

 PDF
PDF Citation
Citation Print
Print


Introduction
Hypoxia and stem cells
Stem cells exposed to hypoxia must physiologically adapt to low oxygen conditions to maintain their pluripotency and survival. Metabolic switching during hypoxia is essential in decreasing the accumulation of oxidative metabolism-derived reactive oxygen species (ROS) and the availability of oxygen (1–3). Recent studies indicate that differences in metabolic profiles of pluripotent stem cells are closely related to self-renewal and initial cell fate decision of stem cells. Gu and colleagues reported that naive pluripotent stem cells in humans exhibit a high glycolytic flux and pentose phosphate pathway activity. However, human embryonic stem cells (hESCs) primed under feeder-free conditions exhibit a low glycolytic flux (4). Moreover, metabolic switching induced by hypoxia prevents the excessive generation of ROS and reduces the demand for ATP, both closely associated with maintaining the physiological functions and survival of stem cells (1–3). Despite the metabolic switching, chronic hypoxia-induced ROS accumulation leads to mitochondrial apoptosis in embryonic and mesenchymal stem cells (5, 6). Hypoxia-preconditioned mesenchymal stem cells (HP-MSCs) exhibit higher transplantation survival rates and therapeutic potential than normoxia-preconditioned mesenchymal stem cells (MSCs) (7, 8). Other study showed that HP-MSCs exhibit higher concentrations of fatty acids and synthase-mediated lipogenesis, which stimulates cell migration, proliferation, and survival (9, 10). Additionally, hypoxia induces the hexosamine biosynthesis pathway, thereby increasing O-linked-N-acetyl-glucosaminylation concentrations, which is critical for anti-apoptosis in mouse embryonic stem cells (mESCs) exposed to hypoxia (11).
Hypoxia-inducible factor 1 (HIF1)
Semenza et al. first reported that hypoxia-inducible factor 1 (HIF1) is a human erythropoietin gene inducer in ischemic liver and kidney tissues (12). Alpha (HIF1α) and beta (HIF1β) subunits of HIF1 are dimerized in the nucleus, which activates the heterodimeric transcription factor for the HIF1-target genes (13, 14). HIF1 subunits belong to the basic helix loop helix (bHLH) and PER-ARNT-SIM (PAS) domain protein family (Fig. 1). The bHLH and PAS domains within HIF are required for its dimerization and for binding it to DNA. After nuclear translocation, HIF1 binds to the E-box-like hypoxia-responsive element (HRE), which comprises the consensus sequence 5′-[A/G] CGTG-3′ (3). The HIF1α subunit has two transactivation domains (TADs): specifically, the N-terminal TAD (NTAD), comprising amino acids 531~575, and the C-terminal TAD (CTAD), comprising amino acids 786~826. Deletion of inhibitory domain (ID) sequences enhances the transcriptional activity of the HIF1-target genes (15). HIF1α also contains an oxygen-dependent degradation domain (ODDD), which includes prolyl hydroxylase (PHD)-targeted prolyl hydroxylation sites (P402 and P564) (15, 16). HIF1α is controlled by post-translational modifications, including ubiquitination mediated by PHD, which is followed by its stabilization. In contrast, CTAD comprises another hydroxylation site (N803) (17). The hydroxylation of CTAD, which is induced by factor-inhibiting HIF1 (FIH), inhibits the interaction between HIF1α and CREB-binding proteins (CBP/p300), which decrease gene transcriptional activity of HIF1 (18). However, HIF1β does not contain an ODDD, CTAD, or ID. Owing to structural differences between the two HIF1 subunits, HIF1α plays an important role in regulating gene transcription activity of HIF1. Therefore, a deeper understanding of HIF1α is required to comprehend the functional regulation of stem cells under hypoxia.
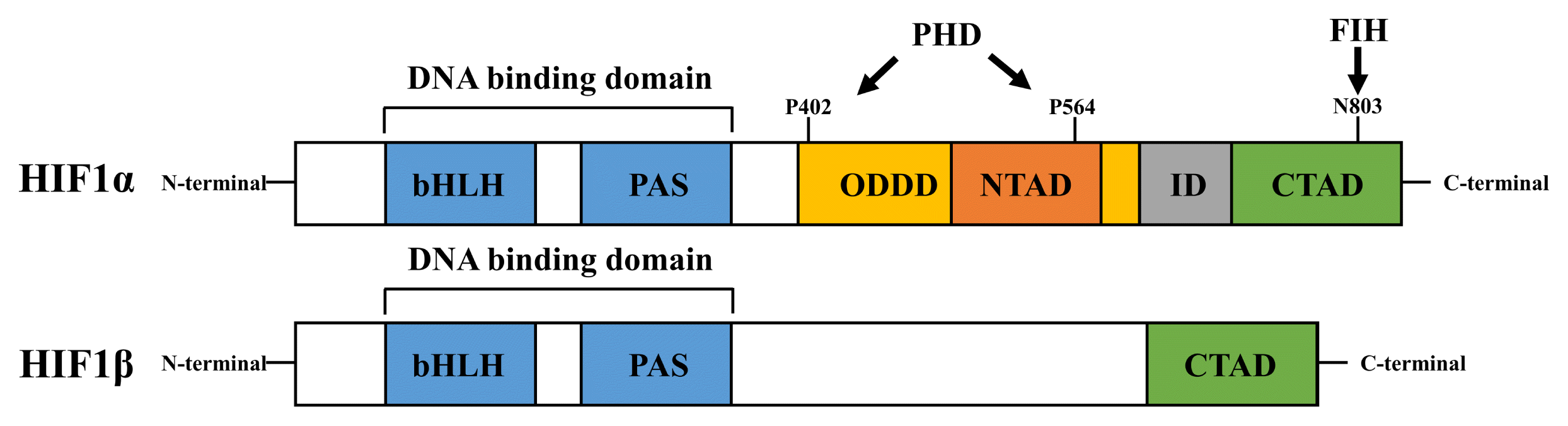 | Fig. 1Schematic structures of HIF1α and HIF1β domains. Both HIF1α and HIF1β possess bHLH and PAS domains for the formation of heterodimeric complexes and for DNA binding. HIF1α has two transactivation domains (NTAD and CTAD) and an inhibitory domain (ID), whereas HIF1β possesses only the CTAD domain. PHD hydroxylases possess two proline residues (P402 and P564) in the NTAD domain, whereas FIH hydroxylases possess an asparagine residue (N803) in the CTAD domain in HIF1α. These hydroxylated residues are ubiquitinated by VHL.
|
HIF1 stimulates the expression of various hypoxia-responsive genes, which in turn regulate various kinds of cellular physiology including metabolic reprogramming, anti-apoptosis, migration, proliferation, amyloid β production, and prion stabilization 3, 19–23). Recent studies have reported the role of HIF1α in stem cell physiology. Von Hippel–Lindau (VHL)-HIF1 signaling induces metabolic compartmentalization in embryonic cells, which is important for cardiac development and functional maturation (24). Sustained HIF1 activation induces cardiac chamber defects and dysfunction when conduction system regulating genes are altered (24). HIF1α induced by the GRP78-Akt axis is critical in enhancing functions, including the proliferation and survival of MSCs under hypoxia (25, 26). In hematopoietic stem cells, treatment with the 4-prolyl hydroxylase inhibitor FG-4497 stabilizes the HIF1 transcription factor and enhances cell mobilization (27). Furthermore, midkine and arachidonic acid induces HIF1α expression, critical for anti-apoptosis, IL-6 production, and proliferation in mESCs under hypoxia (28–30).
HIF1α regulation by hypoxia
HIF1α is tightly regulated by oxygen-dependent and oxygen-independent pathways (31–33). Unlike HIF1α, HIF1β is constitutively expressed (34). The suppressive effect of hypoxia on global gene transcription is reportedly induced by inhibitions against recruitment of RNA polymerase III and tRNA gene transcription, whereas HIF1A gene transcription is regulated by Sp1 and NF-κB under hypoxia (35–37). Tossato et al. showed that hypoxia treatment decreases HIF1A mRNA levels in patients with breast cancer (38). Moreover, Chamboredon et al. reported that hypoxia progressively decays HIF1A mRNA in endothelial cells via tristetraprolin, a mRNA-destabilizing protein (39). In stem cells, chronic hypoxia suppresses the expression of HIF1A mRNA in hESCs, but increases the expression of HIF1A mRNA and HIF1α protein in MSCs derived from rat bone marrow (40, 41). These findings suggest that the effect of hypoxia on HIF1A mRNA transcription genes differs relative to the type of stem cell impacted by hypoxia. In addition, previous studies have shown the regulatory role of the target of rapamycin (TOR) pathway in regulating HIF1 in mammals. Many studies have shown that activating the Akt/mammalian target of rapamycin (mTOR) pathway increases the cap-dependent mRNA translation of HIF1α (42–44). In addition to the capacity of mTOR to translationally upregulate HIF1α, mTOR has been reported to increase the stabilization of HIF1α and gene transcription activity of HIF1. Mint3 phosphorylation induced by mTOR stabilizes HIF1α by inactivating FIH (45). In addition, a regulatory protein of mTOR (Raptor), a subunit of the mTOR complex 1 (mTORC1), directly interacts with an mTOR signaling motif of HIF1α when upregulating HIF1-target gene transcription activity (46). However, the effect of hypoxia on mTOR has been debated. It is well documented that mTOR activity is reduced under hypoxic conditions by the tuberous sclerosis protein 1 and 2 complex and REDD1, a protein that regulates development and DNA damage response 1 is inactivated under hypoxic conditions (9, 47, 48). In contrast, several studies have shown that hypoxia-activated Akt induces mTOR signaling by enhancing vascular cell proliferation and angiogenesis (49). In addition, silencing HIF1A or inhibiting fatty acid synthase halts the hypoxia-induced mTORC1 signaling pathway in UCB-MSCs, suggesting that HIF1α-induced lipogenesis is critical for phosphorylation in the mTOR pathway (9). The activation of the Akt/mTORC1 pathway by HIF1α enhances the proliferation, migration, and survival of MSCs under hypoxia (9, 50). Under normoxic conditions, HIF1α interacts with the VHL protein, which subsequently stimulates ubiquitin-mediated degradation via hydroxylation of proline residues. Conversely, hypoxia stabilizes HIF1α in a VHL-dependent pathway and the stabilized HIF1α binds to the HRE for gene transcription (51).
Taken together, these findings indicate that HIF1α regulation has the capacity to determine the fate of stem cells and their bioactivity. However, the detailed mechanism by which HIF1α regulates stem cells (including processes of gene transcription, translation, stabilization, nuclear translocation, and transcriptional activation) remains poorly understood. This review discusses the recent understanding of the roles that various HIF1α regulatory factors play under hypoxic conditions, including the roles played by calcium, 2-oxoglutarate-dependent deoxygenase (2OGDD), microtubule network, importin α, and coactivators.
Go to : 
HIF1α Regulatory Factors
Calcium
Recent gene enrichment analysis data reveal that calcium-regulated calcineurin/NFATc4 signaling is a potential pathway regulating stemness in neural stem cells grown under hypoxic conditions (52). The upregulation of intracellular calcium levels is a response observed in many cell types exposed to hypoxia (53). Rat retinal progenitor cells and neural stem cells exposed to hypoxia exhibit increased intracellular calcium levels (54). The key role of calcium in adapting to hypoxia has been well documented; this occurs by the regulation of multiple signaling and gene expression processes (53). Recently, hypoxia-mediated calcium upregulation has been closely linked to the stimulation of HIF1A gene transcription, HIF1α translation, and HIF1α stabilization (16). One study has reported that the release of hypoxia-induced intracellular calcium increases the expression of HIF1α, which is further enhanced by pretreatment with ionomycin, an ionophore (55). Recently, Kim et al. showed that MSCs primed with both hypoxia and calcium enhanced stemness and the capacity for immunomodulatory activity, thereby attenuating graft-versus-host disease (56). This finding suggests that applying calcium to stem cells could be a promising strategy to enhance the efficacy of hypoxia-preconditioned stem cell transplants.
Previous studies have shown that HIF1α activates calcium signaling by inducing calcium channel expression. HIF1α induced by hypoxia and CoCl2 increases the expression of sodium-calcium exchanger-1, leading to intracellular calcium homeostasis and neuroprotection (57). Moreover, the regulatory effect of hypoxia on ER calcium sensors [including stromal interaction molecule 1 (STIM1)-induced store-operated Ca2+ entry (SOCE)] produces a positive feedback between HIF1α and STIM1 (58). Additionally, chronic hypoxia stimulates the expressions of calcium release-activated channels Orai1, Orai2 (but not Orai3) and STIM1 in pulmonary arterial smooth muscle cells. Furthermore, Orai2 expression depends on the presence of HIF1α (59). These findings indicate that the role of HIF1α in the expression of SOCE components is cell type-specific. Moreover, recent studies have shown that hypoxia-induced HIF1α increases the expression of transient receptor potential channel 1 (TRPC1) (60). Another study has shown that inducing pseudohypoxia by treating cells with CoCl2 or dimethyloxalylglycine N-(methoxyoxoacetyl)-glycine methyl ester (DMOG) increases the expression of Ca2+/Mn2+-transporting SPCA2, an isoform of the Golgi secretory pathway Ca2+-ATPase (61). Hypoxia and HIF1 induction with deferoxamine sometimes reduce the expression of sarcoplasmic reticulum calcium ATPase 2a in embryonic cardiac myocytes (62). Based on these findings, we conclude that hypoxia-induced HIF1α levels are closely associated with the regulation of calcium signaling in both intra- and extracellular pathways.
Calcium signaling is closely associated with HIF1α regulating processes, such as gene transcription, translation, and stabilization (Fig. 2). HIF1α induced by hypoxia increases STIM1 transcription-mediating SOCE, which then upregulates mRNA expression and stability of HIF1α by activating CAMKII-dependent p300 (58). Silencing the mitochondrial calcium uniporter (MCU) suppresses the expression of HIF1A mRNA independently of proteosomal degradation mediated by ubiquitin (38). A stable expression of the full-length TRPM2 channel, a member of the melastatin subfamily of TRP channels, increases gene transcription and stabilization of HIF1α, which is reversed by the stable expression of the short-length TRPM2 channel (63).
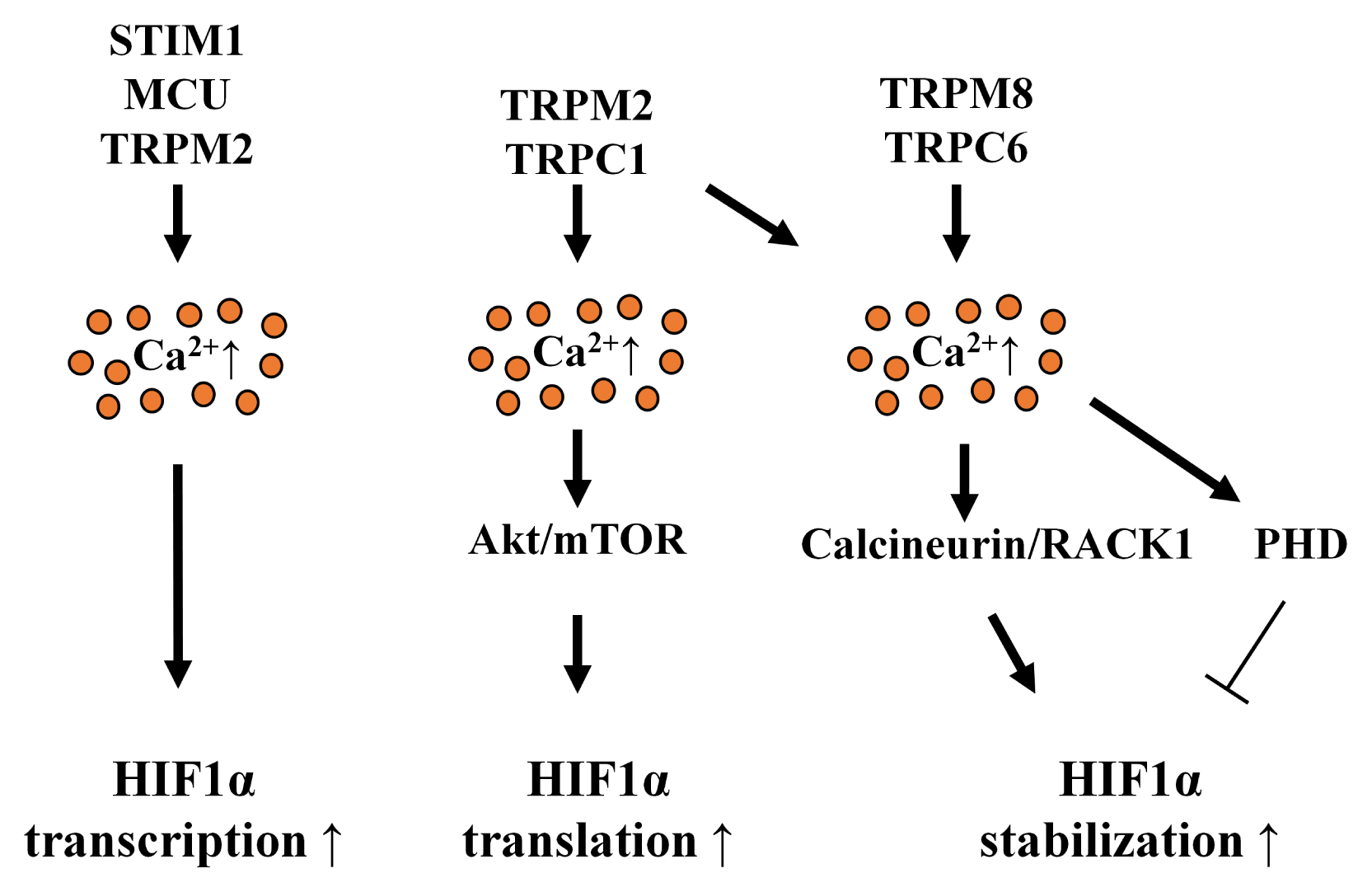 | Fig. 2Regulatory mechanism of calcium on HIF1α induction. Calcium channels regulating intracellular calcium levels induce HIF1α expression by inducing gene transcription, translation, and stabilization. STIM1, MCU, and TRPM2-activated intracellular calcium signaling increase HIF1A mRNA expression. TRPM2 and TRPC1 activate the Akt/mTORC1 pathway, which increases HIF1α translation. TRPM8 and TRPC6 stabilize HIF1α via the calcineurin/RACK pathway and by PHD-mediated prolyl hydroxylation, respectively.
|
Hypoxia-induced extracellular calcium influx stimulates HIF1α translation, which contributes to the upregulation of approximately 50% of HIF1α protein levels (64). Calcium-stimulated protein kinase C (PKC) α is required to translate mTOR-dependent HIF1α under hypoxia-suppressing global protein synthesis (64). In addition, the translation of HIF1α is controlled by activating mTORC1-mediated S6K1 (64, 65). Under normoxia, silencing TRPC1 attenuates the translation of HIF1α by inhibiting receptor-operated calcium re-entry (66). TRPC1 expression is increased during incubation under hypoxic conditions, which induces HIF1α protein levels via the Akt pathway. However, the effect of TRPC1 on HIF1α regulation is independent of HIF1A gene transcription and HIF1α stability (60). These studies indicate that TRPC1-induced calcium influx stimulates HIF1α gene translation under both normoxia and hypoxia. Most research in this area has focused on the role of specific hyper-activated calcium channels in cancer biology. However, only a few studies have described the relationship between calcium channels and HIF1 in stem cells. However, many stem cell studies have reported that intracellular calcium signaling plays a critical role in the somatic differentiation and migration of stem cells (67–69). These findings suggest that HIF1α can be induced by calcium signaling in stem cells under both normoxia and hypoxia.
In addition to clarifying the regulatory role of calcium in HIF1A gene transcription and HIF1α translation, previous studies have examined the relationship between calcium and HIF1α stability. Intracellular calcium chelation by BAPTA stops the interaction between VHL and HIF1 α through PHD inhibition, which further reduces the nuclear accumulation and binding of HIF1α and HIF1 to the gene promoter of carbonyl anhydrase 9 (70). Moreover, the overexpression of the thermo-sensitive calcium channel TRPM8 stabilizes HIF1α by inducing the oxygen-independent de-phosphorylation of RACK1 and inducing the binding of RACK1 to HIF1α and calcineurin, a calcium-dependent serine/threonine phosphatase (71). Calcineurin-induced RACK1 de-phosphorylation blocks the dimerization of RACK1, which further inhibits the oxygen-independent degradation of HIF1α by RACK1 (72). Silencing TRPC6 increases intracellular α-ketoglutarate levels, which promotes the prolyl hydroxylation of HIF1α under hypoxic conditions (73). However, the interplay between calcium and FIH remains poorly understood.
2OGDDs
PHD and FIH are oxygen-consuming enzymes belonging to the 2OGDD family of proteins. These proteins are capable of regulating the stability of HIF1α and epigenetics (74). In fact, 2-oxoglutarate (2OG), oxygen, and cofactors [such as a ferrous iron (Fe2+)] are necessary to activate the 2OGDD protein family (75). Succinate and fumarate (a succinate metabolite formed by succinate dehydrogenase) competes for the 2OG-binding site, which subsequently inactivates PHD and leads to the stabilization of HIF1 in embryonic stem cells under normoxic conditions (76–78). The succinate concentration in the mitochondrial matrix is lower than 0.5 mM under normoxia, but it increases to approximately 6 mM under hypoxia (79). Previous research has indicated the crucial role of the intracellular 2OG-to-succinate ratio in maintaining the pluripotency of embryonic stem cells. Indeed, naive mESCs exhibit high activities of intracellular 2OG and 2OGDD enzymes (80). Specifically, the addition of 2OG increases the pluripotency of mESCs, whereas treatment with succinate promotes the differentiation of mESCs (80, 81). Additionally, naive human pluripotent stem cells show high concentrations of 2OG, which appear to be critical in maintaining stemness (82).
Recent advances have been made in our understanding of the mechanisms involved in the stabilization of HIF1α regulated by 2OGDDs (Fig. 3). ROS produced from mitochondrial complex III represents PHD-inhibiting factors for stabilizing HIF1α under hypoxia (83). Mitochondrial ROS inhibits HIF1α activity via PHD2 dimerization, which is induced by the oxidation of cysteine residues required for Fe2+ and ascorbate (74, 84). Additionally, ferritin expression (stimulated by LPS) lowers intracellular Fe2+ levels, which inhibits PHD-mediated hydroxylation, resulting in the stabilization of HIF1α under normoxia (85). Under cellular hypoxic conditions, FIH (a 2OGDD protein) is more sensitive to peroxide and oxygen than to PHDs (86, 87). HIF1α is the most susceptible isoform of HIF1 when FIH is modified (88, 89). Deletion of FIH in mouse embryonic fibroblast cells stimulates glucose and oxidative metabolism, which increases oxygen availability under normoxia (90). Furthermore, levels of FIH expression are higher in skeletal muscles than in other tissues. The loss of FIH in skeletal muscle accelerates adaptation to hypoxia, mediated by HIF1, which enhances glycolysis and cellular metabolic responses to hypoxia (90). Moreover, ascorbate decreases HIF1 activity by reducing iron at the hydroxylase activation site in FIH, even after HIF1α is stabilized by 1% O2, DMOG, and CoCl2 (91). In contrast, ascorbate and ascorbate-2-phosphate have been shown to inhibit HIF1α stability in both PHD-or FIH-independent manner (92). Collectively, these findings imply that 2OGDD regulators (including 2OG, oxygen, Fe2+, and ascorbate) are potential targets for modulating the HIF1-mediated hypoxic response in stem cells.
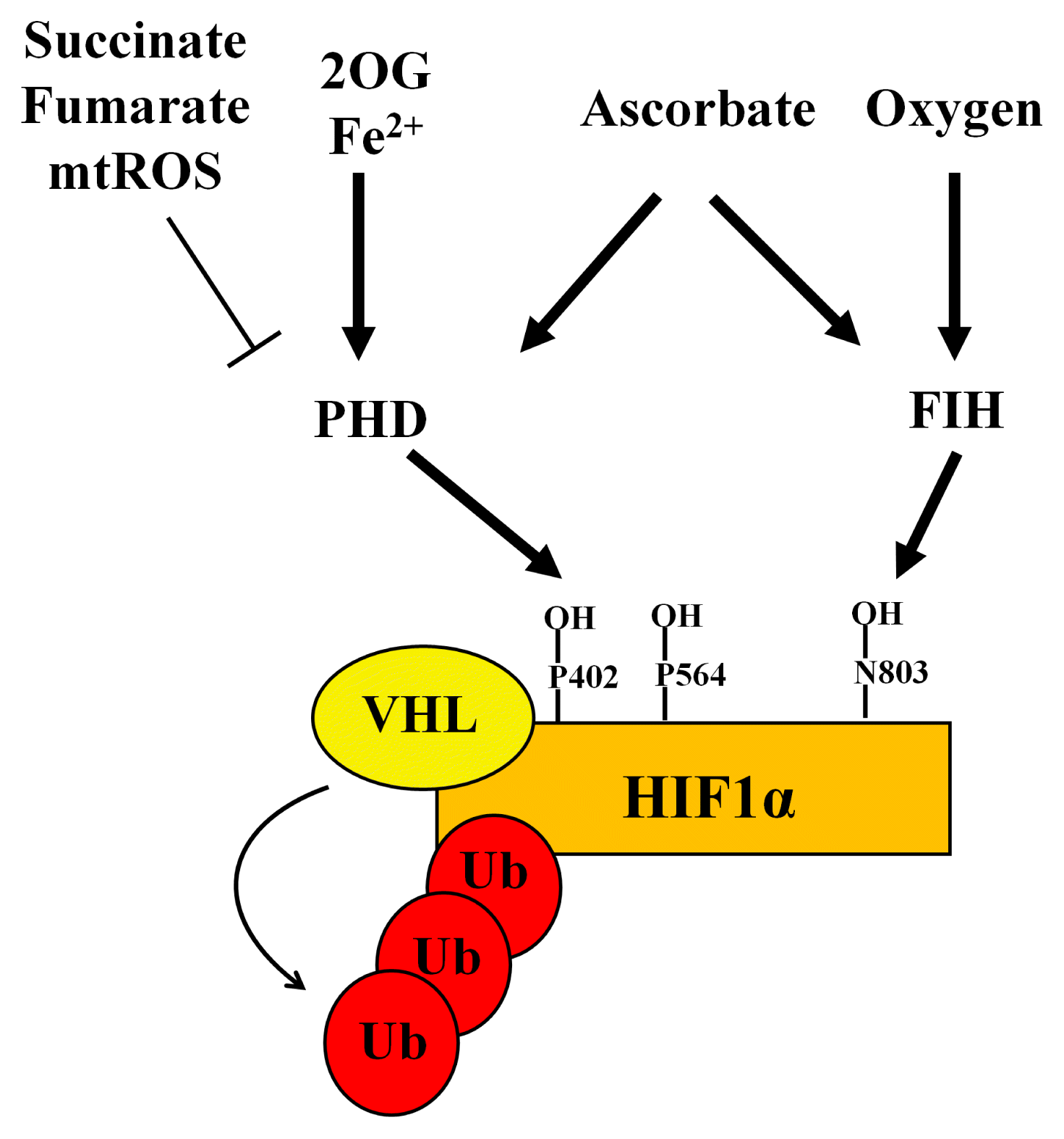 | Fig. 3Regulatory factors of 2OGDDs for HIF1α stabilization. Succinate, fumarate, and mitochondrial ROS inhibit PHD activation, leading to HIF1α stability. Conversely, 2OG, Fe2+, and ascorbate are required for PHD activation, followed by VHL-induced HIF1α ubiquitination. FIH is more sensitive to oxygen than VHL. Like PHD, ascorbate also activates FIH, which leads to asparaginyl hydroxylation, leading in turn to the destabilization of HIF1α.
|
Microtubule network and importin
Considering that HIF1α stability is upregulated by VHL inhibition under hypoxia, HIF1α nuclear translocation may be a potential target for enhancing HIF1 activity in hypoxic stem cells. However, the mechanism of the HIF1α nuclear translocation process in stem cells under hypoxia is poorly understood. It has been reported that HIF1α nuclear translocation is tightly regulated by microtubule network stability, cytoplasmic dynein activity, dynein adaptor proteins, and importins (Fig. 4). Although many investigators have studied the regulation of microtubule network under hypoxia, the effect of hypoxia on microtubule network is still controversial (93–96). Several studies have shown that hypoxia treatment stabilizes microtubule network, thus enhancing the chemoresistance of tumor cells (93, 94). Conversely, other studies have reported that hypoxia reduces microtubule stability and polymerization (95, 96). Despite these conflicting results, other studies have shown that stabilizing microtubules during hypoxia is critical for the nuclear translocation of HIF1α and for enhancing the chemoresistance of tumor cells (97–99). Chronic hypoxia suppresses the nuclear translocation of HIF1α by disrupting the microtubule network, which can be recovered by taxol (a microtubule stabilizer) pretreatment (97). In addition, stabilized microtubule inhibits Ran-mediated VHL nuclear translocation, leading to the stabilization of HIF1α in the nucleus (100). HIF1α interacts indirectly with microtubule via cytoplasmic dynein, a minus end-directed motor protein (99). Thus, the interaction between HIF1α and dynein is critical for HIF1α nuclear translocation. Dynein-associated cargo transport is regulated by dynein-interacting adaptor proteins, such as dynactin, LIS1, NUDEL, NUDE, Hook, and BICD (101–103). Additions of N-terminal fragment BICD2 and dynactin stabilize the formation of dynein–dynactin complexes, which enhance the processivity of dynein (104). In addition, recent our study demonstrated that BICD1 mediates interaction between dynein and HIF1α, critical for nuclear translocation of HIF1α in MSCs under hypoxia (105). Therefore, further investigations into the relationships between dynein adaptor proteins and HIF1α may provide new strategies to improve HIF1α activity, which could increase the efficiency of hypoxia adaptation in stem cells.
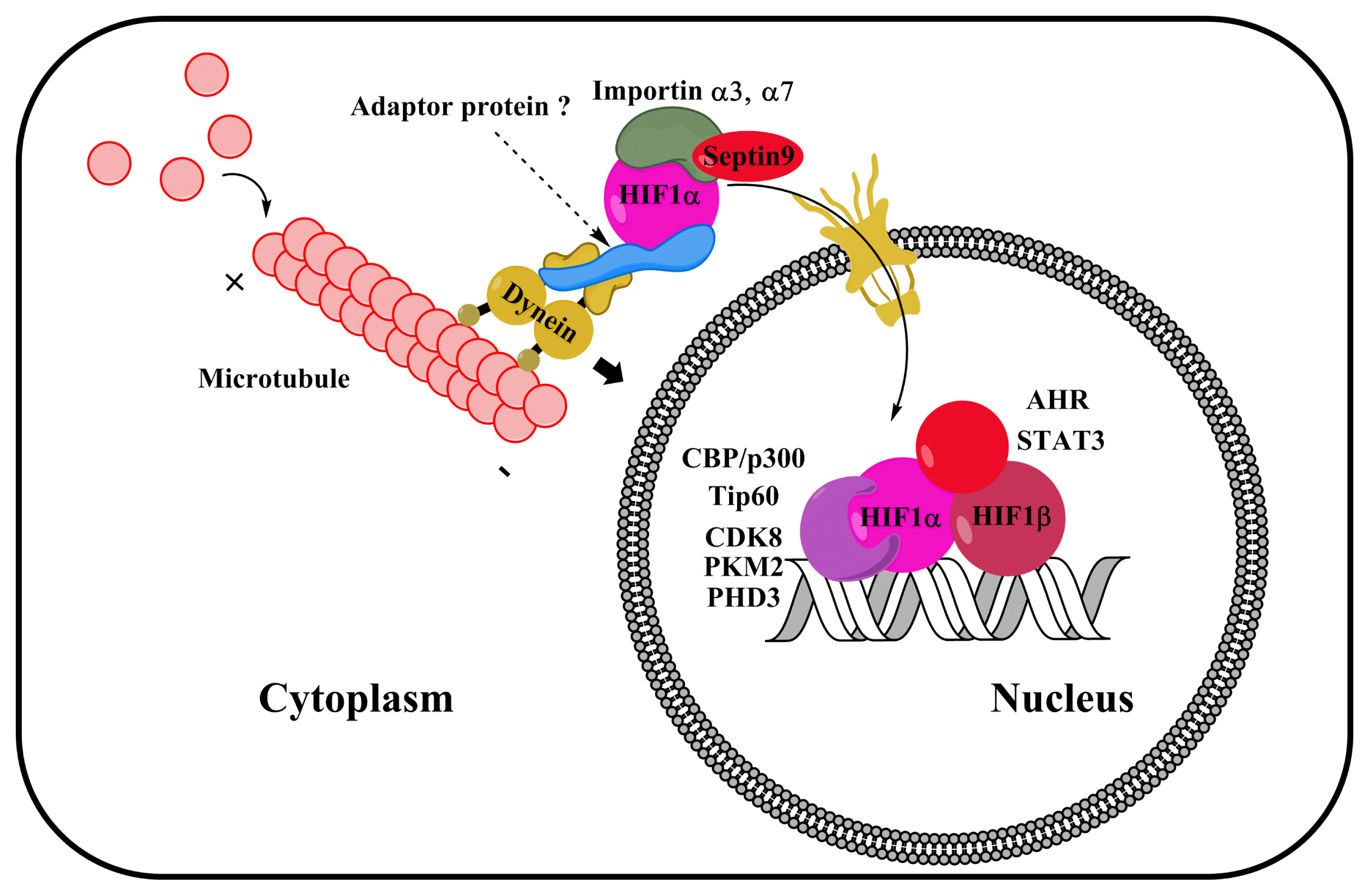 | Fig. 4Roles of microtubule network, importin, and coactivators in the nuclear translocation and activation of gene transcription in HIF1α. HIF1α nuclear translocation is regulated by microtubule stability and dynein activation. Interaction between HIF1α and importins α3 and α7 is important for the import of HIF1α into the cell nucleus. Dynein adaptor proteins (including BICD) may regulate the dynein-associated nuclear translocation of HIF1α. CBP/p300, Tip60, CDK8, PKM2, and PHD3 bind to HIF1α to coactivate gene transcription. Other gene transcription factors, such as STAT3 and AHR, also interact with HIF1α by synergistically activating HIF1-target genes expression.
|
Under hypoxia, the nuclear translocation of HIF1α depends on a nuclear localization signal (NLS) (106). Importin α binds to the NLS region of HIF1α, forming a heterodimer complex with importin β, which in turn allows HIF1α to dock with RanBP2, a component of nuclear pore complexes (107–109). It is well known that RanBP2, a SUMO E3 ligase, plays a key role in the nuclear importation of gene transcription factors (110). A major isotype of importin α that regulates the nuclear translocation of HIF1α appears to be cell type specific. One study reported that the NLS of HIF1α interacts with several types of importin α in U2OS osteosarcoma cells, including α1, α3, α5, and α7 (106). In addition, importin α3 is a major isotype regulating the nuclear translocation of HIF1α (106). Another study indicates that an importin α7 is required for the nuclear accumulation of HIF1α and the activity of HIF1 in HeLa cells (111). However, it is not yet known which isotypes of importin α drive the nuclear translocation of HIF1α in stem cells. In addition, a recent study of prostate cancer cells showed that isoform 1 of Septin 9, a GTP-binding protein, facilitates HIF1α nuclear translocation via its interaction with importin α7 (112). Likewise, calcium stimulates interactions between S100A6 and armadillo repeats of importin α, followed by the inhibition of NLS formation of cargo–importin α transport complexes that regulate the nuclear translocation of cargo proteins (113). These findings indicate that the interaction between importin α and HIF1 α improves the nuclear translocation of HIF1α.
HIF1 coactivators
Nuclear-translocated HIF1α interacts with various coactivators including CBP/p300, Tip60, CDK8, pyruvate kinase M2 (PKM2), PHD3, STAT3, and aryl hydrocarbon receptor (AHR) that are important in regulating HIF1 gene transcription activity (Fig. 4). In particular, lysine acetyl-transferase CBP/p300 is known to coactivate HIF1 gene transcription (114, 115). Mutational analysis has revealed that the CTAD of HIF1α is required for HIF1α to interact with CBP/p300, which is essential for the recruitment of SRC-1 to HIF1α under hypoxic conditions (116, 117). However, inhibiting the interaction between CBP/p300 and HIF1 via deletions of the first constant domain of CBP inhibits 35–50% of the HIF1-responsive genes, suggesting that CBP/p300 is not sufficient to activate HIF1 (118).
The Tip60 chromatin-modifying complex is regarded as a HIF1 coactivator and is required for HIF1α to activate RNA polymerase II and histone acetylation (119). Based on an analysis of transcriptome data, HIF1α employs TIP60, CDK8, or both as HIF1 coactivators. Their contributions to HIF1-responsive gene expression are >60% in colorectal cancer cells (119). Additionally, several studies have suggested that PKM2 and PHD3 are HIF1 coactivators. For example, Luo et al. reported that PKM2 mediates PHD3-dependent HIF1 transactivation and glycolytic reprogramming in HeLa cells (120). Consistent with this finding, Schoepflin et al. showed that PHD3 is a PKM2-independent HIF1 coactivator in pulposus cells of the cell nucleus (121). Collectively, these findings indicate that the major HIF1 coactivating partner differs according to the cell type. Therefore, further investigations are required to better understand the contribution of HIF1 coactivators in HIF1-target genes transcription in stem cells.
Recent studies have presented several gene transcription factors, such as STAT3 and AHR, as interacting partners with HIF1α (20, 122). In cancer stem cells, carboplatin-induced HIF1 activates signaling in calcium-dependent STAT3, thereby inducing pluripotency and enrichment in stem cells (123). Glioma stem-like cells in hypoxic conditions reveal high expression levels of vasorin (regulated by the HIF1α/STAT3 gene transcription complex), which is closely associated with tumorigenic capacity mediated by enhanced Notch1 signaling (20). Vasorin binds to Notch1 in the intracellular domain, which stimulates the production of Notch1 by γ-secretase (20). In addition, STAT3 and HIF1α synergistically stimulate the transcription levels of HIF1-target genes (124). Moreover, HIF1α has been reported to interact directly with the γ-secretase complex to regulate Notch signaling (125). These findings imply that HIF1α has both transcriptional and non-transcriptional roles in activating Notch signaling genes.
AHR is a transcription factor stimulated by various types of small molecules produced by gut flora during nutrient metabolism and by environmental stimuli (126). Similar to HIF1α, AHR also shares HIF1β as an interacting partner for dimerization and for activating gene transcription. Some studies have demonstrated that HIF1 β is essential for sustaining glycolysis in CD8+ effector T cells, suggesting a crosstalk between AHR and HIF1α (127, 128). HIF1α and AHR have also been reported to cooperate during the metabolic reprogramming of lymphocytes, including type 1 regulatory T cells and macrophages (129–131). Furthermore, kynurenine (a tryptophan metabolite) acts as an endogenous ligand to activate AHR (132). Kynurenine activates AHR via the PKC pathway, which stimulates the interaction between AHR and HIF1 β, which in turn upregulates gene transcription activity of HIF1 (122).
Go to :
Conclusions
Although several studies have advanced our understanding of HIF1α regulating processes, most research has focused only on the HIF1α regulating mechanism in cancer cells; however, additional studies on stem cells are needed to demonstrate the regulatory mechanism and physiological role of HIF1α. HIF1-mediated adaptation to hypoxia is essential in maintaining the biological functions and survival of transplanted stem cells. Therefore, HIF1α has the potential to improve the therapeutic efficacy of stem cell transplants. Recent studies have demonstrated that hypoxic preconditioning and HIF1α overexpression increases the transplant efficacy and immunomodulatory functions of stem cells (25, 41, 133). Moreover, oxygen-independent HIF1α induction via the regulation of calcium, 2OGDDs, microtubule network, and coactivators may provide a novel strategy to modulate and enhance the survival of transplanted stem cells. Therefore, a comprehensive understanding of the underlying mechanisms that affect HIF1α biology in stem cells will provide novel insights into stem cell biology and regenerative medicine.
Go to :
 XML Download
XML Download