

 PDF
PDF Citation
Citation Print
Print


Introduction
Mesenchymal stem cells (MSCs) are multipotent stem cells with main potential to differentiate into adipocytes, osteoblasts, chondrocytes (1, 2), and possible differentiation potential into hepatocytes, cardiomyocytes, skeletal myocytes and neurons (3). Recently, MSCs have been increasingly used in regenerative medicine because of their ability to treat tissue injury (4, 5), and their immunomodulatory properties (6–8); therefore, they have been part of clinical practice for more than 10 years (9) and were successfully isolated from a number of tissues including: bone marrow (10), adipose tissue (11, 12), umbilical cord blood (13), placenta (14), amniotic fluid (15, 16) in addition to umbilical cords (1). Umbilical cord tissue is one of the major sources of MSCs since isolation of umbilical cord MSCs (UCMSCs) is easier, does not require an invasive procedure and has a higher proliferation potential compared with other sources (2). Moreover, umbilical cord is usually discarded after birth so the collection of cells does not raise any major ethical concerns (17).
In standard culture conditions, MSCs are adherent to plastic surfaces of culture vessels and could be functionally isolated from tissues based on this ability. MSCs are also defined by their multi-differentiation potency and expression of cell surface markers (CD105, CD73, and CD90) and lack expression of (CD45, CD34, CD14) (18, 19). There are two main techniques to isolate MSCs from umbilical cord tissue: enzyme digestion and explants (2). Digestion-based protocols generally use enzymes like collagenase and hyaluronidase to digest tissues and release cells from the extracellular matrix of Wharton jelly, which then adhere to the surface of culture vessels (19). The main disadvantage of these protocols is that enzyme solutions are often poorly defined and have a broad range of incubation times (30 min~16 hours) depending on the purity and source of the enzymes. This raises the risk of degradation of cellular external lamina preventing MSCs from adhering to the surface of culture vessels after enzymatic digestion (2, 20).
On the other hand, explant protocols are cost-effective and simpler than enzymatic digestion protocols since they don’t require incubation with enzymes and depend on the MSCs ability to migrate from umbilical cord tissues and adhere to the surface of culture vessels (21). Explant protocols differ from each other by a number of factors that could affect the quality of isolation protocol such as: size of explants (22), type of culture vessels (plates, petri dishes or flasks), supplements added to culture media (21), and time required to change medium for first time (22–24). In this regard, many laboratories develop and optimize their own isolation and expansion protocol (20, 25).
Almost all protocols use fetal bovine serum (FBS) as a supplement in culture media as a source of hormones, attachment factors and other molecules required for isolation and expansion of MSCs in both research and clinical fields. However, FBS supplement medium has some limitations associated with clinical applications due to its animal source. These limitations include contamination risk by prions, bacteria, and viruses, and the potential for immunological reactions against xeno-genic components, in addition to high lot-to- lot variations (26, 27). In recent years, xenogenic -free and serum-free supplements such as human serum (28, 29), platelets lysate (27), cord blood serum (CBS) (30) and chemical defined media have been tested as FBS alternatives (31). Although, Serum-free media eliminate batch-to-batch variability they may still contain xenogenic proteins and has risk to transmitting infections. In addition, serum-free media are highly expensive compared with human blood derived supplement media; serum, plasma and platelet lysates (32).
CBS contains cytokines and growth factors which are required for proliferation and growth of stem cells such as (EGF, FGF, NGF, VEGF, PDGF, IGF, TGF, interleukins, interferons). It has been used as FBS substitute in culture media for animal MSCs, human bone-marrow- MSCs and umbilical cord blood derived MSCs (30, 33).
Optimization of simple, cost-effective and xenogenic-free protocols for isolating and expanding UCMSCs is an important factor in the successful application to deliver a sufficient number of cells to patients (20). Therefore, this study investigated the suitability of using pooled CBS as a replacement for FBS for developing suitable protocol to isolate UCMSCs by explant method without using digesting enzymes and in the expansion of UCMSCs.
Materials and Methods
Preparation of pooled CBS
After receiving the approval from Damascus University ethical committee, written informed consents were obtained from all participating pregnant women for donating their cord tissues and blood. Cord blood was collected from six donors after delivery using 50 ml sterile canonical tube without anti-coagulation. Blood then was allowed to clot for 2~4 h at room temperature. The clotted blood was centrifuged at 1,000 g at 20°C for 30 min. Sera were collected and pooled together to eliminate variations, then inactivated by heating at 56°C for 30 min. Finally, pooled cord blood serum was filtrated using 0.22-μm filters (Ministart, Sartourius Stediumbiotech, Germany), and was stored at −80°C.
Isolation of mesenchymal stem cell from umbilical cord tissue
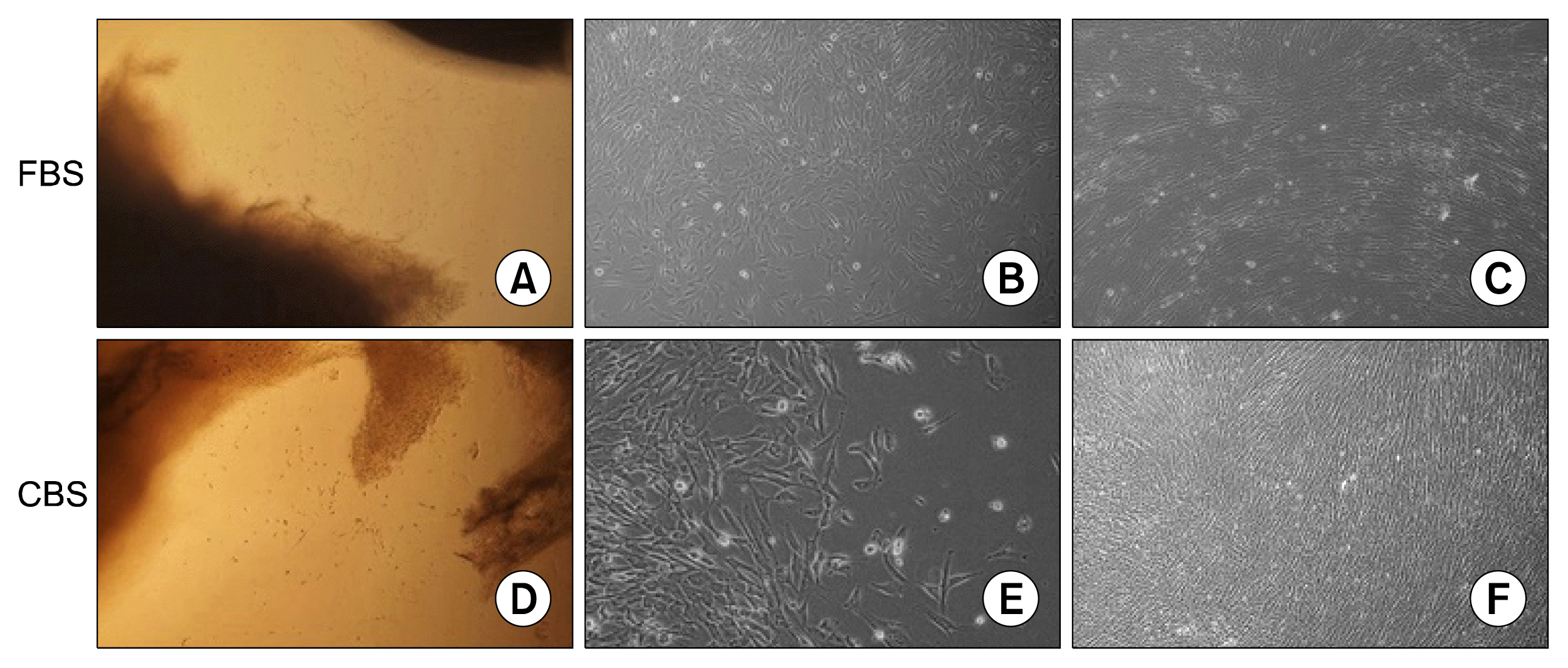
Three human umbilical cords were obtained from women with healthy pregnancies during caesarean deliveries at the end of gestation after signing informed consents. Umbilical cords (UCs) were collected in phosphate-buffered saline (PBS) (Euroclone, Italy) supplemented with antibiotics, 100 U/ml penicillin and 100 μg/ml streptomycin (Euroclone, Italy) and transported to the laboratory. UCs were washed with PBS under a sterile laminar flow cell culture hood and cut into 5 cm2 segments. The segments were cut longitudinally, and blood vessels were removed, and segments transferred to 25 cm2 flasks and incubated at 37°C in a humidified atmosphere with 5% CO2 in DMEM (Euroclone, Italy) supplemented with either 10% FBS (Euroclone, Italy) or 10% CBS. Flasks were left undisturbed for 7 days, after which the medium was changed for the first time and later was changed every 3~4 days. After 2 weeks, the UC explants were removed and the adherent cells were allowed to expand until they reach about 80% confluence. Morphology of isolated and expanded MSC cells was assessed using inverted microscope IX50 (Olympus, Japan) during the cell culture period.
Expansion and Proliferation kinetics
After reaching about 80% confluence, cells were sub-cultured using trypsin, EDTA 0.05% (Euroclone, Italy) to detach cells, then were centrifuged at 200 g for 5 min. Cells were counted using trypan blue exclusion and reseeded at 10,000/well density in 12 well plate in DMEM supplemented with either 10% FBS or 10% CBS. After 7 days, cells were trypsinized and counted. This procedure was repeated from passage 1 to passage 3. Population doubling time (PDT) was calculated for passage 1 and passage 3 according to the following equation:
where, ΔT: is culture period in hours, NH: number of viable cells at the end of passage and N0: number of plated cell.
The fold increase was also calculated according to the equation:
In vitro differentiation
Adipogenic and osteogenic differentiation
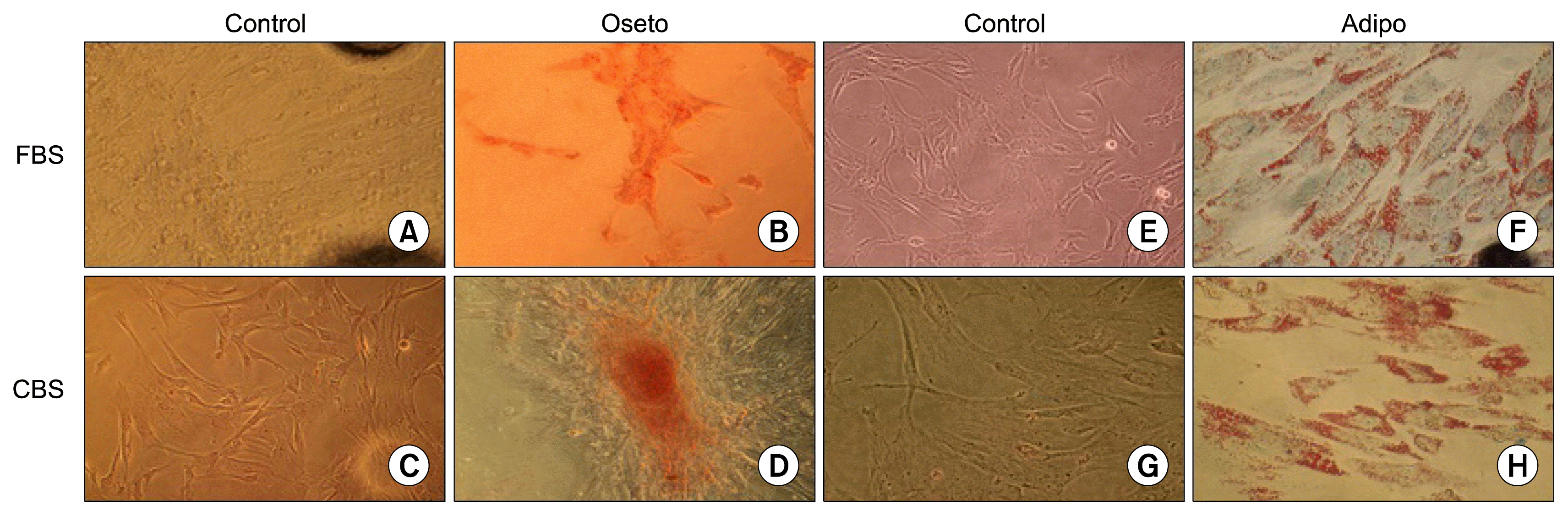
At the end of P2, cells were plated at 5,000/well in 96 well plate in (DMEM+10% CBS) or (DMEM+10% FBS) medium. After reaching 80~100% confluence, adipogenic differentiation medium (hMSC adipogenic differentiation kit, Euroclone, Italy) or osteogenic differentiation medium (hMSC osteogenic differentiation kit, Euroclone, Italy) were added, and media were changed every 3~4 days. After 3 weeks, cells were fixed with 10% formaldehyde. For adipogenic differentiation, lipid formation was assessed by staining with 2% sudan III for 5 min at room temperature then cells were washed with distilled water. For osteogenic differentiation, calcium deposits were detected by staining with 2% alizarin red (pH 4.2) for 20 min at room temperature followed by washing with distilled water. Lipid droplets and calcium deposits were detected by inverted microscope IX50 (Olympus, Japan). Control cells were maintained in (DMEM+10% CBS) or (DMEM+10% FBS).
Characterization of MSC
Molecular phenotyping
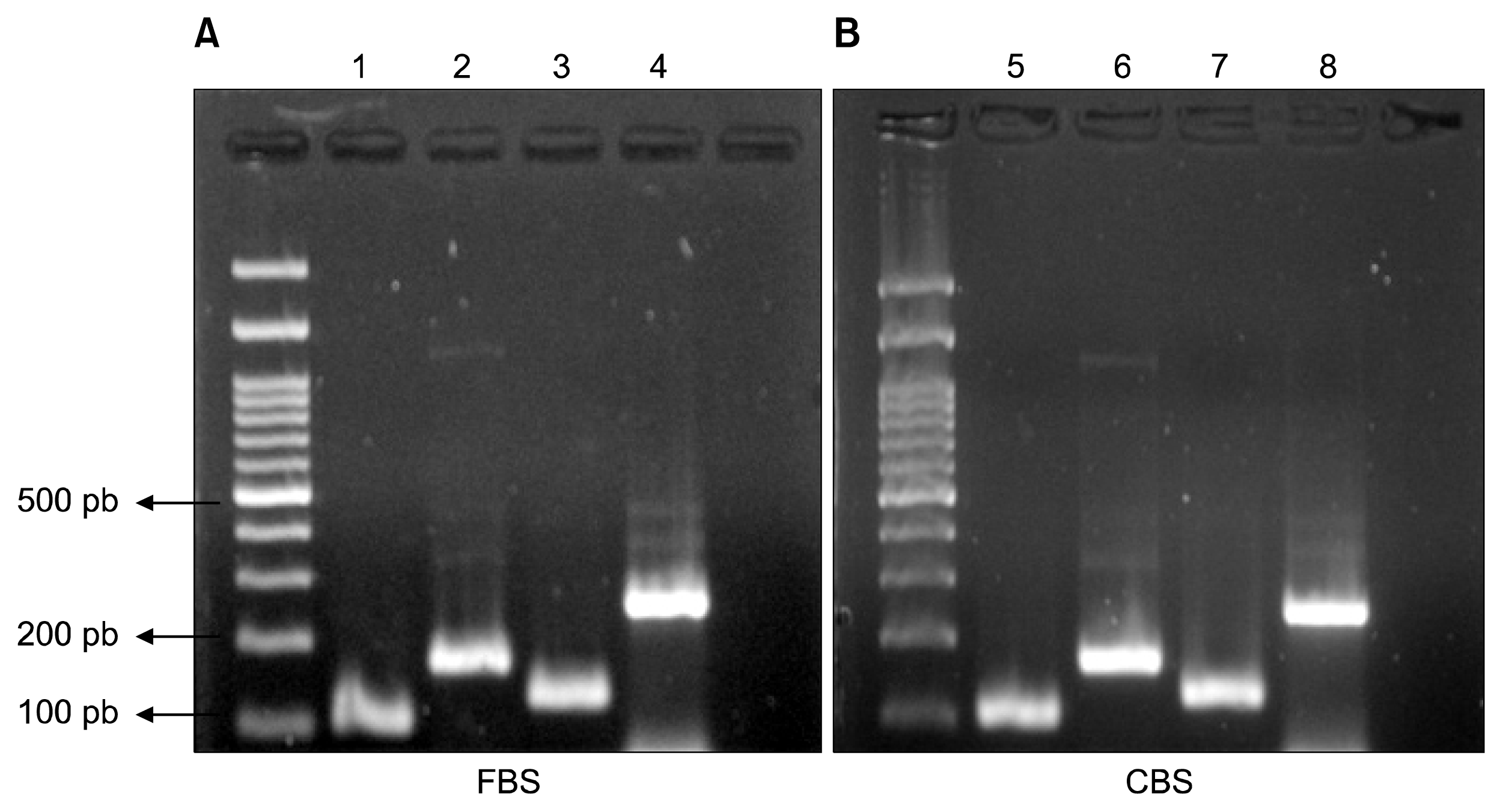
Total RNA was extracted from cells at end of P3 using (RNeasy Mini Kit–Qiagen, Germany). Reverse transcription was carried out using (RevertAid First Strand Synthesis Kit-Thermo fischer, USA) and RT-PCR reaction was performed using (Dream Taq, Thermo fischer, USA) to detect expression of CD44, CD90, CD105, using beta-actin gene expression as a control under the condition detailed in Table 1.
Primers that were used in RT-PCR reaction are shown in the Table 2.
2% agarose gel electrophoresis was performed to detect PCR products using 100 pb ladder (GeneDireX, Taiwan) and ethidium bromide (Carl Roth, Germany) to visualize amplicons.
Immunophenotypic characterization
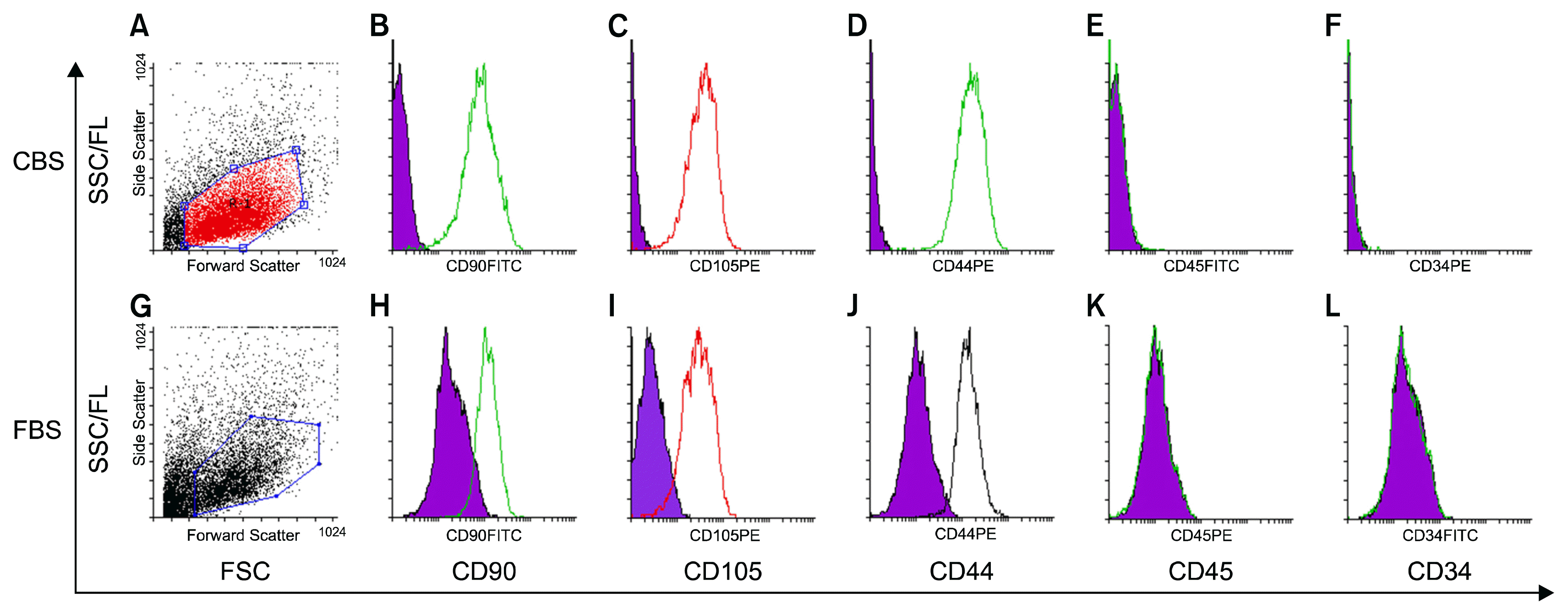
Cells at passage 2, isolated and expended in medium supplemented with either 10% CBS or 10% FBS, were trypsinized as described previously, washed with PBS and (5×105) cells were suspended and stained with antibodies: CD105-PE (Invitrogen, USA), CD90-FITC (Santa Cruz Biotechnology, USA), CD44-PE (BioRad, USA), CD34-PE (BD Bioscience, USA), CD45-FITC (Invitrogen, USA), CD14-FITC (BioRad, USA). Samples were analyzed using the FACS-Calibur (Becton Dickinson, USA) and DATA were analyzed with CellQuest software (Becton Dickinson, USA) and Flowing Software V2.5.1 (Turku Centre for Biotechnology, Finland).
Immunofluorescence
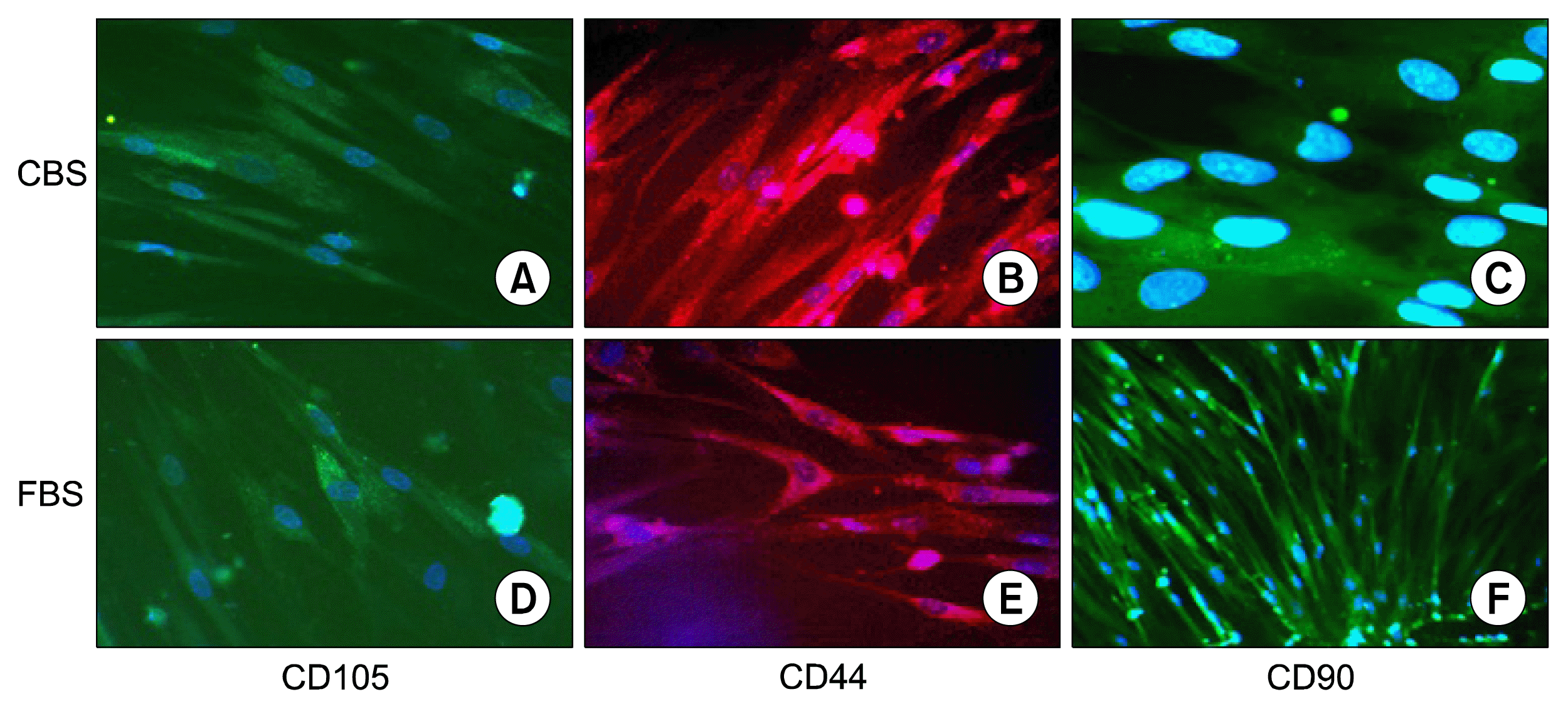
Cells were grown at chamber (chamber slide system, Labtek, USA) washed with PBS and fixed with 10% formaldehyde for 30 min. Blocking was performed by incubating fixed cells in blocking solution contains 2% bovine serum albumin (BSA) (Carl Roth, Germany) and 2% fetal bovine serum (FBS) (Sigma-Aldrich, Germany) for 1 hour. Then cells were incubated overnight at 4°C with antibodies against CD105-PE (Invitrogen, USA), CD90- FITC (Santa Cruz biotechnology, USA), CD44-FITC (BD Bioscience, USA). After washing, cell nuclei were counterstained with DAPI (4′,6-diamidino-2-phenylindole) 1 μg/ml for 5 min. Images were taken using Olympus fluorescent microscope (Olympus, Japan).
Results
Isolation of MSCs from umbilical cord tissue
After 7~12 days of explants culture, many cells migrated from explants, adhered to the surface of flasks, and were cultured in medium supplemented with either CBS or FBS. Explants were removed after 2 weeks and adherent cells had the ability to proliferate and expand, and were later subcultured when reaching 70~90% confluence. Adherent Cells isolated from UC tissues in medium supplemented with either 10% CBS or 10% FBS showed fibroblastic morphology (spindle- shaped) with multiple nucleoli which is a typical morphology of MSCs. Moreover, cells maintained their morphology when sub-cultured beyond passage 3 in medium supplemented with either CBS or FBS (Fig. 1).
Expansion and proliferation kinetics
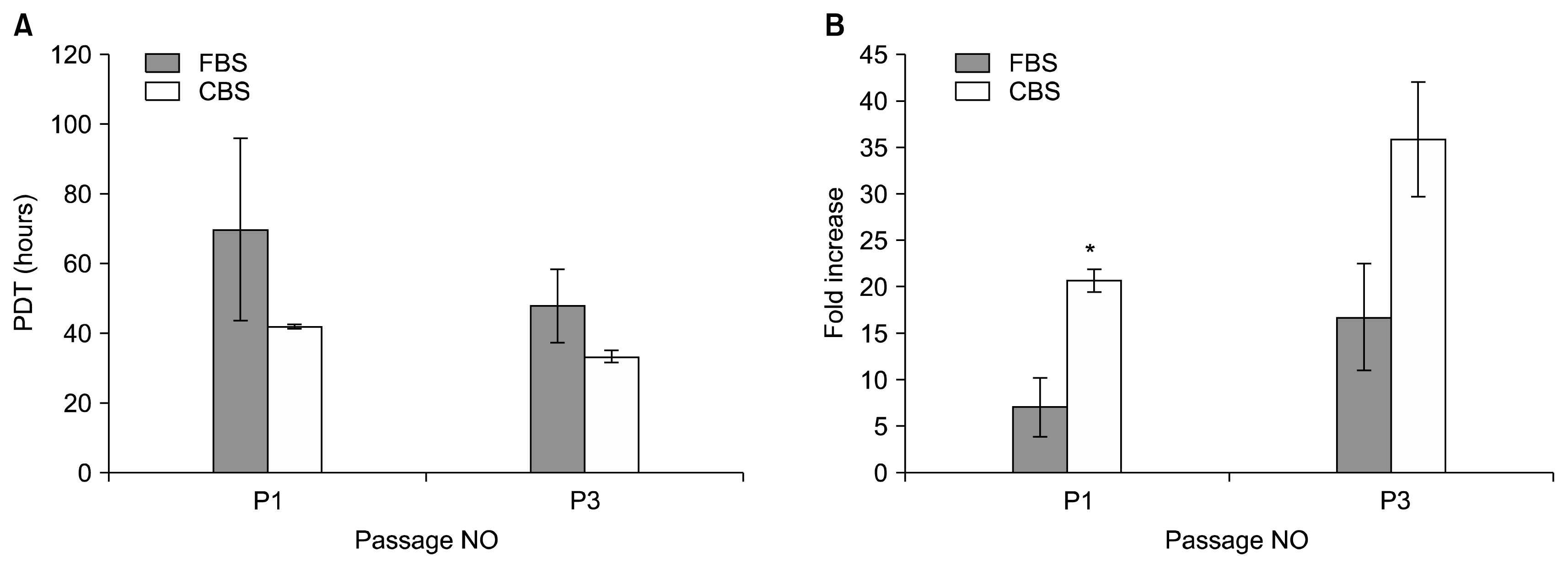
Population doubling time (PDT) for isolated MSCs using medium supplemented with either 10% CBS or 10% FBS did not change significantly between two media throughout culture period from passage 1 to 3 (p-value> 0.05), with lower PDT for cells cultured in medium supplement with 10% CBS than for cells cultured in medium supplement with 10% FBS. In addition, fold increase (FD) for cells cultured in medium supplement with 10% CBS was higher than for cells cultured in medium supplement with 10% FBS throughout culture period from passage 1 to 3 (p-value<0.05), with significance appeared only in passage 1 (Fig. 2).
Adipogenic and osteogenic differentiation
MSCs cultured in medium supplement with either CBS or FBS and directed to differentiate to adipocytes showed morphology changes and displayed the accumulation of lipid vacuoles, which stained positively with sudan III (Fig. 3F, 3H). In addition, MSC cultured in osteogenic differentiated medium showed morphology changes and stained in red color when stained with alizarin red, indicating the presence of calcium deposits for cultured cells in medium supplement with either CBS or FBS (Fig. 3B, 3D).
Molecular phenotyping
RNA was extracted from MSCs in P3 cultured in (DMEM, 10% CBS) and (DMEM, 10% FBS), after which RT-PCR reaction was performed. RT-PCR products for MSCs specific markers including CD105, CD44, and CD90 were visualized on gel electrophoresis (Fig. 4).
Immunophenotyping characterization
Immunophenotypic profile for surface markers of expanded cells in P3 was evaluated by flow cytometry. Cells cultured in (DMEM, 10% CBS) and (DMEM, 10% FBS) medium were positive for (CD90, CD105, CD44) and negative for (CD34, CD45) surface markers. Nonetheless, expression of (CD90, CD105, CD44) was considerably higher for cells cultured in CBS than those in FBS as shown in (Fig. 5).
Immunofluorescence
The expression of MSCs specific markers was also confirmed by immunofluorescence. Immunofluorescent analysis of MSCs cultured in CBS supplemented medium and FBS supplemented medium confirmed the expression of specific markers (CD105, CD44, and CD90) of MSCs cultured in either medium (Fig. 6).
Discussion
Mesenchymal stem cells are multipotent cells that were increasingly used in clinical studies (4). Two basic methods are implemented to isolate UCMSCs; explant and enzymatic methods that could differ from lab to lab (2). In addition, the majority of protocols still use FBS as supplement in culture media, which is considered the golden standard as medium supplement despite its numerous limitations (26).
In recent years, researchers have focused on optimizing conditions for isolation and expansion of MSCs for clinical trial proposes using xenogenic-free medium including serum-free medium and human product supplement media (34). Platelet lysate (17, 27), adult human serum (29), human plasma (28) and human umbilical cord serum have been used as alternative supplement of fetal bovine serum (30). Many groups have studied the effects of human blood-derived supplement medium on MSCs culture showing that these media could be useful in clinical grade MSC production by decreasing scale up time (13, 27, 35, 36). On the other hand, whole pieces of umbilical cord have also been cultured in human blood-derived supplement medium (23, 37) and in fetal bovine serum supplement media (38). However, a few researchers have studied the effect of umbilical cord blood serum on isolating UC tissue-derived mesenchymal stem cells by the explant method. Indeed, the effect of CBS was more studied on bone marrow derived MSCs than Umbilical cord tissue derived MSCs (30, 35, 39, 40).
In our study, we tested a simple protocol to isolate UCMSCs by explant method with minimal manipulation of UC by culturing 5 mm pieces of umbilical cord tissue without chopping the cord pieces.
Our results indicated that MSCs with fibroblastic morphology can be isolated from 5 cm2 umbilical cord pieces by its ability to migrate and adhere to plastic surfaces when cord pieces were cultured in CBS supplement medium, similar to their morphology when using FBS supplement medium in the same conditions (Fig. 1). Our results were similar to Khushnuma et al, who used a similar protocol to isolate UCMSCs, except for adding fibroblast growth factor (FGF) to culture media supplement with umbilical cord blood serum (41). In our study, we applied CBS as supplement and used explant method without adding any growth factors and showed that MSCs are easily and cost-effectively isolated from cord tissue. This indicates that using explant method combined with cord blood serum alone could be used as an alternative to FBS, beside the advantage of CBS as a xenogenic-free medium.
CBS also showed that it enhanced self-renewal and differentiation potential of bone marrow derived MSCs (40). We also showed that UCMSCs maintained their morphology and multipotent differentiation capacity, i.e., their ability to differentiate into adipocytes and osteocytes when they were cultured in CBS as well as FBS supplement media (Fig. 2).
MSCs isolated and expanded in CBS supplemented medium up to passage 3 had considerably lower population doubling times than for cells isolated and expanded in FBS supplemented medium and narrower range; PDT for cells in CBS ranged between 30 and 48 h, while PDT for cells in FBS ranged between 38 and 95 h). Moreover, MSCs cultured in CBS supplement medium had higher fold increase compared to MSCs cultured in FBS supplement medium with significant difference only in passage 1, and no difference between the two media in passage 3 possibly due to equivalent adaption of cells to either supplement media during elongated culturing (Fig. 3). Importantly, low population doubling times and high fold increase allows getting a large cell count of MSCs suitable for clinical application in short period, i.e., in early passages.
At molecular level, MSCs are characterized by expressing CD90, CD105 and CD44 surface markers, and lacking the expression of CD34 and CD45 (18). Our data from RT-PCR, flow cytometry, and immunofluorescence collectively showed that isolated and expanded cells from umbilical cord tissue using explant method and either CBS supplement medium or FBS supplement medium corresponded well with the standard positive and negative molecular profile, indicating that explant method combined with CBS–supplement media did not alter expression of MSCs surface markers during isolation and expansion period (Figs. 4~6). Interestingly, expressing of CD90, CD105 and CD44 surface markers were considerably higher in CBS than FBS supplement media, possibly indicating a favored profile when culturing cells in CBS (Fig. 5).
Conclusions
Our data strongly support the plausibility of using explant method combined with pooled CBS as alternatives of enzyme digestion and FBS in the clinical setup as this method is simple and cost-effective to isolate, culture and expand UCMSCs, maintaining their stemness characteristics including self-renewal ability, multi-differentiation potential, and unique surface markers. The simplicity and easiness of this protocol makes it favorable for basic and clinical research as well as training schemes.
 XML Download
XML Download