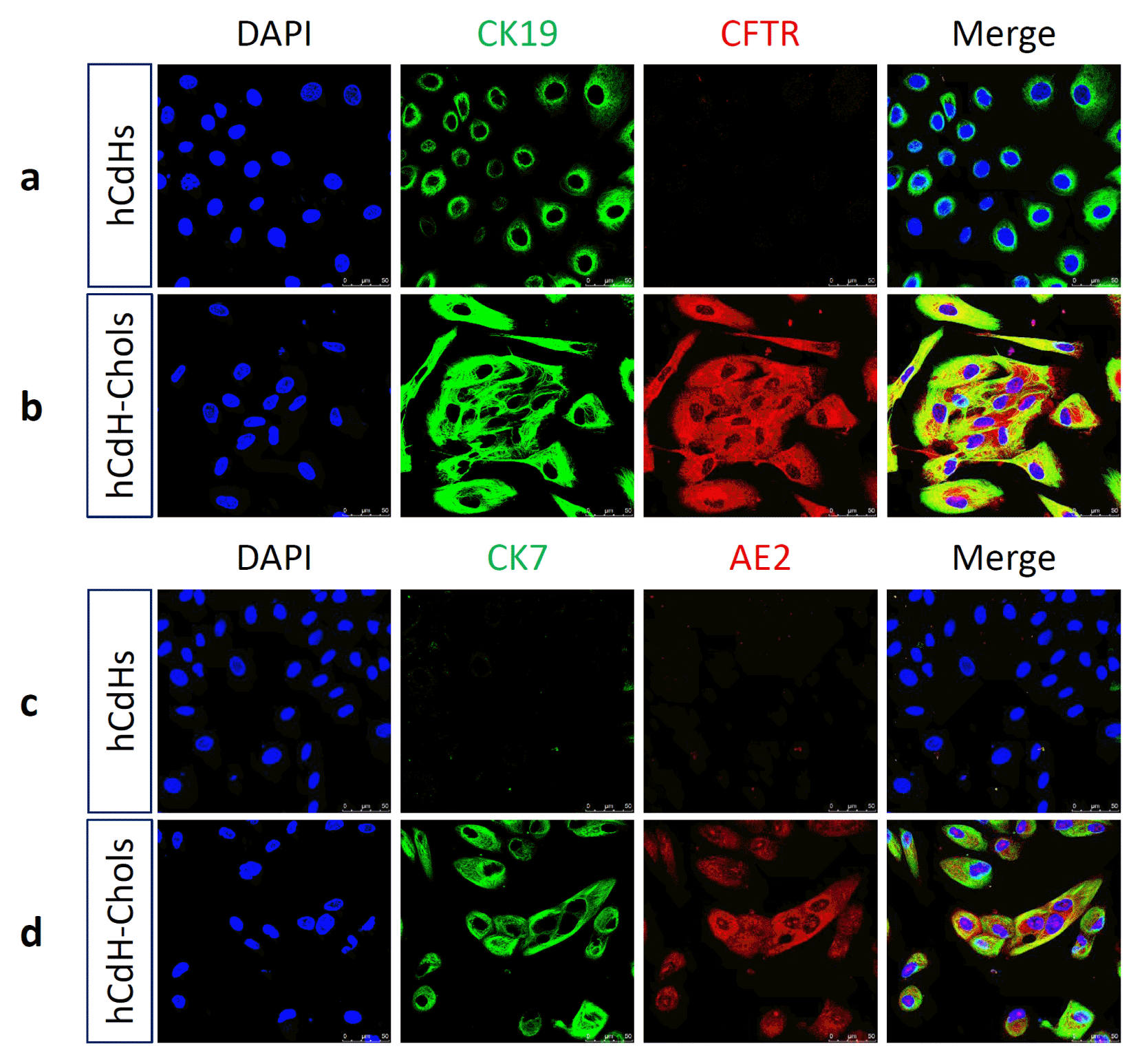 | Fig. 4hCdH-Chol expressed cholangiocyte markers seen through confocal microscopy. (a) Negative control hCdH stained with CK19 (green), CFTR (red). (b) CK19 (green), CFTR (red). (c) CK7 (green), AE2 (red). (d) CK7 (green), AE2 (red), and merged together showing a lack of expression of these genes. (d) hCdH-chol stained with CK7 (green) and AE2 (red) shows upregulation of these genes as seen through RT-qpcr. The nuclei were counterstained with Hoechst 33342 (blue). Scale bars, 50 μm.
|
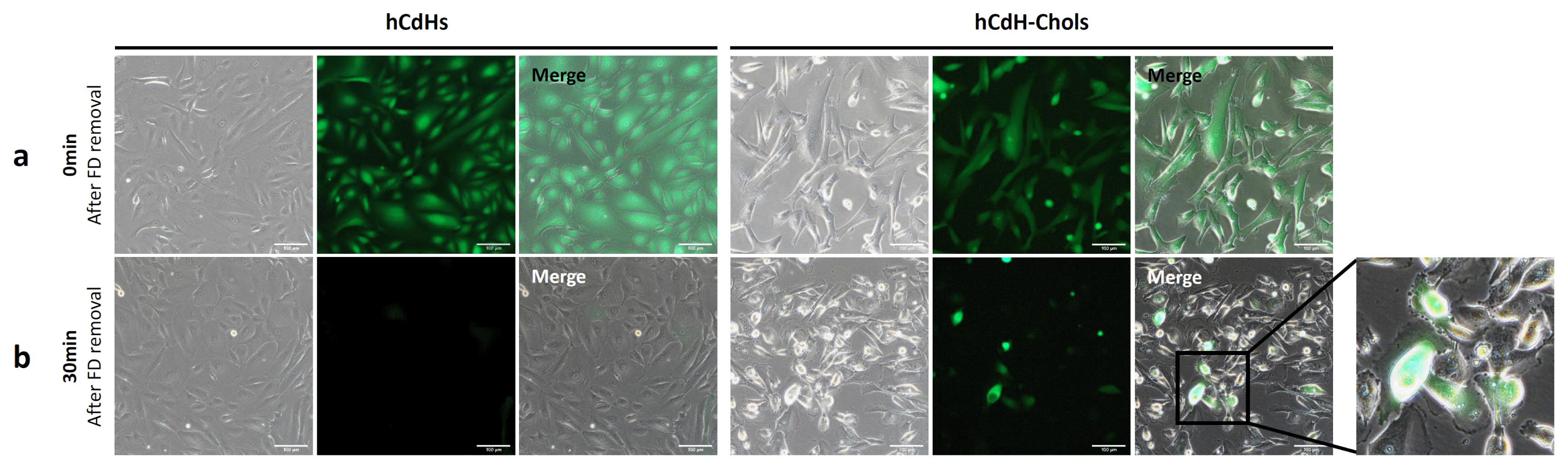
Table 1 summarizes the different chemicals used for cholangiocyte differentiation, and the materials used for artificial bile ducts can be found in Table 2, 3. Table 3 shows the full names of abbreviations used in the text.
Table 1
Differences in cholangiocyte protocols
| Author | Cell Source | Chemicals used | Functional assay used |
|---|---|---|---|
| Sampaziotis et al. (29) | Human induced pluripotent stem cells | 20 ng/ml EGF | Rhodamine 123 assay, cholyl-lysyl-fluorescein assay, ALP staining, GGT activity. |
| Dianat et al. (31) | Human pluripotent stem cells and HepaRG cells | 50 ng/ml GH, 25 ng/ml EGF, 10 ng/ml IL-6, 10 μM sodium taurocholate hydrate. | Acetylcholine, somatostatin and ATP stimulation and cholyl-lysyl-fluorescein assay |
| Ogawa et al. (32) | Human pluripotent stem cells co-cultured with OP9 stromal cells | 20 ng/ml HGF, 50 ng/ml EGF, 5 ng/ml TGFβ | Rhodamine 123 asay and Forskolin-induced swelling assay |
| De Assuncao et al. (33) | Human induced pluripotent stem cells | 100 μg/ml TGFβ | Calcium stimulation |
| Kido et al. (34) | CPM+progenitor cells | 40 ng/ml R-spondin 1, 40 ng/ml WNT3a | None |
![]()
Table 2
Differences in scaffold biomaterials
| Authors | Polymer used | Cells used |
|---|---|---|
| Aikawa et al. (35) | Polycaprolactone and polyglycolic acid | None |
| Perez Alonso et al. (36) | Collagen tubes coated with 2% agarose hydrogel | None |
| Miyazawa et al. (37) | Polycaprolactone and polylactic acid reinforced with polylactic fibers | Bone marrow cells |
| Park et al. (38) | Polyvinyl alcohol and polycaprolactone | None |
| Sampaziotis et al. (30) | Collagen | Extrahepatic cholangiocyte organoids |
![]()
Table 3
Abbreviations and full names
![]()
Go to :
Conclusions
Cholangiopathies remain problematic due to the lack of therapeutic solutions found on the market. Liver transplantation being the only solution is a disadvantage as rejection, adverse immune responses, and lack of donors limit its use. At the present time, only one drug can be found on the market to treat cholangiopathies. However, this has proven to be inadequate as a treatment as the complexity involved in the mechanisms of these diseases is an obstacle.
While much research has been done in term of cholangiocyte differentiation as well as the use of three-dimensional bioprinting to produce an artificial bile duct, there are still significant problems to overcome. The problematic use of hPSCs and induced pluripotent stem cells lies in the use of retroviral integration, teratoma formation, low production efficiency, and time consumption.
Therefore, much research needs to be done to improve the quality of life and the mortality rate of patients suffering from cholangiopathies. Recent findings and research will open the door to more hopeful work to find a cure through regenerative medicine. The future looks bright as the advances in biliary tree medicine keep ameliorating from past decades up until the present.
Go to :
 XML Download
XML Download