

 PDF
PDF Citation
Citation Print
Print


Introduction
In the past decade, there have been a large number of researches on mesenchymal stem cells (MSCs), reporting their biological characteristics, experimental and clinical applications (1). MSCs are recognized by two important properties of extensive self-renewal ability and multi-lineage differentiation potential into mesenchymal and non-mesenchymal cell lineages (2). During the last few years isolations of adult mesenchymal stem cells from different sources have been reported. Bone marrow-derived MSCs (BM-MSCs) were first discovered by Friedenstein et al. (3), as an adherent fibroblast-like population, are still the most frequently investigated cell type and often identified as the gold standard. However, similar cell population have been also isolated from other tissues such as liver, lung, spleen (4), dental pulp (5), umbilical cord (6), peripheral blood (7), and adipose tissue (2).
On the other hand, although bone marrow has been considered as a main source for isolation of multipotent MSCs, because of the painful isolation process and low cell yield of BM-MSCs (8), adipose tissue has recently been attracting attention as a suitable and rich source of MSCs, giving multipotent cell population, named adipose tissue derived MSCs (AT-MSCs) (9). Unlike bone marrow, adipose tissue is abundantly accessible source of stem cell that can be collected in a large volume with minimal morbidity, which therefore reduces the time in culture required to generate a therapeutic cell dose (9). AT-MSCs were first extracted and described by Radble et al. (10), who collected rat adipose tissue sections through an open surgery. During the last decade, there has been an important attempt within the scientific community to focus on the characterization of MSCs were obtained from adipose tissue compared with those from bone marrow in different species (8, 11, 12). Even in those limited researches, there is disputation regarding in particular the growth properties of AT-MSCs versus BM-MSCs. While some studies have indicated, BM-MSCs possess a higher proliferation rate than AT-MSCs in culture media (13), the others have reported the higher significant comparative growth rate of AT-MSCs (14). Furthermore, differentiation potential as a main factor of isolated MSCs, beside of proliferation capacity and senescence of these cells are important aspect with regard to their application in cell therapy and tissue engineering (15).
MSCs derived from Guinea pig as an animal model which has several parallels with the human condition can be used in auditory, immune system, skin and joint disease studies. Anatomy of the Guinea pig ear is important for inner ear studies (16). Similar immune system to human, similar antigen-macrophage interaction and delayed cutaneous hypersensitivity reaction (17) initiation and osteoarthritis progression in Guinea pig which develops spontaneously with advancing age are similar to human (18). Therefore, the goals of the current study were to isolate, culture and compare Guinea pig BM-MSCs and AT-MSCs for their growth characterization, and proliferation capacity using growth curve analysis and population doubling time (PDT). Furthermore, we analyzed BM-MSCs and AT-MSCs differentiation potential to osteogenic and adipogenic cell lineages. Our observations in this study could provide some experimental evidences on choosing a suitable cell source for a particular therapeutic purpose in Guinea pig animal model.
Go to : 
Materials and Methods
Animals
In this experimental study, bone marrow and adipose tissue were collected from three adult male (4 months old) Guinea pigs weighing 400~450 g, which obtained from the Laboratory Animal Center of Shiraz University of Medical Science, Shiraz, Iran. All procedures are in accordance with animal guideline care of Ethical Committee of Shiraz University of Medical Sciences. All applicable institutional and national guidelines for the care and use of laboratory animals were followed. They were housed under standard conditions for a week prior to use.
AT-MSCs isolation
To establish the AT-MSCs culture, the animals were sacrificed by diethyl ether. Abdominal adipose tissue was isolated and collected in 15-ml sterile tubes. Under sterile conditions, the excised adipose tissue was rinsed with phosphate-buffered saline (PBS, Sigma-Aldrich), minced into small pieces and digested with collagenase type I (Sigma-Aldrich) at 37°C for 20 minutes. The samples were neutralized with an equal volume of Dulbecco Modified Eagle Medium (DMEM) supplemented with 10% fetal bovine serum (FBS, Gibco) and centrifuged at 1,200 rpm for 7 minutes. The pellet was then filtered through a 200 μm nylon mesh to remove undigested tissue. Adipose digested cells were suspended in DMEM supplemented with 10% FBS, 1% penicillin and streptomycin (Sigma-Aldrich) and 1% L-glutamine (Sigma-Aldrich) and cultured in 75-cm2 flasks. The cultures were incubated at 37°C with 5% CO2 and saturated humidity. The first culture media was changed after 24 h to remove non-adherent cells and the adherent cells were cultured and passaged to expand the MSCs population. The subsequent medium exchange was performed every 3~4 days till the cultures approximately 80~90% confluence. The adherent cells were washed twice with PBS and the cells were harvested using with 0.25% trypsin (Gibco) for 2~3 min and the enzyme was inactivated with same amount of culture media. AT-MSCs were passaged up to eight times. At each passage the cells were counted and analyzed for cellular growth. Spindle-shaped morphology of AT-MSCs was observed and analyzed using light microscopy at every passage.
BM-MSCs isolation
To establish the BM-MSCs culture, under sterile condition both femur and tibia from the guinea pigs were excised and carefully cleaned of adherent soft tissue. The ends of the bones were cut away and bone marrow was harvested by flushing with 10 ml syringe with DMEM supplemented media as explained above. After washing and centrifugation at 1,200 RPM for 5 minutes, cell pellet was collected and cultured in a 75-cm2 flask in a DMEM medium supplemented by 15% FBS and antibiotics. The cultures were incubated at 37°C in a 5% CO2 environment and saturated humidity. After incubation, the culture media was changed and passaged as described above for AT-MSC.
Reverse transcription-polymerase chain reaction (RT-PCR)
AT-MSCs and BM-MSCs were examined for expression of surface markers using RT-PCR. Total RNA of AT-MSCs and BM-MSCs at passage 3 of the Guinea pigs were extracted according to manufacturer’s instructions using RNX-Plus buffer (Cinnagen, Iran). Total RNA concentration was evaluated by spectrophotometer. After that, complementary DNA (cDNA) synthesis from DNA-free RNA (500 ng) samples was done by using Accu Power Cycle Script RT PreMix Kit (Bioneer, Korea) according to the manufacturer’s protocol. Before reverse transcription, the RNA samples were digested with DNase to remove contaminating genomic DNA. Specific primers were designed based on sequences corresponding to highly conserved regions of CD90, CD45, and CD34 in Guinea pig. The primer sequences used are summarized in Table 1.
Table 1
The primers used for RT-PCR analysis of mesenchymal cell confirmation of adipose tissue and bone marrow derived mesenchymal stem cells in Guinea pig
![]()
The RT-PCR amplification conditions for CD90, CD45, and CD34 were as followed: an initial amplification at 95°C for 5 min followed by 30 cycles of denaturation at 95°C for 30 sec, annealing at 64°C, 62°C, and 61°C for 30 sec, respectively and extension at 72°C for 30 sec, with a final polymerization at 72°C for 5 min. PCR Products were analyzed on 1.5% agarose gel electrophoresis and visualized by power UV light and photographed.
Cell counting and growth curves
In the present investigation, growth curves were plotted for each MSCs derived from abdominal adipose tissue and bone marrow tissues in order to better compare growth kinetics of the cells (19). For the assessment of growth characteristics, BM-MSCs and AT-MSCs at passage 2, 5 and 8 were seeded in a 24-well plates at a density of approximately 5×104 cells per well in triplicate. Cells were collected from each well 1~7 days after seeding and counted microscopically to produce cell growth curves. The curves were drawn using GraphPad Prism (Version 5.01; Graph-Pad software Inc., San Diego, CA, USA).
Calculation of PDT
To compare the in-vitro proliferation rate, the PDT value was determined for each studied cells. PDT was calculated using the formula PDT=T ln2/ln (Xe/Xb), in which T is the incubation time in hours, Xb represents the cell number at the beginning of the incubation time and Xe corresponds to the cell number at the end of incubation time.
Osteogenic and adipogenic differentiation assay
In order to compare the differentiation potential of BM-MSCs and AT-MSCs, cells of passage 3 were used and osteogenic and adipogenic differentiation were induced. For osteogenic differentiation BM-MSCs and AT-MSCs were seeded in 6-well plates. After the cells reaching 70% confluency, they were cultured for 3 weeks in osteogenic medium containing low glucose DMEM supplement with 100 nM dexamethasone (Sigma-Aldrich), 0.05 mM ascorbate-2-phosphate (Wako Chemicals, Richmond, VA, USA), 10 mM b-glycerophosphate (Sigma-Aldrich), 1% antibiotic/antimycotic and 10% FBS. The medium was replaced every 3 days. At day 21, the cells were fixed by 10% formalin solution (Sigma-Aldrich), and then stained using Alizarin Red (Sigma-Aldrich) to detect calcified extracellular matrix and osteogenic differentiation.
For adipogenic differentiation BM-MSCs and AT-MSCs were seeded in 6-well plates. When they reached 70% confluency, were induced to adipogenic differentiation with adipogenic induction medium containing DMEM low glucose, 10% FBS, 0.5 mM isobutyl-methylxanthine (Sigma-Aldrich), 10% FBS, 0.5 mM isobutyl-methylxanthine (Sigma-Aldrich), 1 μM dexamethasone, 10 μM insulin, 200 μM indomethacin (Sigma-Aldrich). The plates were maintained for three weeks and medium was replaced every 3~4 days. At the end of period, the cultures were fixed by 10% formalin solution for 10 minutes. Fixed cells were subjected to Oil Red O (Sigma-Aldrich), which specifically stains lipid droplets.
Statistical analysis
The mean and SE of counted cells in growth curve analysis were compared using one-way ANOVA (SPSS for Windows, version 11.5, SPSS Inc, Chicago, USA) and Tukey post-hoc test. Values of p≤0.05 were considered significant.
Go to :
Results
Phenotypic characterization of the cells
BM-MSCs and AT-MSCs were isolated from Guinea pig. After 48 hours, cells attached to the base of the tissue culture flask. The number of round-shaped cells gradually decreased and the growth rate of the fibroblastic cells gradually increased in culture media. Eight successive passages were done after reaching 70~80% confluency each time. Fibroblast-like cells were observed in all passages (Fig. 1).
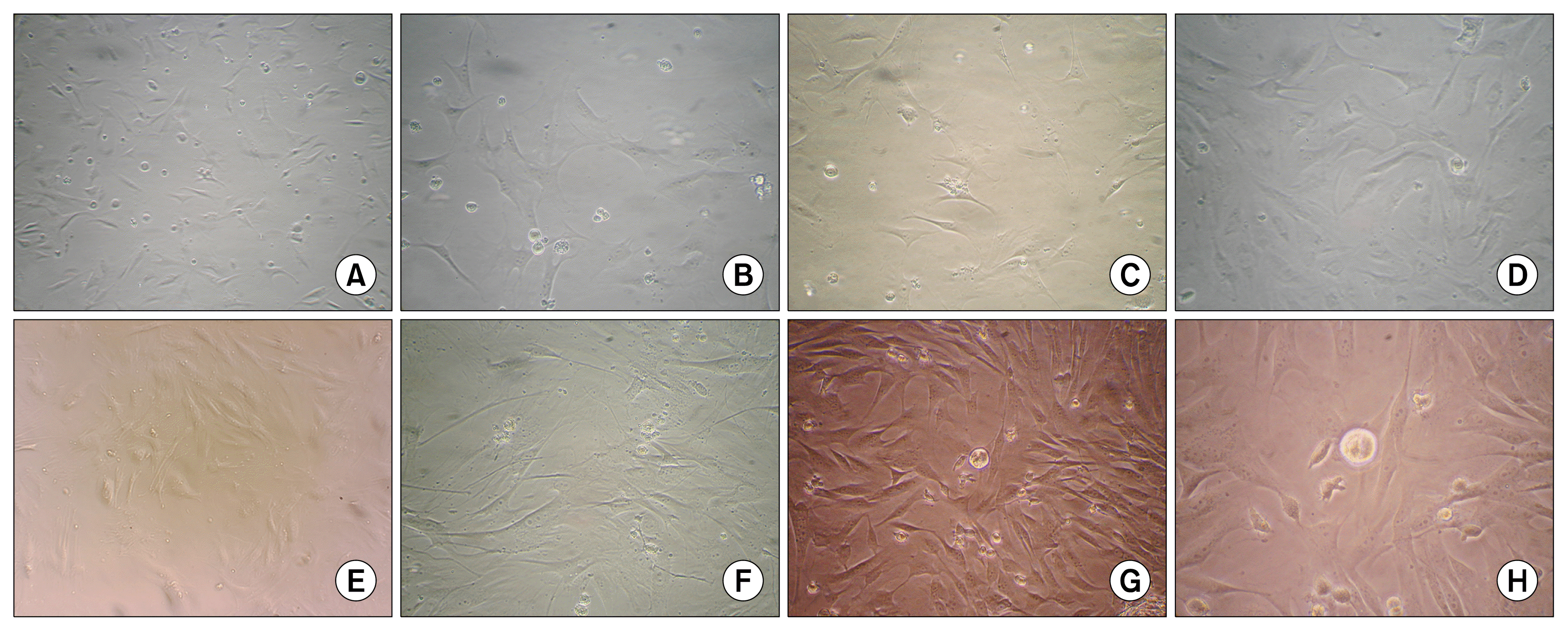 | Fig. 1Morphologic characteristics of adipose tissue-derived and bone marrow-derived mesenchymal stem cells (AT-MSCs and BM-MSCs, respectively) of Guinea pig. Most MSCs showed fibroblastic morphology regardless of the cell source. (A) Primary culture of AT-MSCs (×40), (B) Passage 2 of AT-MSCs (×100), (C) passage 5 of AT-MSCs (×100), (D) passage 8 of AT-MSCs (×100), (E) Primary culture of BM-MSCs (×100), (F) passage 2 of BM-MSCs (×100), (G) passage 5 of BM-MSCs (×100), and (H) passage 8 of BM-MSCs (×200).
|
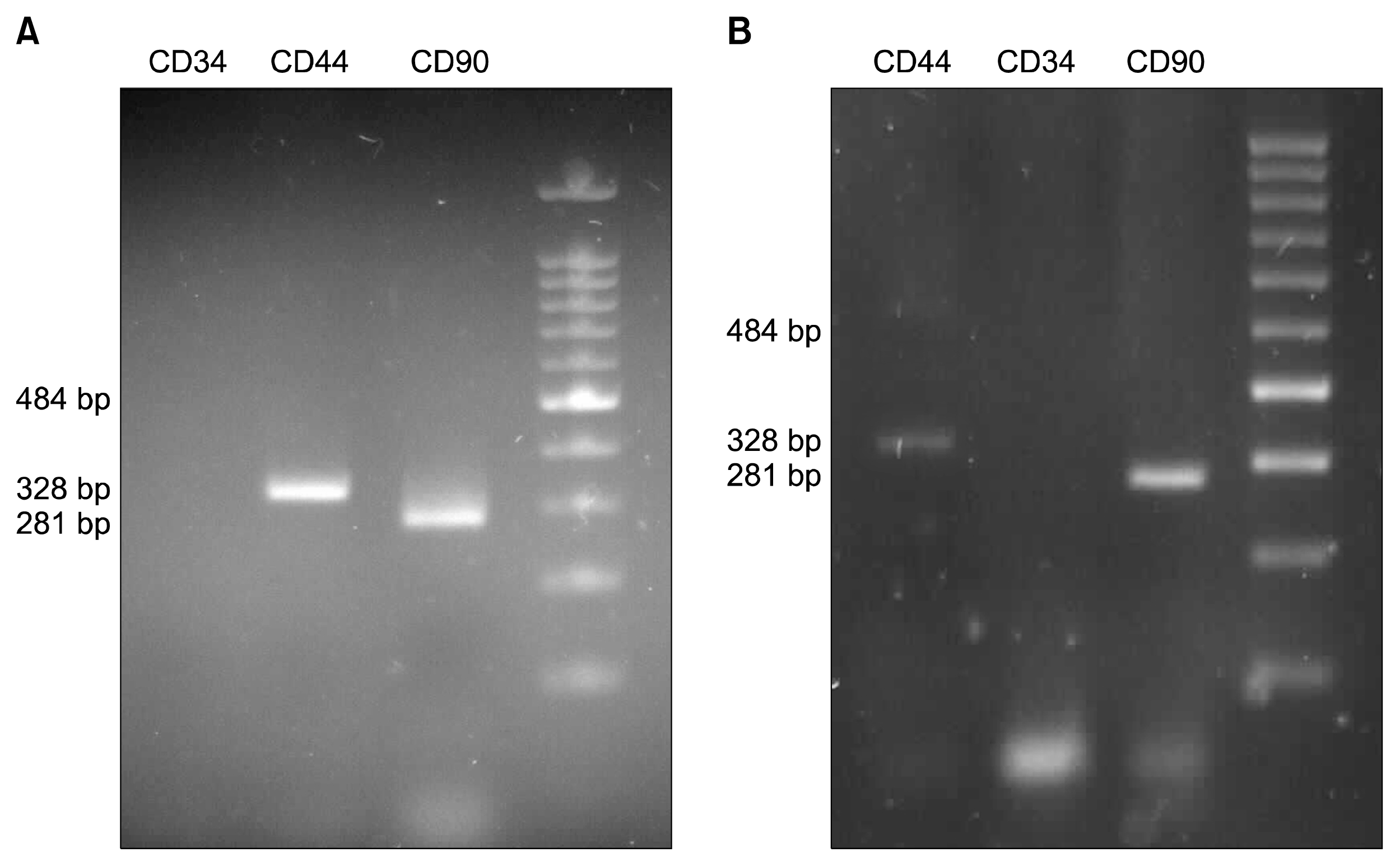
Cell surface markers of AT-MSCs and BM-MSCs
The expressions of cell surface markers were shown in AT-MSCs and BM-MSCs by RT-PCR analysis (Fig. 2). Both AT-MSCs and BM-MSCs were positive for MSC markers (CD44 and CD90) and negative for hematopoietic markers (CD34).
Growth Characteristics of the MSCs
According to our result the PDT of the passages 2, 5, and 8 of the AT-MSCs were 59.7 h, 64.2 h and 80.9 h, respectively. Also PDT was 62.9 h, 65.6 h and 91.4 h at the passages 2, 5, and 8 for BM-MSCs. Both AT-MSCs and BM-MSCs showed enough good proliferation rates in passages 2, 5, and 8, especially at passages 2 and 8. By comparing of growth curves, proliferation rate of AT-MSCs was more than BM-MSCs in passages 2 (Fig. 3A), 5 (Fig. 3B), and 8 (Fig. 3C; p<0.05). In this study the proliferation rate of AT-MSCs (Fig. 4A) and BM-MSCs (Fig. 4B) decreased gradually from passage 2 to passage 8.
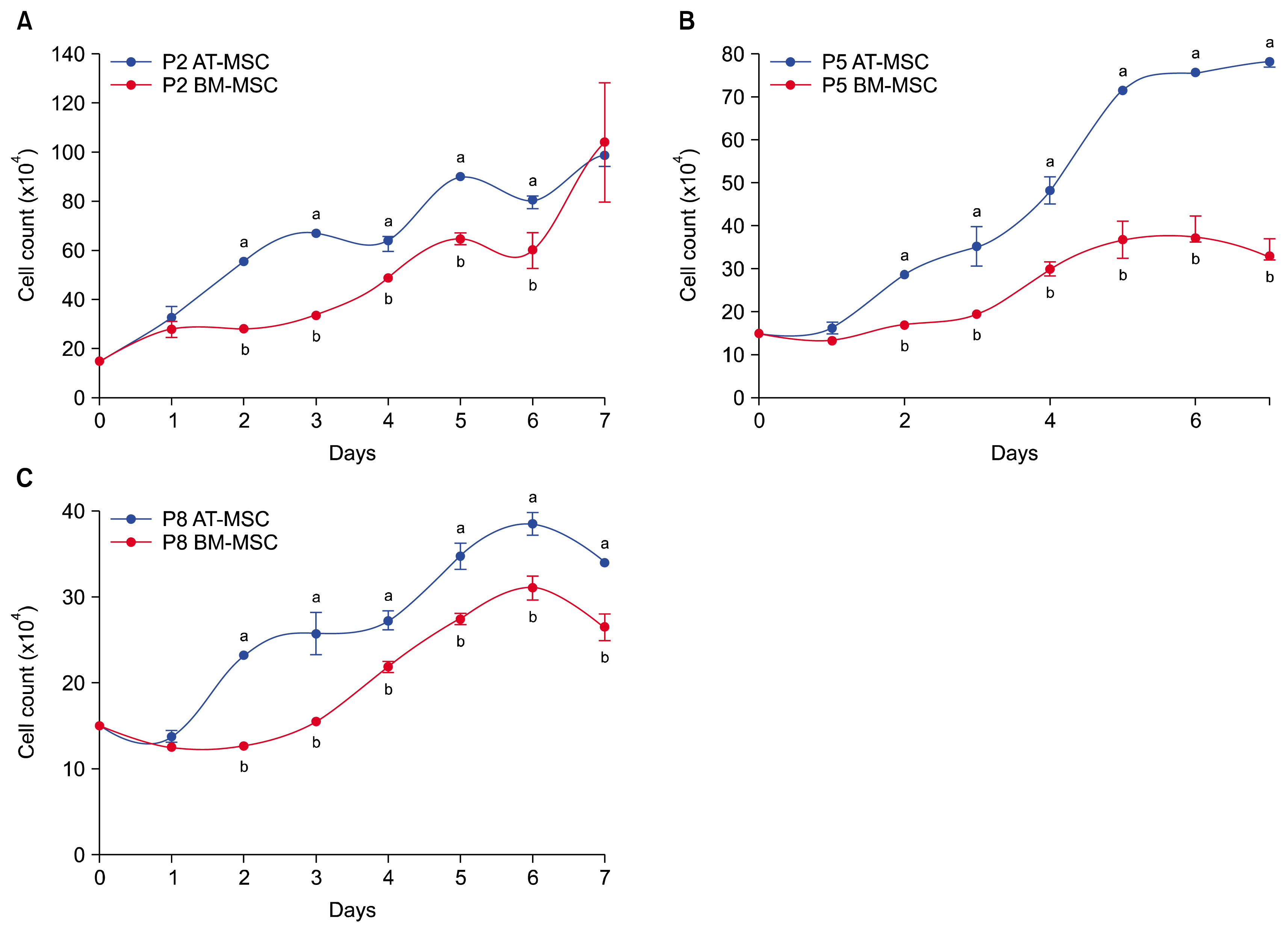 | Fig. 3Comparison of mean and standard error of cell counts between growth curves of guinea pig bone marrow-derived mesenchymal stem cells (BM-MSCs) and adipose tissue-derived mesenchymal stem cells (AT-MSCs) at passages 2, 5 and 8. Proliferation rate of AT-MSCs was more than BM-MSCs, (A) in passages 2, (B) in passages 5, and (C) in passages 8. a,bSuperscript letters show differences between the number of cells in each curve in the same day (p<0.05).
|
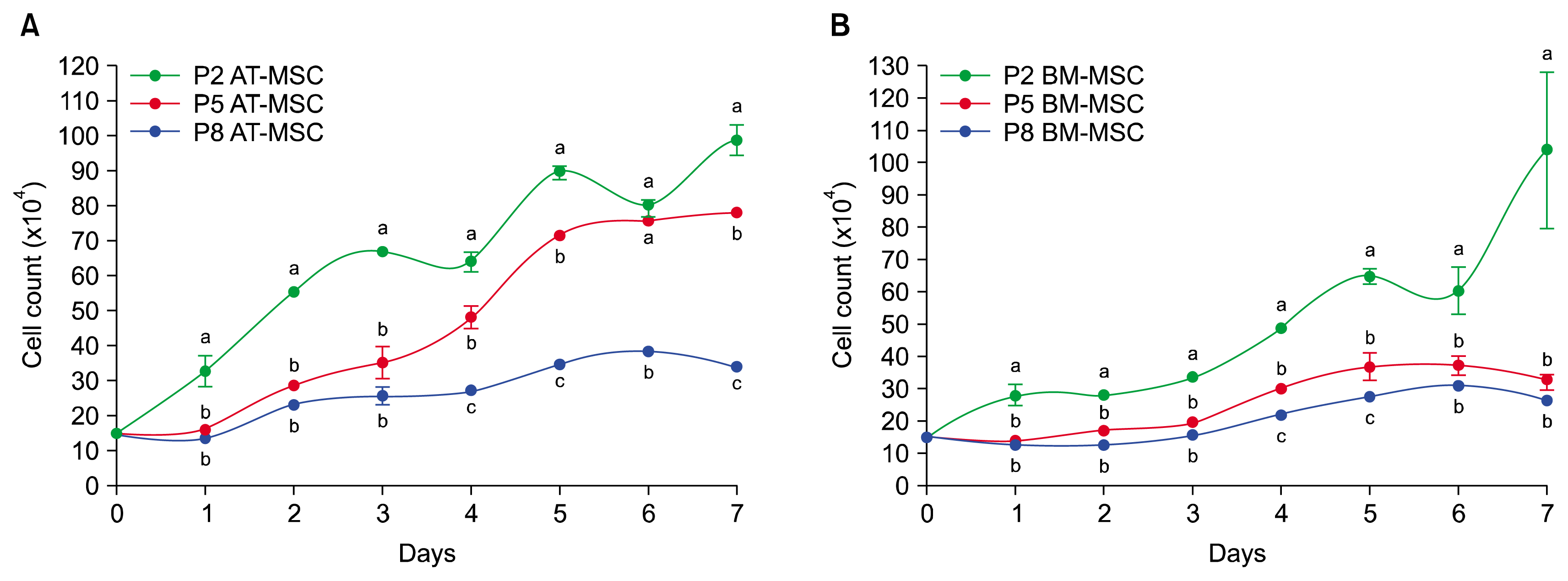 | Fig. 4Comparison of mean and standard error of cell counts in growth curves of guinea pig bone marrow-derived mesenchymal stem cells (BM-MSCs) and adipose tissue-derived mesenchymal stem cells (AT-MSCs) between passages 2, 5 and 8. The proliferation rate of AT-MSCs (A) and BM-MSCs (B) decreased gradually from passage 2 to passage 8. a,b,cSuperscript letters show differences between the number of cells in each curve in the same day (p<0.05).
|
Differentiation assay
Both types of MSCs successfully differentiated into adipocytes and osteoblasts lineages (Fig. 5). The negative control (non-induced) cells for each type of differentiation were negative for Alizarin Red and Oil Red O stains. Osteogenesis of BM-MSCs and AT-MSCs demonstrates mineralized matrix 21 days after induction. Adipogenesis of BM-MSCs and AT-MSCs as detected by the formation of lipid droplets stained with Oil Red O staining, 21 days after induction.

Go to :
Discussion
In addition to bone marrow, as a main source of MSCs, adipose tissue has been known as a source of MSCs (14). Moreover, abdominal adipose tissue is abundantly accessible and often applied for isolation of MSCs and tissue engineering (20). Therefore, some previous researches compared MSCs from adipose tissue with those from bone marrow but the results concerning comparative growth properties and differentiation potential of the cells remained as the subject of controversy (14, 21). Since, in the present research, Guinea pig AT-MSCs of abdominal regions as well as BM-MSCs were isolated and compared in terms of their in-vitro morphology, growth characteristics and differentiation capacity. Isolated Guinea pig AT-MSCs and BM-MSCs had plastic adherence capability and expanded in-vitro. These cells exhibited typical fibroblastic-like morphology, as one of MSCs recognition parameters, in all passages. To further characterize these cells, cell surface markers were examined by using RT-PCR. MSCs from both sources displayed, presence of mesenchymal cell surface markers (CD44 and CD90) and the lack of hematopoietic cell surface marker (CD34).
According to the growth curve analysis, growth and proliferation rate of AT-MSCs and BM-MSCs decreased gradually from passage 2 to passage 8. Consistent with our results, with increasing passage number of the human MSC, proliferation, growth rate and the number of cells in culture were decreased (22, 23). By comparing of growth curves, growth and proliferation rate of AT-MSCs was more than BM-MSCs, especially in passage 8 in Guinea pigs. Also, in the rat, AT-MSCs possessed a higher expansion rate than BM-MSCs (15). Moreover, in human, AT-MSCs have also been shown to have higher proliferation capacities than BM-MSC (24). The PDT of Guinea pig BM-MSCs in analyzed passages 2, 5, and 8 were more than AT-MSCs. Moreover, in the present study it was observed that BM-MSCs showed a bit increase in size versus AT-MSCs. The same as our findings, Peng et al. (14) described PDT of 45.2 h for AT-MSC and 61.2 h for BM-MSC in human. Moreover, they revealed that the human BM-MSC were morphologically larger than AT-MSC (14). Therefore, BM-MSCs density can be rapidly increased in culture and reached saturation density earlier than AT- MSCs. Hyper-density of cell population displayed a negative role against cell proliferation because of contact inhibition and reciprocal effects of cells on each other (14, 25). AT-MSCs seemed to have less percentage of senescent cells and more proliferation rate during the passages in culture media than the BM-MSCs (15). It should however be noted that the origin of the cells, the cultivation conditions and various medium supplements may have an effect on PDT of MSCs (26). Wagner et al. (27) analyzed global gene expression profiles of human MSCs isolated from adipose tissue versus bone marrow; who demonstrated that higher proliferation capacity in AT-MSCs could be due to significant differences in the gene expression patterns and higher expressed of cell division cycle associated 8 (CDCA8), and cyclin B2 (CCNB2) gens in AT-MSC than in BM-MSC.
In addition to proliferation potential, differentiation capacity has been considered as an important quality of MSCs. In this experimental study, to assess the differentiation potential of AT-MSCs and BM-MSCs, passage 3 cells were used. Differentiation is a process which dramatically changes a cell in size, morphology, membrane potential and metabolic activity caused by highly-controlled modifications in gene expression (28). According to Alizarin Red staining for mineralized matrix in osteogenic differentiation and Oil Red staining for lipid droplets in osteogenic differentiation, AT-MSCs and BM-MSCs differentiated into adipose and bone cell lineage similar to previous studies (29, 30). Although various differentiation ability in MSCs that reported in some previous research could be due to, different genes expressed in MSCs from different ontogenetic sources and culture conditions (27).
Go to :
Conclusion
Despite the importance of Guinea pig as an experimental animal model for many conditions, the characterization of Guinea pig MSCs is still limited. In this study, we have successfully isolated and compared MSCs population from Guinea pig adipose and marrow tissue. AT-MSCs and BM-MSCs sources are highly accessible, mechanically stable and readily expanded in culture flask. These cells had enough good proliferation capacity and display high potential toward adipogenic and osteogenic differentiation, while proliferation rate and PDT of AT-MSCs were better than the same cell derived from BM-MSCs. The resent study results showed that although adipose tissue may prove to be a more efficient source of MSC and there were some differences between BM-MSCs and AT-MSCs, but these cell sources can be considered as an appropriate candidate for use in preclinical studies and regenerative medicine applications.
Go to :
 XML Download
XML Download