

 PDF
PDF Citation
Citation Print
Print


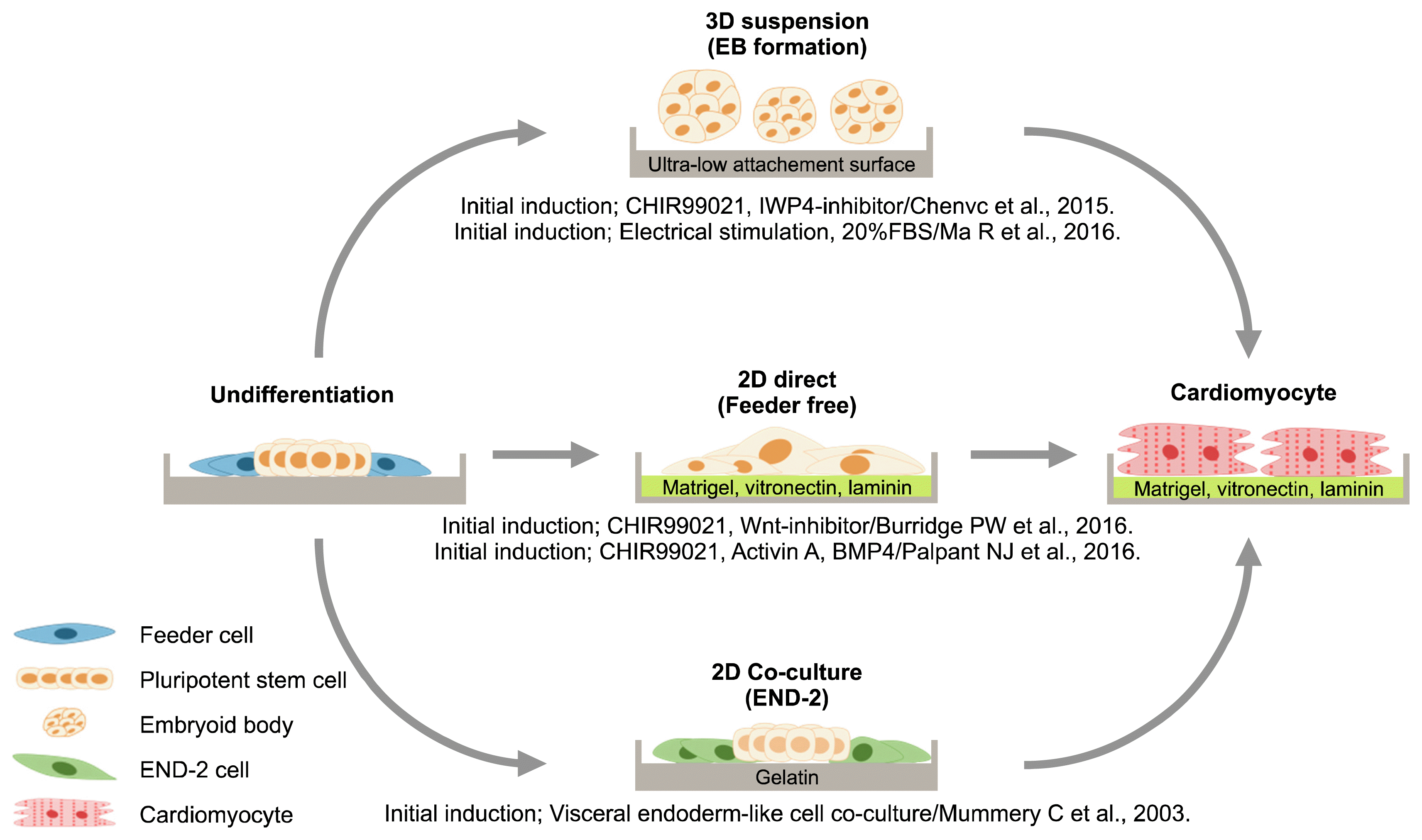
Human pluripotent stem cells derived cardiomyocytes (hPSC-CMs) differentiation strategies
Human embryonic stem cells (hESCs), induced pluripotent stem cells (hiPSCs) and nuclear transfer embryonic stem cells (Nt-ES), are all considered to possess pluripotency, although their derivation methods and origins differ. Such a capacity could provide an answer to overcoming the lack of cellular sources from which functional cardiomyocytes (CMs) can be obtained. In the last two decades, strenuous research efforts have made huge stride in developing efficient differentiation and purification methods for CMs, and their potential use in the areas of cell therapy development and new drug discovery is steadily being materialized. Recapitulating key developmental events of cardiogenesis has been a customary practice for inducing CMs differentiation. Such attempts have been conducted 1) through formation of 3-dimensional floating cell aggregates (embryoid bodies, EBs) that consist of three germ layers cells, 2) by directly supplementing differentiation milieu to monolayer of adherent hPSCs or 3) by co-culturing with supporting cells known to facilitate CMs differentiation (Fig. 1).
3D method using embryoid body (EB) formation
Since the establishment of hESCs lines in 1998, and later hiPSCs in 2007, EBs formation has been a popular choice of method for inducing CMs differentiation. Floating hPSCs colonies spontaneously form spherical EBs that constitute cells representative of all three germ layers. Supplementation of a cocktail of growth factors to the suspension culture of EBs expedites CMs differentiation as some cells undergo perceptible morphological changes into contractile cells with sarcomeric structures as early as day 20 post growth factor treatment (1). However, the attempts to control the consistency and efficiency of EBs based approaches are severely thwarted mainly due to the variability in the EB sizes. In recent years though, studies have reported various means to control the EB sizes in order to minimize differentiation variability associated with the size. For instance, micro aggrewell plates (commercialized by STEMCELL Technologies Inc), allow aggregation of a defined number of cells by simple centrifugation. The ease with which a large number of uniform size EBs can be produced paves the way for a simple and commercial scale production of differentiated cells (2).
2D-directed monolayer differentiation
Monolayer differentiation of hPSCs is initiated without the intermediately EB formation stage, and differentiation cues are directly supplemented to the adherent hPSCs in feeder-free systems. One of the advantages of the monolayer differentiation is that it enables investigators to gain a better control of the differentiation. The morphological changes are easily observable under a light microscope therefore different growth factors can be supplied in a stage-specific manner in attempts to as closely recapitulate the embryonic environment as possible. 2D directed differentiation of CMs is often based on Wnt signaling pathway, and regulates the differentiation by adding growth factors such bone morphogenetic protein 4 (BMP4), Activin A and fibroblast growth factor 2 (FGF2) to the culture medium containing serum (3). Such method is, however, not suitable for clinical application of resulting CMs as the serum is an animal derivative. Xeno-free culture medium consisting of RPMI and B27 supplement has been developed, and together with above-mentioned growth factors, CMs differentiation efficiency between 50~90% was achieved (4). In recent years, small molecules, known for higher potency and lower manufacturing costs than growth factors, have been widely adopted for initiating CMs differentiation. GSK3-inbitiors (CHIR99021, BIO, TWS119, TDZE-8, 3F8) and wnt inhibitors (IWR-1, IWP2, IWP3, IWP-4, XAV-939, KY02111, WNt-C59) are commonly used small molecules for CMs differentiation yielding an efficiency between 80~90% (5, 6).
Co-culture system
CMs differentiation is induced by culturing hPSCs with endoderm lineage cells that are known to play intricate, yet important roles for the heart development. A co-culture system of hPSCs with mouse visceral endoderm-like cells (END-2) has been reported to be conducive to generating contractile cells at an efficiency of about 25 to 40%. Co-culture differentiation methods can induce differentiation using a small number of hPSCs, and has an advantage of achieving comparable differentiation efficiency within a relatively short period of time without a plethora of growth factors (7). However, there is a disadvantage of having to separate differentiated CMs from the residual END-2 for subsequent applications.
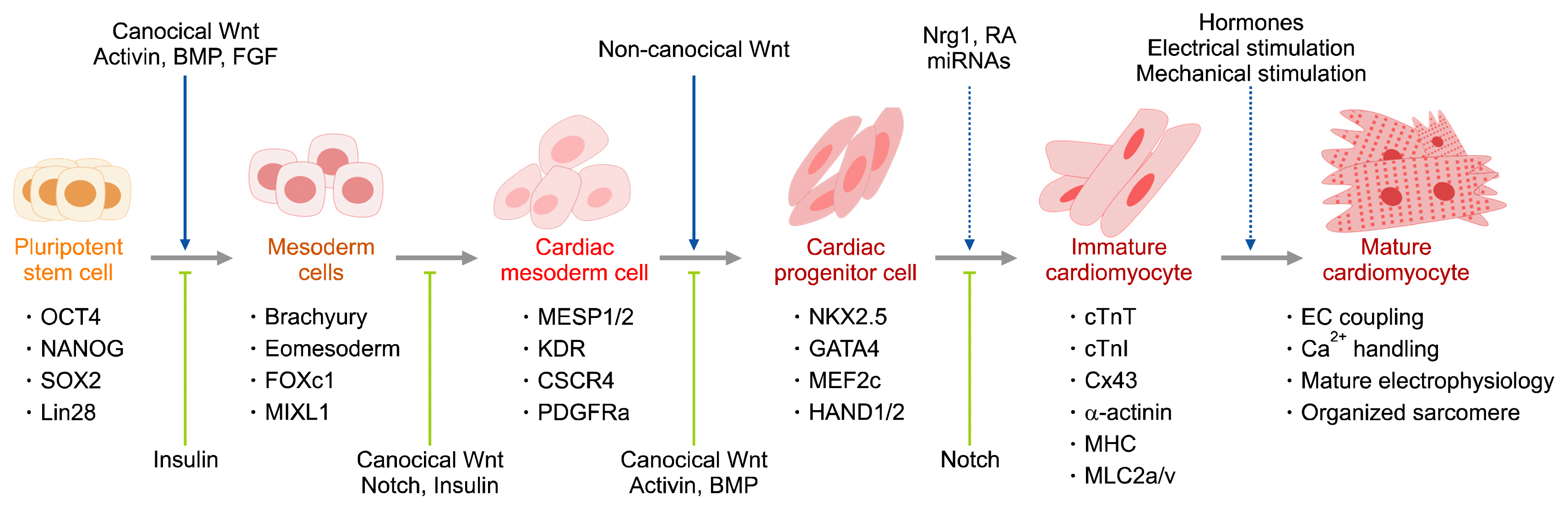
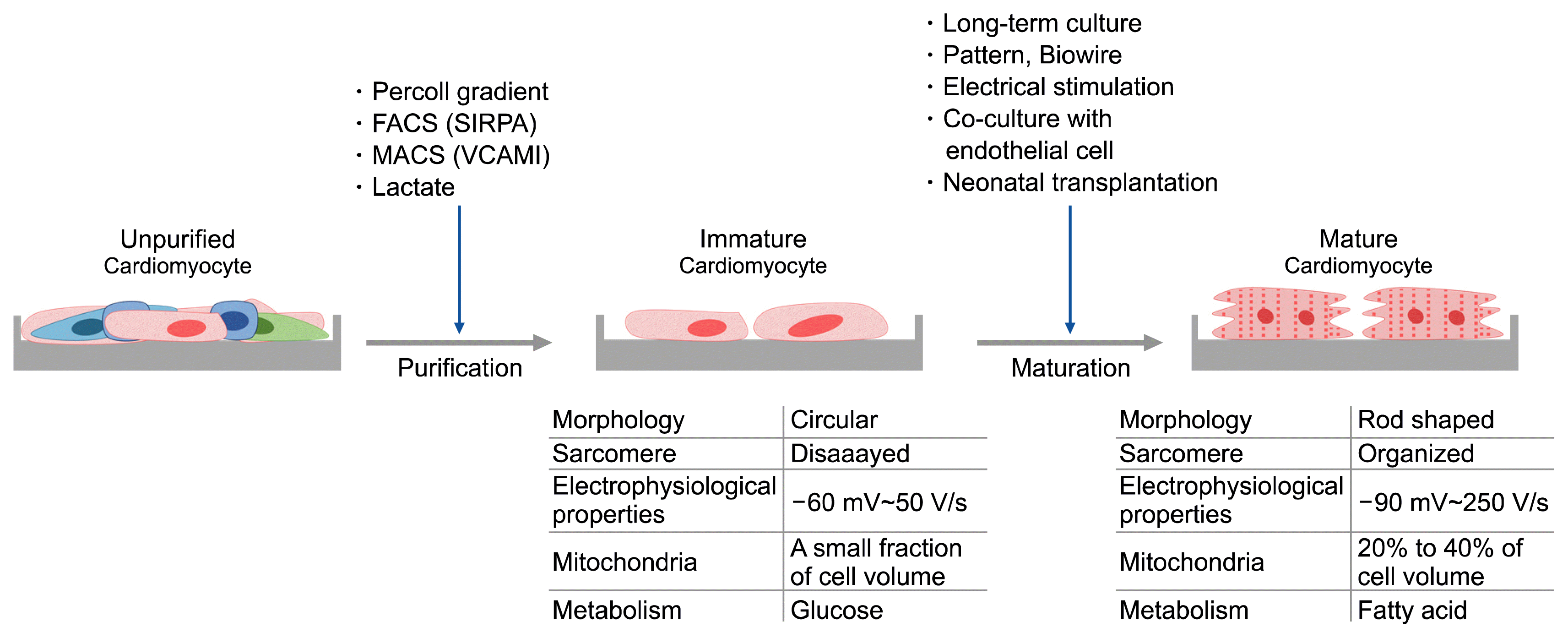
Characterization and purification of hPSC-CMs
hPSC-CMs emerges sequentially from the undifferentiated state through the primitive mesoderm to CMs in an analogous manner to cardiogenesis during the early human developmental stage. In the initial stages of the differentiation, the expression of undifferentiated markers (e.g. Oct4 and Nanog) dissipates while inversely, the expression level of representative markers of early mesoderm differentiation increases (e.g. Brachyury and DKK-1). As the differentiation progresses, the differentiating cells take up immature CMs characteristics as demonstrated by CMs (e.g. NKX2.5 and Gata4) and muscle filament (e.g. cTnT, alpha-actin and MHC) associated markers expressions, spontaneous contractility and electrophysiological properties (Fig. 2) (8). Over time, such cells mature into fully functioning cells akin to adult CMs (1). Since the differentiation is never 100% efficient, the presence of contaminating by-products is inevitable, thus techniques for separating and purifying CMs are required (9). Current purification strategies, and probably one of the most used methods, include the following; centrifugation, flow cytometry and metabolic screening (Fig. 3).
Centrifugation (percoll gradient)
CMs purification by density gradient is somewhat considered as the “golden standard”. Since CMs are larger and denser than non-CMs, density gradient centrifugation, in which cells are fractionated by floating density, could effectively separate CMs from other cell types. This was routinely used to isolate primary CMs until 2000, when mouse ESCs derived CMs were first purified by the same method (10). Later, ~60% hPSC-CMs were purified from impurities such as EBs, small clumps and single cells, and ~82.6% CMs were enriched following a 2D directed method (11, 12).
Flow cytometry
Flow cytometry analysis using cell-specific markers has the advantage of being applicable to various cell types in a simple and highly efficient manner, but no marker specific to CMs has been reported. For this reason, many researchers have identified CMs related surface proteins during differentiation, and validated their authenticity as CMs specific markers. KDR and C-KIT (13) and Flk-1 and PDGFR- α (14) have been used to isolate hPSC-CMs progenitors, which were then further matured into functional CMs. Such purification methods yielded populations of 50% and 60% CMs respectively (13, 14). In 2011, signal-regulatory protein alpha (SIRPA) was found to be exclusively expressed in hPSC-CMs, and when an antibody against SIRPA was used to isolate CMs from the differentiating culture, it yielded up to 98% cTnT-positive cells (15). Vascular cell adhesion molecule 1 (VCAM1) has also been reported as a viable surface protein marker for isolating hPSC-CMs (16). After screening 242 antibodies for human cell surface molecules, it was found that 80% cTnT positive hPSC-CMs also expressed VCAM1 11 days post differentiation. Thus, purification using VCAM1 antibody at day 11 yielded ~98% VCAM1 positive population of which ~95% expressed cTnT, and displayed molecular and functional features of CMs.
Gene modification
Various reporter hPSCs lines have been generated to develop an efficient fluorescent-based purification method. The generation of a reporter hPSCs line harboring the green flourescence protein (GFP) Nkx2.5 gene was reported, and purification based on a high level of GFP fluorescence upon differentiation yielded 85% CMs (17, 18). The α-myosin heavy chain (αMHC) gene is broadly expressed during the course of murine cardiac development therefore its sensitivity for purifying hPSC-CMs was examined by constructing a myocardial-specific enhanced GFP reporter in H9 hESCs line (19). Using the cell line, the investigators were able to isolate myocardial precursors, which were capable of further developing into atrial- and ventricular-like CMs. CMs specific reporter cell lines serve the purpose of purifying CMs, but this method is an unlikely option for clinical applications.
Metabolic selection
Glucose is the primary energy source for metabolically active mammalian cells. Glycolysis breaks down glucose into pyruvate and lactate while generating two molecules of ATP without the presence of oxygen. Pyruvate is then used as a substrate in the mitochondrial tricarboxylic acid cycle to yield 36 ATP molecules via oxidative phosphorylation. CMs, unlike most somatic cells, are capable of using lactic acid as an alternative metabolic substrate under glucose-depleted environments. To attest this metabolic homeostasis, differentiating hPSCs culture was subjected to glucose-depleted environments with varying concentrations of lactate (20). Therein, non-CMs failed to survive as ~98% alpha-actinin positive cells were detected 20~30 days after differentiation. The fact that hPSC-CMs had remained a high intracellular ATP in such environments revealed that the metabolic selection is an effective purification strategy for enriching viable and functional hPSC-CMs.
Maturation methods for hPSC-CMs
The action potential generated by ion channel activity in intracellular segmental structures is important for the spontaneous contractility of mature CMs. However, hPSC-CMs are immature in comparison to those derived from adult tissues in terms biological (cell morphology and contractility) and functional (cellular metabolism and electrophysiology) standpoint. Therefore, for the application of hPSC-CMs in cell therapy development and drug screening, the cells are required to resemble as closely as possible the fully fledged adult CMs in terms of functionality. Techniques such as 1) long-term culture, 2) biomimetic and tissue engineering techniques, and 3) electrical stimulation have been employed as methods to induce maturation of hPSC-CMs (Fig. 3).
Characteristic comparison between immature hPSC-CMs and mature adult CMs
Morphological properties
Although mature human adult CMs have a long rod-shaped arrangement of sarcomere structures, hPSC-CMs are smaller and have an irregular round segmental shape compared to mature CMs. The membrane capacitance, which increases in proportion to the cell surface area, is lower than adult CMs due to the size of immature CMs (21). In adult CMs, transverse tubules (T-tubules) that regulate the calcium release and contraction of the sarcoplasmic reticulum (SR) are well developed, but those in hPSC-CMs are poorly developed in comparison (22).
Electrophysiological properties
In order to generate action potentials of CMs, signal transmission systems between various ion channels must work in concert. However, in immature CMs, low activity among ion channels may cause impairment to spontaneous contractility, and unstable cell function may present crucial pitfalls in their applications. The gap junction, which regulates the conduction velocity of CMs, is an important factor in determining the maturation of CMs. In the case of immature CMs, Connexin and N-cadherin are distributed circumferentially around the cell membrane, while those in mature CMs are distributed in the intercalated disks (23). Since the difference in the intracellular distribution of the gap junction proteins may attribute the functional differences of CMs, modulating the distribution of the gap junction protein is a very important for the stabilization of CMs function.
Metabolic properties
CMs are one of the most energy-demanding cells in the human body, and over 35% of their volume is composed of mitochondria. The type of energy metabolism of CMs depends on their maturity. Not only mature adult CMs have higher mitochondrial numbers compared to hPSC-CMs, but the inner membranes of the mitochondria are also well developed. The mitochondrial oxidative capacity increases in the mature CMs, and ~90% of energy generation is ascribed to fatty acid β-oxidation (24). Immature and proliferating CMs, in contrast, have fewer mitochondrial numbers, and they are functionally under developed. Indeed, oxidative metabolism only accounts for only <15% of the total ATP produced, and their reliance on glycolysis is much preferred (25).
Maturation method for hPSC-CMs
Long term culture
hPSC-CMs are generated 20~30 days post induction thus, it is inevitable that they are biologically, functionally and structurally immature in comparison to adult CMs that continue to mature for 6~10 years after birth. A long-term culture protocol was devised to examine whether hPSC-CMs take up more mature characteristics over a prolonged period of time in culture (26). In this study, the early stage hPSC-CMs (20~40 days after initiating differentiation) were largely small and rounded cells with poor subcellular organizations, and an underdeveloped contractile machinery as evidenced by the misalignment of myofibrils. Morphological changes into more elongated shape were accompanied with an increased size over the ensuing 2 months, and by day 80~100 post differentiation induction, an improved myofibril alignment, density and morphology was observed. In addition, a 2-fold increase in the contractile ability was evident in the late stage hPSC-CMs when sarcomere length changes were measured during the contraction. Other functional characteristic parameters such as calcium uptake, action potential properties and cardiac gene expression profile of the late-stage hPSC-CMs closely resembled that of adult CMs.
Mimicking in vivo niche using tissue engineering approach
Because human cardiac muscle cells are exposed to pressure and electrical stimulation in vivo, they are structurally rod-shaped to minimize the impact. A method of improving the maturation of CMs through a cell culture system mimicking the natural environment is studied. As a result of culturing immature CMs by changing the surface of the cell culture dish to a patterned topography, a linear of myofibril was developed in the same direction as that of the substrate, and the distribution of the gap junction and the function of the conduction velocity improved (27). CMs are composed of 3D arrangement of interstitial fibroblast, blood vessel, extracellular matrix (ECM) and CMs (28). Therefore, in order to regulate the maturation of immature CMs, a three-dimensional extracellular matrix capable of simulating the environment similar to myocardial tissue was prepared by tissue engineering techniques, which improved maturation of hPSC-CMs.
Electric stimulation
CMs form a synchronic structure with tissues with high electrical signal activity. In order to regulate the maturity of immature CMs by taking advantage of this property, the cells were cultured under the conditions of electric stimulation, and as a result, stable cell arrangement, intercellular synchronization and contractile capacity were all increased. In addition, the structure and function of mitochondria, sodium-calcium exchanger, action potential and conduction velocity were also improved (29).
Application of hPSC-CMs
It is widely recognized that hPSC-CMs can be used in the following applications; 1) disease modeling, 2) cell therapy development for treating acute myocardial infarction, 3) human cell based assays for arrhythmogenesis and cardiotoxicity evaluation of candidate molecules for drug discovery.
Cardiac disease modeling
hPSCs derived from patients with genetic heart diseases reflect individual genetic information, and CMs generated from such cells provide an accurate in vitro model of the diseases. This allows you gain sights into the cause of disorder and its consequences. Pathogenic studies of the human heart disease using an animal disease model has limitations in its accuracy because of differences in cell size, heart rate, and ion channel activity between the two species. Since the inception of iPSCs technology, CMs from patients’ own iPSCs have been used to elucidate pathophysiology of representative genetic cardiac diseases such as Leopard syndrome, long QT syndrome, Timothy syndrome, Pompe disease, familial hypertrophic and dilated cardiomyopathy, and the genetic analysis and characterization of electrophysiological functions have been reported to reflect the characteristics of the diseases (30–36). Therefore, it is expected that cardiac disease modeling using hiPSC-CMs will be able to illuminate unseen pathological mechanisms of heart diseases for developing improved and efficient treatment methods.
Cell therapy
The first approved stem cell therapeutic product in the world, Hearticellgram-AMI is a bone marrow derived mesenchymal stem cells (MSCs) based therapy for treating acute myocardial infarction (47). A profusion of other clinical trials is currently taking place around the world to validate the feasibility of various other types of MSCs for treating myocardial infarction. In most of the trials, the safety of such therapies is consistently confirmed, but we are often left perplexed regarding to their efficacy. In the rare cases of therapeutic outcomes, the mode of therapeutic action is revealed to be indirect; through the paracrine effect of the stem cells secretory factors (48). Since the donor cells are expected to live only for a short period of time after transplantation, the potency of the paracrine effect remains questionable (49). The potential of using hPSC-CMs for treating myocardial infarction is manifested by pre-clinical and clinical studies (Table 1). In contrast to adult/fetal tissues derived MSCs, the transplanted hPSC-CMs functionally integrated into the injured cardiac tissue, and improved the function of cardiac muscles by synchronizing with the heart tissue of the recipient. The integrated cells remained viable up to 6 months post transplantation, and the observed functional improvement was also sustained for the same period of time. A significant milestone in PSCs research was reached when hPSC–CMs received the regulatory clearance to initiate human clinical trials for treating acute myocardial infarction in 2015 (46).
Toxicity testing and drug screening
Due to the difficulties associated with acquiring functionally mature human CMs, pharmaceutical developers have relied on animal CMs to screen and evaluate their candidate molecules for cardiotoxicity and arrhythmogenesis potential in preclinical studies. Despite extensive animal trials, there have been many cases of unsuccessful clinical trials due to the unseen cardiotoxicity in pre-clinical settings. Such failures underpin the inadequacy and fallibility of animal-based evaluations, and they are now increasingly considered as an obsolete platform for human application. hPSC-CMs provide considerable advantages for an alternative preclinical tool for providing better predictability of molecules in question prior to human clinical trials. Indeed, recent studies on the efficacy and toxicity of various drugs (Nifedipine, mitoTEMPO, Norepinephrine, etc.) have been carried out using CMs derived from hypertrophic cardiomyopathy (HCM), dilated cardiomyopathy (DCM), Barth syndrome (BTHS), Long QT syndrome (LQTS) 1, 2, 3, 8, catecholaminergic polymorphic ventricular tachycardia (CPVT) and arrhythmogenic right ventricular dysplasia (ARVC) patients, and their value as a cellular resource capable of coping with preclinical tests has been highlighted (Table 2). It is expected that the emergent hPSC-CMs based assays will continue to consolidate their presence in preclinical settings, which will lower the usage of animals and failure rate in clinical trials.
Conclusion
The outlook for hPSC-CMs application is promising: the human clinical trials for assessing safety and efficacy of hPSC-CMs for treating myocardial infarction are underway, a growing number of pharmaceutical companies have expressed their interests in endorsing hPSC-CMs for their preclinical testing, and pathogenic studies using heart disease patients’ iPSC-CMs are leading the way to discover more effective treatments. For a widespread application of hPSC-CMs though, the following should be addressed by investigators. Although hPSC-CMs differentiation could be performed in various ways, more often than not, the consistency and efficiency of differentiation vary when different hPSCs lines are used due to the distinct inherent genetic characteristics of each cell line. Development of highly efficient universal differentiation methods would make CMs readily available for applications at reduced costs. Various maturation techniques for hPSC-CMs have been effectual in their own merits, but characteristics of matured hPSC-CMs still fall short of that derived from adult tissue. This precludes their application in drug development and screening since the way immature CMs respond to molecules will be different to that of adult CMs. Interestingly, from cell therapy perspective, CMs of neonatal, and fetal origins have demonstrated better survival then mature CMs upon transplantation. Thus, optimum maturation also needs to be identified for different applications of hPSC-CMs. Nonetheless, the future prospect of hPSC-CMs is exciting, and looks certain to cause a paradigm shift in discovering novel drugs and therapies for heart diseases.
 XML Download
XML Download