

 PDF
PDF Citation
Citation Print
Print


Introduction
Chronic kidney injury can progress end-stage renal failure that causes an irreversible glomerular and tubular damage lead to loss of renal function (1). Apoptosis, oxidative damage and microvascular rarefaction are responsible for glomerular and tubulointerstitial fibrosis in chronic kidney failure (2).
Aristolochic acid (AA) is obtained from the Chinese herb Aristolochic fangchi (3). In recent studies showed that AA leads to renal damage has been identified (4). AA effects on proximal tubular cells and causes Chronic Kidney Diseases (CKD) and AA nephropathy (AAN) create progress fibrosis rapidly (5). Therefore, renal functions deteriorate and end stage renal failure occurs in a short while (6). Several studies asserted that specific AA-DNA adducts formation in renal epithelial cells after AA injection and these adducts effect proximal tubular epithelial cells (PTEC) regeneration negatively leading to apoptosis. Therefore, irreversible proximal tubular atrophy occurs (7–11). CKD is caused by the development of renal fibrosis. It is characterized by renal interstitial fibrosis, tubular atrophy, interstitial inflammatory cell infiltration, and interstitial matrix accumulation. In fibrotic kidneys, type 1 and type 3 together with basal membrane type 4 collagen are the most common types of collagen. Kidney fibrosis is characterized by collagen synthesis accumulation, cross-linking and decreased degradation (12, 13). AAN patients are observed high serum creatinine rate with anemia and proteinuria. AA induces loss of peritubular capillary that results of hypoxia and tubular cell death (14, 15). Transplantation of mesenchymal stem cells (MSCs) was performed to provide renal repairment of damaged kidney in most of studies. Therefore, stem cells were considered therapeutic tool for the treatment of kidney diseases (16–20).
Placenta is used as a source of MSCs alternatively and placenta is an important reserve for stem cells and progenitor cells. Chorionic villus, amniotic membrane, umbilical cord stroma and amniotic fluid are indicated as a source of MSCs in placenta (21, 22). Diaz-Prado and colleagues determined that isolation, localization, phenotypic characterization and differentiation potential of human amniotic mesenchymal stem cells (hAMSC) (23). AMSC show plastic adherence and fibroblast-like growth as MSCs obtained from bone marrow. They express specific cell surface markers such as CD90, CD44, CD73, CD105, CD166 and CD29 and lack expression CD45, CD34, CD14, HLA-DR (23, 24).
Several studies demonstrated the effect of MSCs that obtain from bone marrow on CKD (25–28). However, effects of placental-derived mesenchymal stem cell in CKD experiments are limited. In the present study, we aimed to investigate whether amnion membrane derived mesenchymal stem cells can repair fibrosis that occurs because of chronic kidney failure and they are effective in mechanisms of proliferation and apoptosis.
Go to : 
Materials and Methods
Isolation of human amnion derived mesenchymal stem cells
Human term placentas of normal pregnancies (range 38~42 weeks, n=6) were obtained after spontaneous delivery or caesarean section with informed consent. Approval of the Ethical Committee of the Medical University of Akdeniz was granted. Isolation of hAMSC was performed according to the protocol of Soncini et al 2007 using enzymatic treatment of the amnion with collagenase A and DNase (both from Roche, Penzberg, Germany) after manual separation from the chorion (29). hAMSC were cultured in DMEM (Lonza, Basel, Switzerland) low glucose supplemented with 15% FBS (FBS Gold, both from Gibco, Invitrogen, Paisley, UK).
Immunophenotyping of cells
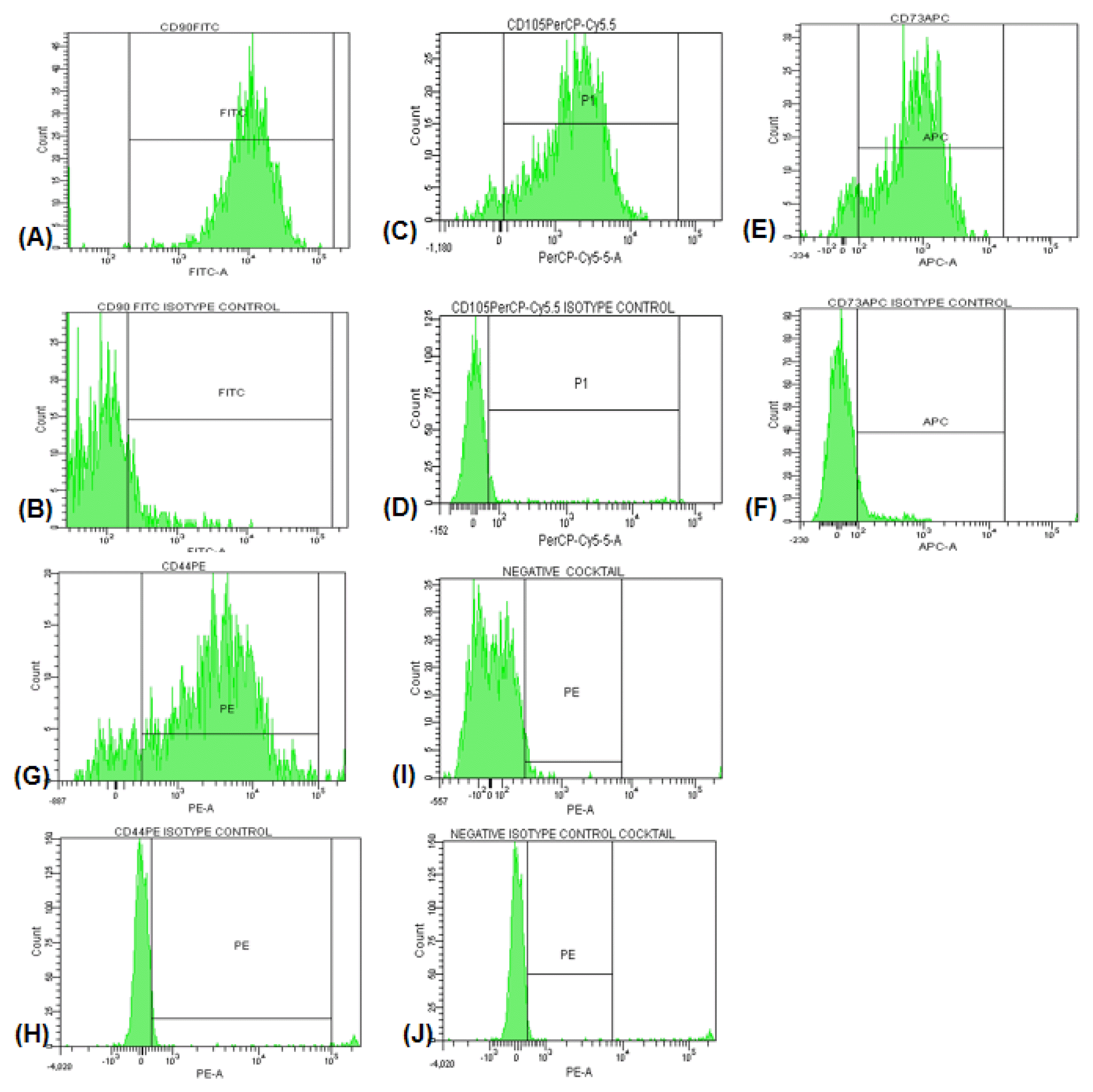
The cell surface marker phenotype of these hAMSCs was analyzed by flow cytometry and shown to be for CD90+, CD105+, CD73+, CD44+ and negative cocktail containing CD34, CD11b, CD19, CD45 and HLA-DR. hAMSCs were analyzed with a FACS Aria III Cell Sorter flow cytometry and the CellQuest software (BD Biosciences, New Jersey, USA).
Chondrogenic, osteogenic and adipogenic differentiation
For induction of osteogenic, chondrogenic, or adipogenic differentiation, hAMSCs were cultured in StemPro Osteogenic, StemPro Chondrogenic, or StemPro Adipogenic differentiation media(Life Tech, Carlsbad, USA), respectively, and with appropriate supplements. At week 3 of postosteogenic, postchondrogenic and postadipogenic inductions cells were washed with phosphate buffered saline (PBS) and fixed in 4% paraformaldehyde for 10 min. The cells were stained with Alizarin Red, Alcian Blue, and Oil Red O dyes (Sigma–Aldrich, St Louis, USA) for detection of calcium deposits, proteoglycans, and fat vacuoles as an indication of osteogenic, chondrogenic, and adipogenic differentiations, respectively.
Animal experiment
The study involving both human (File No: 294) and animals (File No: 2014.07.03) was conducted in accordance with the principles of the Helsinki Declaration and was approved by the ethical committee of Akdeniz University.
Aristolochic acid I (AA; Sigma-Aldrich, St Louis, USA) was used to mimic the structural and functional damage of CKD. AA was dissolved in Dimethyl sulfoxide (DMSO, Sigma-Aldrich, St Louis, USA). Animals were randomized in 6 groups. (i) Control group (n=5), Sham groups that received intraperitoneal injection of DMSO as a vehicle for 6 weeks. (ii) After 6 weeks one of the Sham groups was waited for 30 days (S+30; n=5) and (iii) the other Sham group was waited for 60 days (S+60; n=6) and sacrificed (iv). Every three days, rats received intraperitoneal injections of 10 mg/kg body weight AA for six weeks to induce AA group (n=6). After AA was taken, we waited six weeks thus we made a model of CKD. Then AA group was sacrificed. In hAMSCs groups, after rats were received AA for six weeks, 6×105 hAMSCs were injected from in the tail vein with 500 μl DMEM medium. (v) One of hAMSCs group was waited for 30 days (AA+hAMSCs+ 30; n=5) and (vi) the other MSCs group was waited 60 days (AA+hAMSCs+60; n=6) and sacrificed. Furthermore, all cell treated rats received daily subcutaneous injections of cyclosporine A (1 mg/day, Cell Signaling, Danvers, USA), starting one day before engraftment and continuing for seven days after engraftment.
Functional and histological damage assessment
Functional damage was evaluated by serum creatinine and urea levels. Blood samples were collected prior to sacrifice. Serum creatinine was analyzed by creatinine assay kit (Invitrogen, Paisley, UK) according to manufacturer’s protocol. BUN was measured by serum urea with urea assay kit according to protocol of manufacturer (Invitrogen, Paisley, UK). Tissue damage was assessed through morphological analysis using Masson’s Trichrome and Sirius Red (Sigma-Aldrich, St Louis, USA) staining. Percentage of fibrosis was measured by Image J (Maryland, USA).
Immunohistochemistry and TUNEL analysis
The immunohistochemical procedure has been described elsewhere (30). Briefly, slides were incubated with primer antibodies that are mouse monoclonal PCNA (1:1000, Cell Signaling), rabbit monoclonal Ki67 (1:100 dilution, Abcam) and mouse monoclonal human anti-Mitochondrial Antibody (1:250 dilution, Abcam), IL-6 (1:100 dilution, Abcam) and Collagen type I (1:100 dilution, Novus), through overnight at 4°C. Staining was completed by performing LSAB 2 System-HRP (Dako) and then AEC system was used for developing. Hematoxylin counterstaining was performed. Apoptosis was determined by terminal transferase-mediated deoxyuridine triphosphate nick end-labeling (TUNEL) assay (In situ cell detection kit-POD, Roche, Risch-Rotkreuz, Switzerland) in paraffin-embedded tissue sections according to protocol of the manufacturer.
Western blotting
Immunoblot analysis were performed as described previously (31). Briefly, Membranes were incubated with PCNA (1:2000 dilution, Cell Signaling, Danvers, USA), p57 (1:250 dilution, Santa Cruz, Dallas, USA), PARP-1 (1:500 dilution, Abcam, Cambridge, UK) and Beta Actin (1:5000 dilution, Abcam, Cambridge, UK) at 4°C through overnight. Then the membranes were incubated with horseradish peroxidase ppconjugated IgG (Bio-Rad Laboratories, Inc., Hercules, USA) for 2 h at room temperature. SuperSignal CL-HRP Substrate System (Thermo Fisher Scientific, Waltham, USA) was used for immunolabeling. Membranes were exposed to Hyperfilm (Amersham, Little Chalfont, UK) and analyzed by using Alpha Digi Doc 1000 gel documentation unit (Alpha Innotech Corporation, San Leandro, USA).
Statistical analyses
Data were expressed as mean±SEM. One way ANOVA test was used to compare means of multiple groups using GraphPad Prism (version 4.0; GraphPad Software, La Jolla, USA). The level of statistical significance was set p<0.05.
Go to :
Results
Characterization of hAMSCs
Human placenta amnion membrane-derived MSCs that showed fibroblast-like phenotype were isolated. At passage 3, hAMSCs were characterized with cell surface markers by flow cytometry analyses: CD90, CD105, CD73, CD44 positive and negative cocktail containing CD34, CD11b, CD19, CD45 and HLA-DR with isotype controls. The percentages of cell surface markers were 98.8%, 93.5%, 95.3%, 88.6% and 3.9% respectively (Fig. 1A–J).
Phase contrast microscopic images of human amniotic membrane MSCs in the second passage are given in Fig. 2A and 2B. hMSCs were spindle or triangular shaped and adhered to plastic.
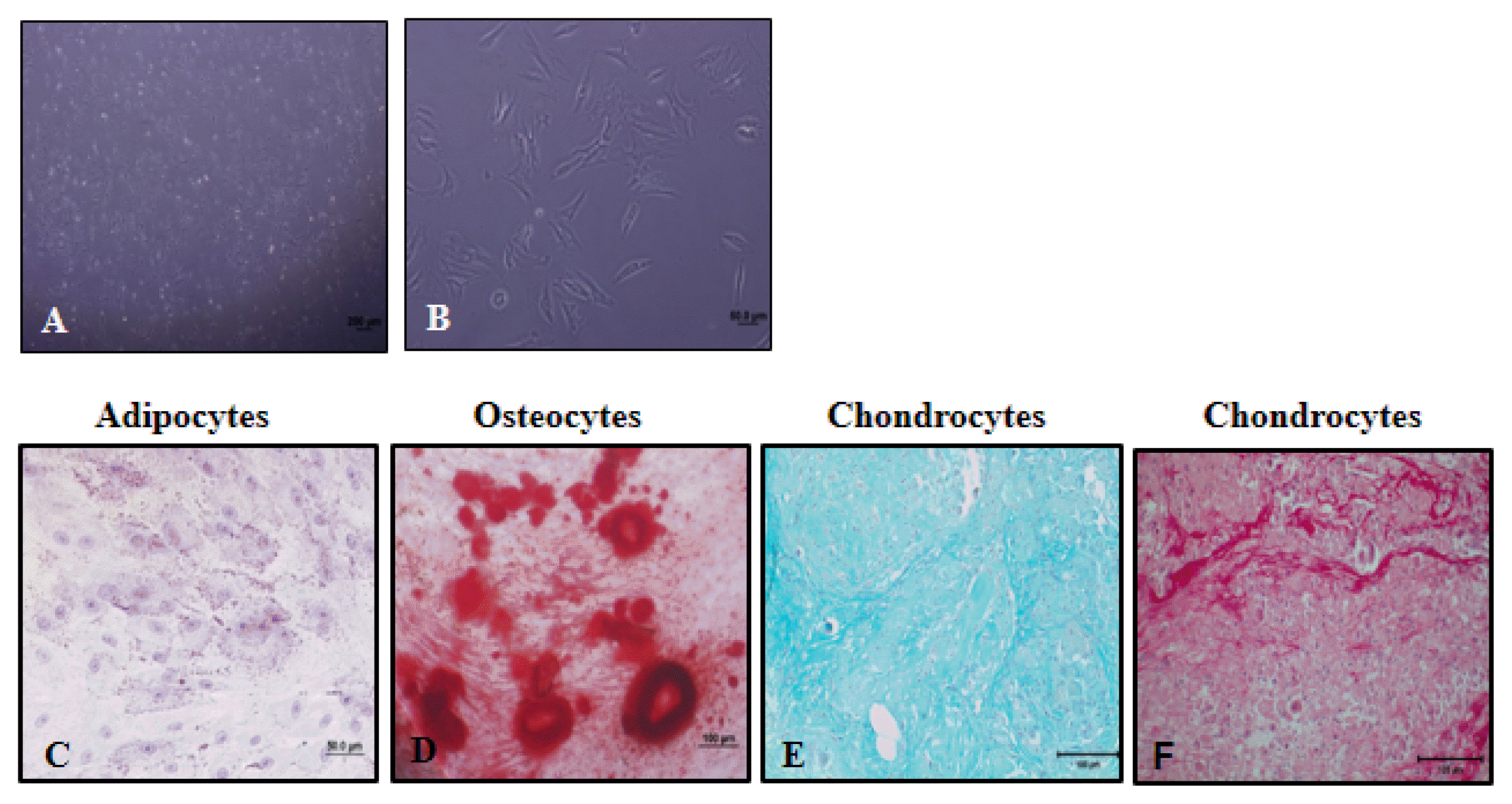 | Fig. 2Phase contrast microscopic images of human amniotic membrane derived mesenchymal stromal cells in 4X (A) and 40X magnifications (B). hAMSCs were differentiated into adipocytes (C., as shown by Oil Red O staining), osteocytes (D., as shown by Alizarin Red S staining) and chondrocytes (E and F, as shown by Alcian Blue and Sirius Red staining).
|
hAMSCs undergo adipogenic, chondrogenic and osteogenic differentiation
hAMSCs could differentiate into adipocytes, chondrocytes and osteocytes. To show lipid droplets, cells were stained with Oil Red O in adipocyte differentiation (Fig. 2C). To determine deposition of calcium, cells were stained with Alizarin Red S in osteocyte differentiation (Fig. 2D). To determine chondrocytes, cells were stained with Alcian Blue (Fig. 2E) and Sirius Red (Fig. 2F). While proteoglycans were stained with Alcian blue, collagens were stained with Sirius Red.
Morphological studies
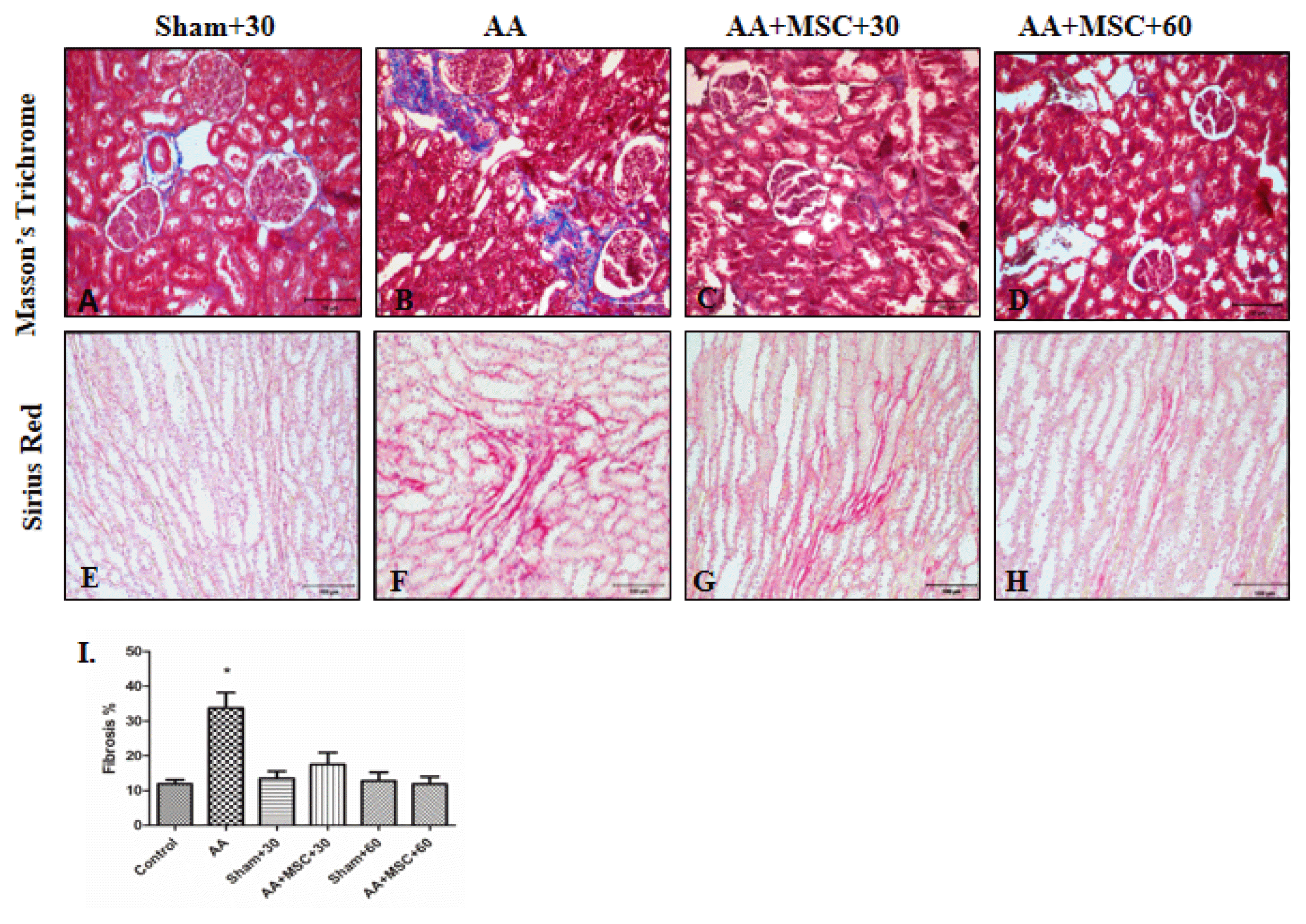
To evaluate renal histology and determine renal fibrosis, paraffin-fixed specimens were sectioned at a thickness of 5 μm and stained with Masson’s Trichrome (Fig. 3A–D) and Sirius Red (Fig. 3E–H). Increased extracellular matrix production was shown with Masson’s Trichrome. Accumulation of collagen was shown with Sirius Red. There were no significant extracellular matrix and collagen in control and sham groups (S+30, S+60) (Fig. 3A, E). Morphological differences were not found between control and sham groups (control and S+60 results not shown). In AA group, significant increase of extracellular matrix was observed near proximal and distal tubules compared to hAMSCs groups (AA+MSCs+30 and AA+MSCs+60) (Fig. 3B). Also in AA group, accumulation of collagen was more than MSCs groups at the same area (Fig. 3F). Although rats were treated with hAMSCs, Masson’s Trichrome and Sirius Red staining showed fibrosis reduction in both AA+hAMSCs groups (Fig. 3C and 3D) and (Fig. 3G and 3H). The percentage of fibrosis measured with Image J.
MSC tracking
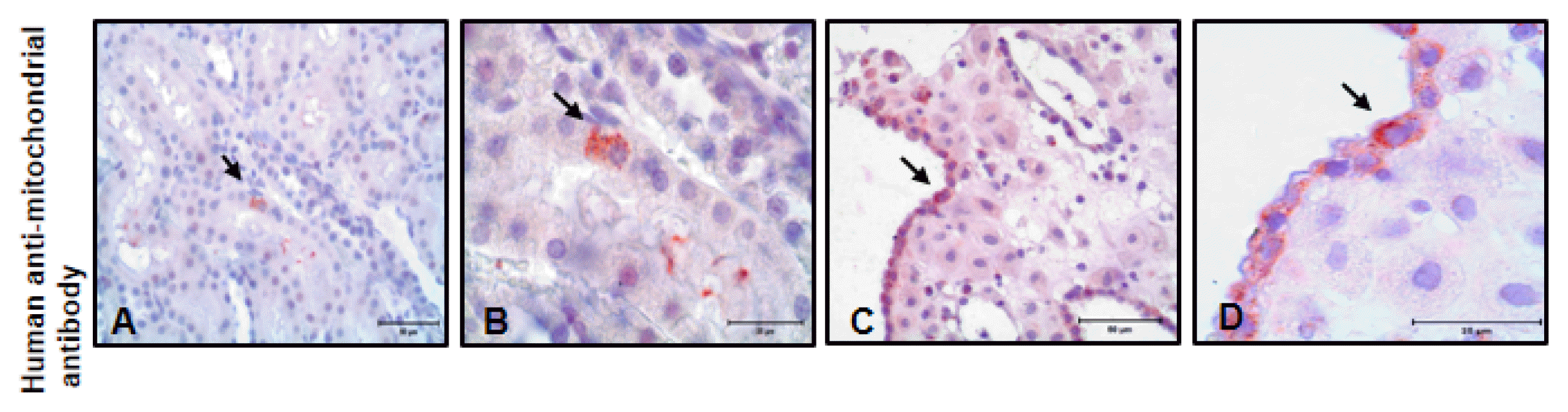
To determine hAMSC localization in kidney tissues, immunohistochemistry analysis was performed. We used human anti-mitochondrial antibody. hAMSC was observed in kidney tissues (Fig. 4A and 4B). Furthermore, human decidua tissue was used as a positive control for human anti-mitochondrial immunostaining (Fig. 4C and 4D).
Proinflammatory cytokine IL-6 and Fibrosis marker collagen type I immunostaining
Expressions of the proinflammatory cytokine IL-6 significantly increased in AA group especially in proximal and distal tubule cells. However, after hAMSCs injection expressions decreased both AA+hAMSC+30 and AA+ hAMSC+60 groups significantly (Fig. 5A–D). Renal interstitial fibrosis was determined a significant increase in the collagen I protein in AA group compared to control group, sham groups, AA+hAMSC+30 and AA+hAMSC+60 groups. In hAMSCs groups expressions of collagen I decreased significantly (Fig. 5E–H).
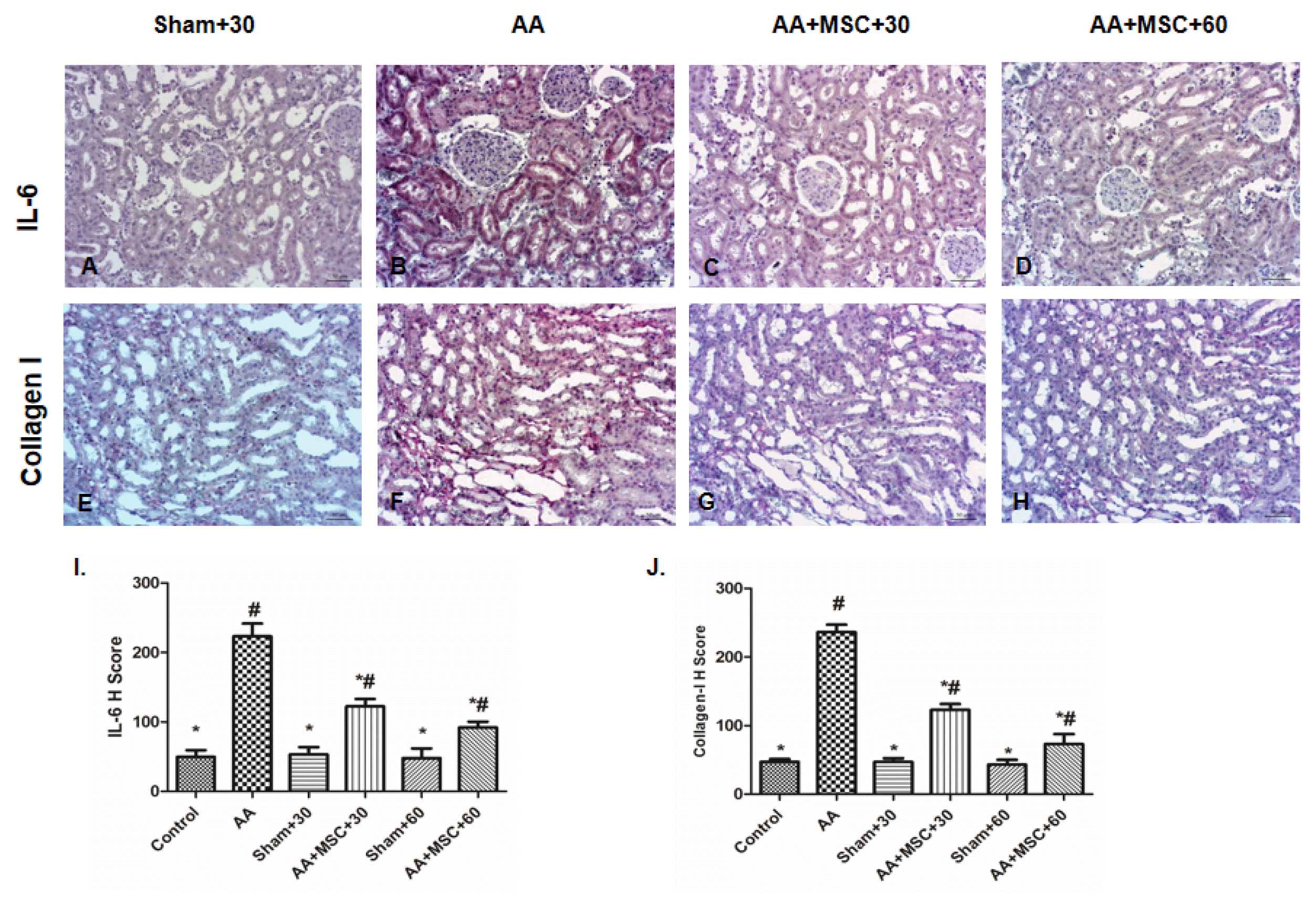 | Fig. 5Immunohistochemical staining of kidney tissue sections for the sham+30, AA, AA+hAMSC+30 and AA+hAMSC+60 groups using IL-6 (A–D), Collagen-I (E–H) I and J, H scores of IL-6 and Collagen-I. Values presented as mean ± SD. AA group compared with Sham, Control, AA+hAMSC+30, and AA+hAMSC +60 groups. Values statistically significant at: *p<0.05, Sham and Control groups compared with AA and hAMSC groups; #p<0.05.
|
Proliferation (PCNA and Ki67 staining) and western blot analysis of PCNA expression
The proliferations markers PCNA and Ki67 were expressed in sham group Kidney Tissue (Fig. 6A and 6E). The immunohistochemistry staining of PCNA and Ki67 in AA group kidneys were observed weak signal (Fig. 6B and 6F). However, after injection of hAMSCs, in both AA +hAMSCs groups cell proliferation was significantly increased. Increase of PCNA and Ki67 staining were observed especially in proximal and distal tubule cells. PCNA and Ki67 staining were not observed significant differences between AA+hAMSC+30 and AA+hAMSC +60 groups (Fig. 6C and 6D, and Fig. 6G and 6H). Western blot analysis results showed that PCNA expression was decreased in the AA group compared with control, S+30 and S+60 groups. Furthermore, after hAMSCs injection, expression of PCNA was increased significantly in the AA+hAMSCs groups (Fig. 7).
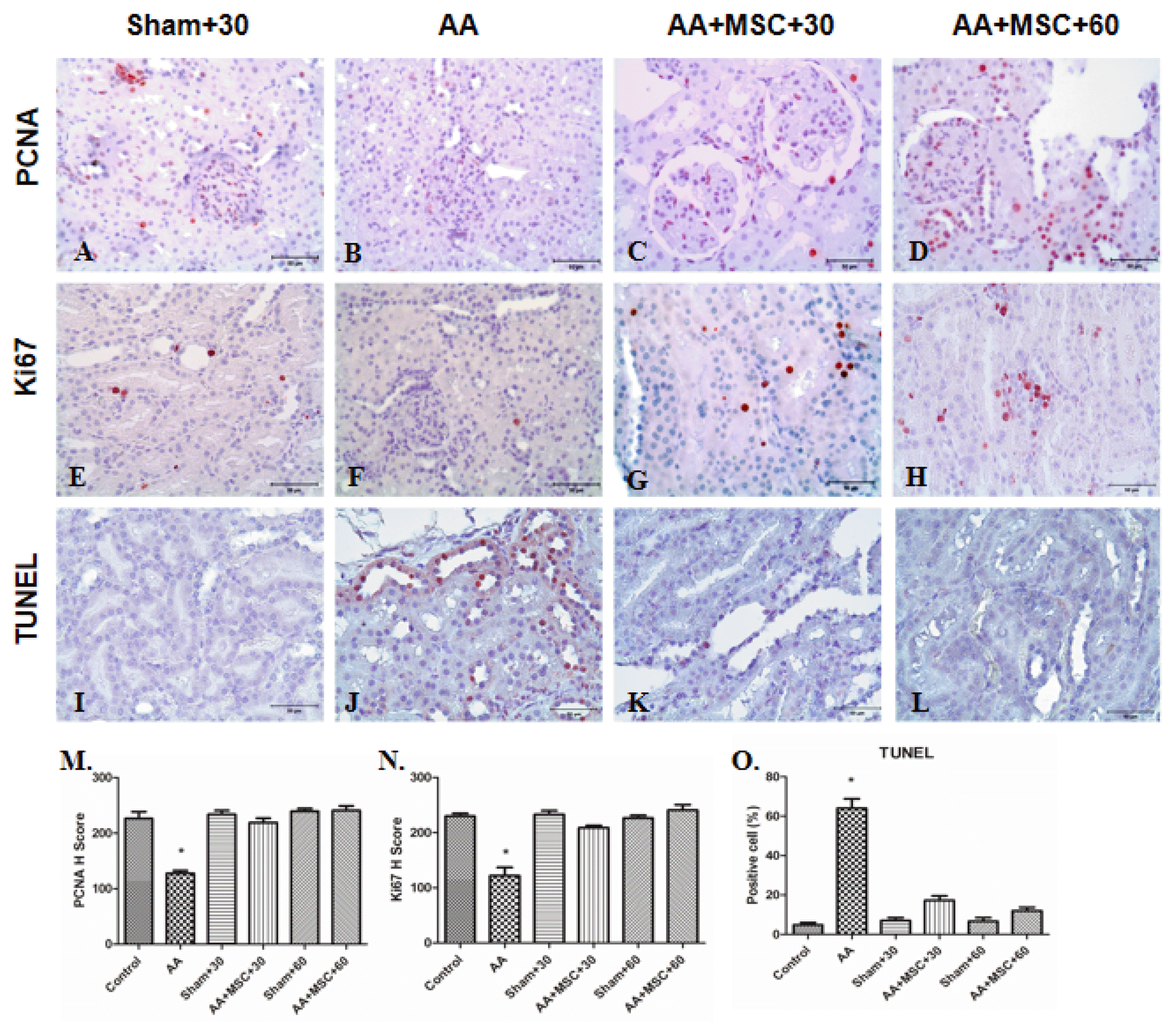 | Fig. 6PCNA, Ki67 and TUNEL immunostaining in kidneys tissue. For PCNA, sham+30 (A), AA (B), AA+ hAMSC+30 (C) and AA+hAMSC+ 60 (D) groups, for Ki67 sham+30 (E), AA (F), AA+hAMSC+30 (G) and AA+hAMSC+60 groups (H) and for TUNEL immunostaining sham+30 (I), AA (J), AA+hAMSC +30 (K) and AA+hAMSC+60 (L) groups. M and N, H scores of PCNA and Ki67. (O) Positive cells number for TUNEL. Values statistically significant at: *p<0.05.
|
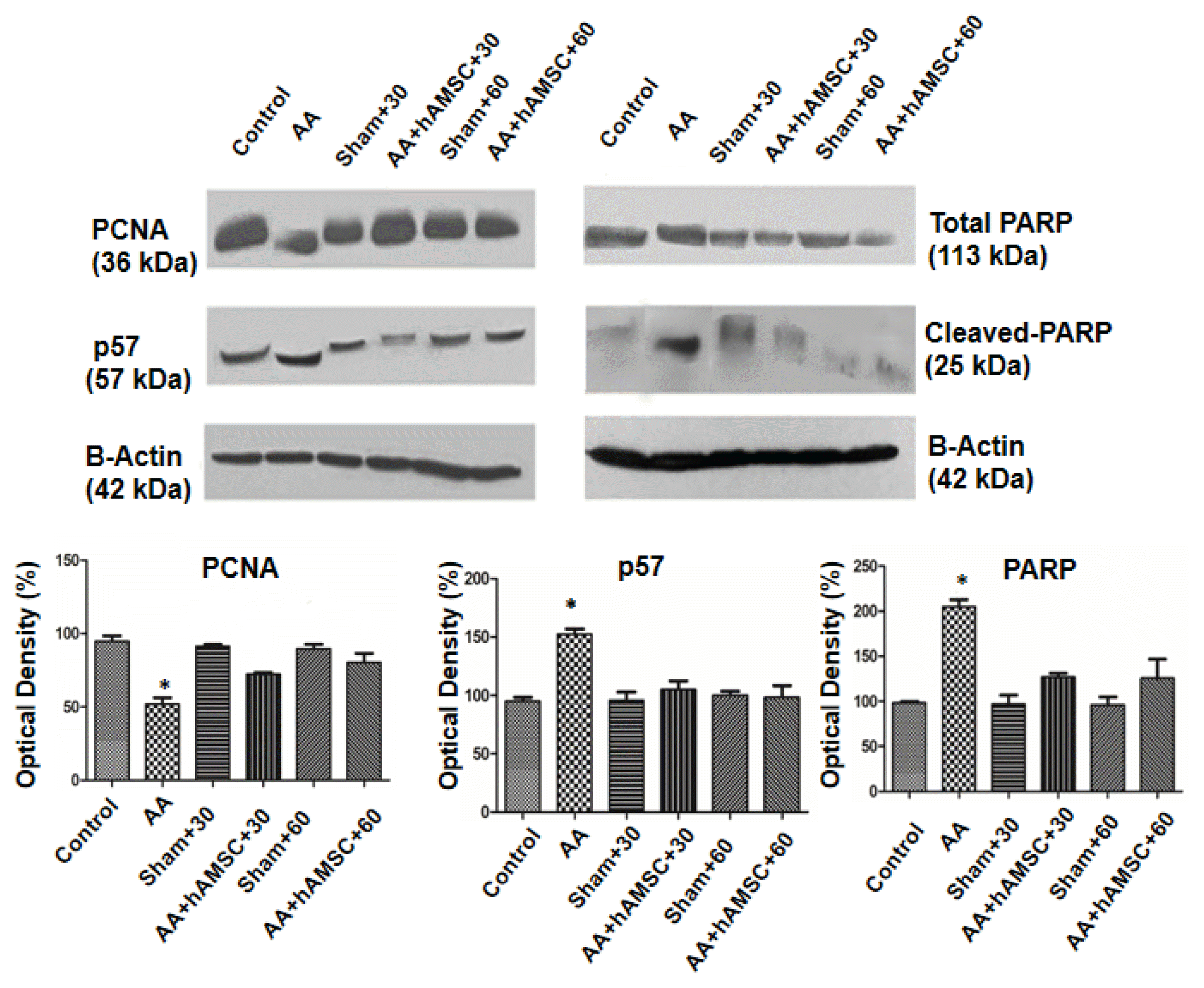 | Fig. 7Western blot results of PCNA, p57, total PARP and cleaved-PARP. AA administration caused increased PARP and p57 expressions but decreased PCNA compared with control, sham+30 and sham+60. After hAMSC administration PARP and p57 expressions decreased and PCNA expression increased significantly. Protein levels between AA+hAMSC+30 and AA+hAMSC +60 were not statistically significant. All results were normalized to beta actin. Graphics represent means of optical densitometry measurements. *p<0.005.
|
Apoptosis (TUNEL staining) and western blot analysis of PARP-1 expression
In AA group, numbers of apoptotic cells were observed to increase compared with sham groups (Fig. 6K and 6L). When rats treated with hAMSCs, apoptotic cells were reduced significantly in tubules (Fig. 6M and 6N). We determined expression of Parp-1 protein as an apoptosis marker. In western blot analysis, Parp-1 expression of AA group was compared with that of control and sham groups. In AA group, apoptosis level was higher than control and sham groups. However, after hAMSCs injection, Parp-1 expression level of both AA+hAMSCs groups was decreased significantly compared with AA group. Therefore, TUNEL results supported western blot analysis of Parp-1 expression (Fig. 7).
Western blot analysis of p57 expression
In AA group, p57 expression was increased compared with control and sham groups. However, p57 expression level was reduced compared to AA group after hAMSCs injection. Differences were not finding between both hAMSC groups and control and sham groups in expression of p57 significantly (Fig. 7).
Functional renal damage studies
Renal function was assessed by serum creatinine and BUN. Therefore, the development of CKD was evaluated in model of AAN. AA group rats showed significantly higher levels of serum creatinine and BUN than normal control and sham groups. When rats were treated with a single intravenous of 6×105 hAMSCs after AAN, serum creatinine and BUN levels were decreased significantly according to levels of AA group. Furthermore, creatinine and BUN levels of the both AA+hAMSCs groups were not significantly different from normal control and sham groups (Fig. 8).
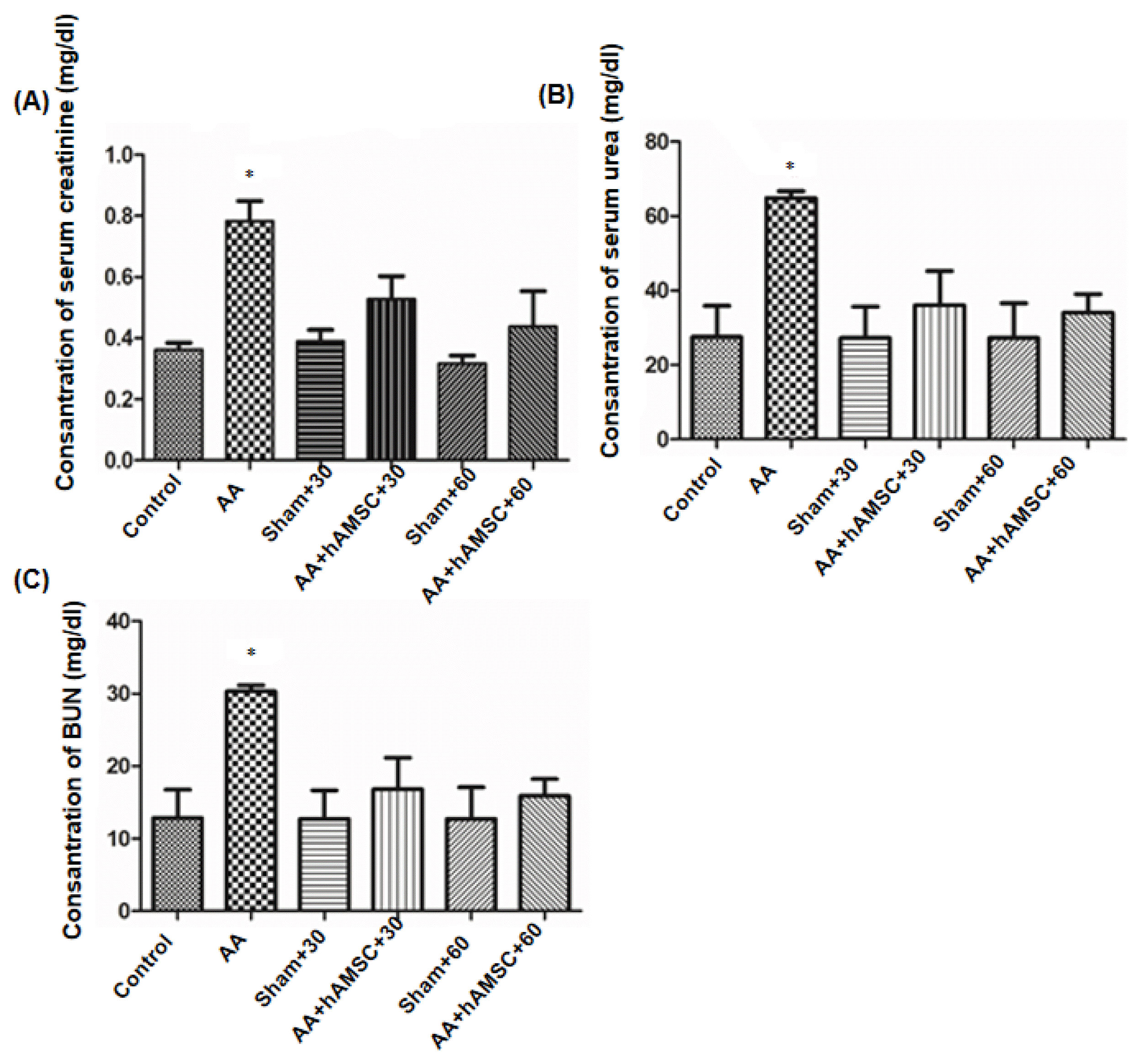 | Fig. 8Functional damage assessment. Serum creatinine (A), serum urea (B) and BUN (C) levels were determined. AA administration caused increased serum creatinine, serum urea and BUN compared to control and sham groups significantly. After hAMSC administration, serum creatinine, serum urea and BUN levels decreased significantly. Graphics represent means of optical densitometry measurements. *p< 0.005
|
Go to :
Discussion
CKD increases day after day as a public health problem. Recently, cell therapies are applying for the repair of kidney injury. More studies, stem cell transplantation was carried out to damage kidney for provision of renal repair (32–36). Human amniotic membrane is thought to be a reservoir of mesenchymal stem cell. For these reason, hAMSCs has been the focus of interest as a source of MSCs in cell transplantation and regenerative medicine. In this study, we showed that a single administration of amniotic membrane derived MSCs prevents CKD after induction of AAN in rats.
Several studies showed that the administration of MSCs protects rats against CKD in different models. To ensure recovery after application of MSCs could be explained by several mechanisms. MSCs can differentiate into kidney cells. One of the mechanisms is paracrine effect. In model of chronic allograft nephropathy, MSCs migrated to the injured tissue and integrated to kidney. Briefly, MSCs has been determined to ameliorate the process of fibrosis when MSCs applied in the CKD model (25, 35, 37, 38).
In present study, CKD model was constituted after 6 weeks AA injection, rats were waited six weeks. Therefore, AA induced CKD model was occurred after total 12 weeks. We determined fibrosis with Masson’s Trichrome and Sirius Red staining. In addition expressions of collagen-I was determined with immunohistochemistry staining. In AA group, fibrosis increased significantly compared with control and sham groups. Particularly, accumulation of collagen occurred in the vicinity glomeruli, proximal and distal tubules. Recent studies showed that the primarily target of AA is proximal tubules. According to Huang et al, AA applied kidney is showed to increase collagen in interstitial area (39). After hAMSC administration, accumulation of collagen decreased compared with AA group. Therefore, tubular damage was considered to ameliorate after hAMSCs injection.
In CKD, damaged tubular cells, macrophages, accumulating fibroblasts and infiltrating lymphocytes produce cytokines and growth factors, causing the kidneys to become inflammatory. This inflammation leads to interstitial fibrosis (40). Therefore, the levels of expression of the proinflammatory cytokine IL-6 was determined by immunohistochemistry. In AA group, expressions of IL-6 significantly increased. However after hAMSCs injection reduced expressions of IL-6 both AA+hAMSC+30 and AA +hAMSC+60 groups.
Renal functions are damaged rapidly in AAN and end-stage renal failure arises in a few months. Both in vitro and in vivo studies, specific AA-DNA adduction formation was demonstrated in renal epithelial cells after AA injection and this formation can prevent proximal tubular epithelial cells (PTEC) proliferation capacity. Besides, apoptosis was determined in CKD and this condition was suggested to be mechanism of tubular epithelial cell deletion. Progressing tubular atrophy correlate with unrecoverable regeneration and apoptosis of PTEC. Additionally, staining of Ki67 and PCNA inform about DNA damage. The reduction of PCNA expression in PTECs, AA-DNA addition formation is considered impairment of DNA repair and this situation induces defective cell proliferation and tubular atrophy (11). In present study, AA injected group was observed to decrease PCNA and Ki67 immunohistochemistry staining compared with control and sham groups. The reason for this is due to degradation of DNA repair mechanism, AA-DNA addition formation occurred. This condition indicated that reduction of proliferation in AA injected group. Furthermore, cell death was determined with TUNEL and western blot analysis of PARP-1 protein expression. In accordance with other studies (36, 41) both TUNEL and western blot analysis indicated that apoptosis increased in AA group compared to other groups. Li et al. reported that cell cycle arrest of PTECs was occurred after AA injection (42). We observed that expression of p57 increased in AA group compared with other groups. Therefore, unrecoverable PTECs can lead to apoptosis.
Sun et al. evaluated the use of human amniotic fluid stem cells (hAFSCs) in cell-based therapy (36). They investigated whether hAFSCs effect on interstitial fibrosis in unilateral urethral obstruction which is one of the CKD model. Therefore, proliferation and apoptosis mechanisms of tubular cells were investigated to evaluate therapeutic effect of hAFSCs injection. The mechanism that covers proliferation and apoptosis of tubular epithelial cell includes paracrine effect. Furthermore, proliferation of resident epithelial cells was suggested that a mechanism of basic-repair in ischemia induced tubular injury model (36).
Gatti et al investigated that MSCs derived microvesicles administration can ameliorate ischemia-reperfusion induced acute kidney disease (AKD) and CKD (41). After injection, microvesicles accumulated in glomeruli and damaged tubules temporarily and they induced proliferation of tubular cell. Furthermore, microvesicles reduced significantly tubular cell apoptosis. These biological effects are specific for MSC derived microvesicles because these effects were not seen in fibroblast derived microvesicles (41).
In this study, fibrosis can be considered to ameliorate due to increasing of renal tissue cell proliferation after hAMSCs administration. This study suggested that hAMSCs have an important role in tubular atrophy and hAMSCs can prevent damage formation in PTECs and other cells. This situation shows that hAMSCs have therapeutic features. In addition to, the increase of proliferation is considered to cause hAMSCs can replace with damaged cell and differentiate into adult tissue cells. Furthermore, hAMSCs have paracrine effects thus they can secrete various growth factors and cytokines.
We observed expression of apoptosis marker and cell cycle inhibitor decreased significantly after hAMSCs administration. Therefore, apoptosis can be prevented due to tubular cell regeneration.
Recent studies were reported that serum creatinine, BUN, urea levels decreased significantly after MSCs injection in various CKD models (26, 41, 43). In present study, when serum creatinine, serum urea and BUN levels were analyzed, we observed to increase in AA group compared with control and sham groups. After hAMSCs administration, levels of serum creatinine, serum urea and BUN decreased significantly. In AA-induced CKD model after hAMSCs injection, reduction of serum creatinine, urea and BUN levels indicate that hAMSCs have renoprotective effects.
Go to :
 XML Download
XML Download