

 PDF
PDF Citation
Citation Print
Print


Introduction
Most human somatic cells do not express telomerase, in other word, telomere sequences are lost during replication and limiting their proliferative capacity (1). In contrast, telomerase is very active in human cancer cells and a small number of normal proliferating cell types such as germ line and progenitor cells (2). The level of telomerase is low in the majority of human stem cells, such as neurons, skin, intestinal crypt, mammary and liver epithelial, pancreas, adrenal cortex, kidney, and mesenchymal stem cells (MSCs) (3). Telomerase activity correlates with the expression pattern of the telomerase reverse transcriptase catalytic subunit; hTERT gene which specifically elongates telomeres. A critical length of telomere repeats is required to avoid replicative senescence or cell death (3). Human mesenchymal stem cells (hMSCs) have attracted much attention because of their multi-differentiation and self renewal properties and their possible use of cell and gene therapies (2). Despite the mentioned advantages, aged MSCs have a limited life span, initial cell viability and cell yield in culture, gradually lose their differentiation potential and undergo senescence-associated growth arrest under current culture conditions, a phenomenon termed replicative senescence, and cells will be senescent or die after several passages (4, 5), which restricts the clinical applications such as regenerative medicine or gene therapy. Furthermore, proliferation and population doubling time in aged MSCs are slowed compared with the young MSCs ones (6, 7). So it is crucial to maintain the proliferation and differentiation capacity of MSCs. It seems aging is the result of the accumulation of oxidative damage caused by free radicals generated as by-products during normal metabolism (8). The use of antioxidants to prevent cellular aging is important. L-carnitine (LC) or γ-trimethylamino-β-hydroxybutric acid is a small water-soluble quaternary amine that is present in different concentrations in human serum and tissues where they vary in relation to different factors such as environmental changes, diets, hormones, sex and age (9, 10). Approximately half of the LC in the human body is supplied by biosynthesis and the rest of the LC can be supplied by food (11). Many physiological effects of LC have been shown. Deyhim et al. demonstrated that LC as an additive improves the platelet metabolism in maintaining the quality of stored platelet concentrate for platelet transfusion therapy (12). Hollister et al. reported LC leads to a modest decrease in the rate of progression of Alzheimer’s disease (13). Bonavita et al. determined intensive acetyl-L-carnitine (ALC), an acetyl derivative of LC, treatment could influence a significant improvement of the main mental parameters of the senile brain, without any of side effects (14). Kobayashi et al. indicated that ALC increases synaptic neurotransmission in the brain and consequently improves learning capacity in aging rats. Musicco et al. reported that ALC reverses the age-related alterations of 10 mitochondrial proteins relative to mitochondrial cristae morphology, to the oxidative phosphorylation and antioxidant systems (15). Recently, consideration has been focused on the antioxidant effects of LC but the specific mechanism is still unclear (9, 16). It has been speculated that LC protects cells from the oxidative damages, by inhibiting free-radical propagation and by contributing to repair of oxidized membrane phospho-lipids (17, 18). Although many studies show the neurological effects of LC and ALC in the aging brain, but its accurate mechanisms are not clear (11). This study aimed to investigate the effects of LC in different doses on the hTERT gene expression, telomere length and on the cell proliferation rates of the aged human adipose tissue-derived MSCs.
Go to : 
Materials and Methods
Subjects and isolation of adipose tissue-derived hMSCs
The adipose tissue samples were collected from the six normal women (45~58 years old) after undergoing liposuction surgery performed at the Imam Reza Hospital, Tabriz University of medical Sciences, Tabriz, Iran. This stage of the project was approved by the ethics committee of the hospital, as well as obtaining informed consents from all subjects. The prepared samples were put in the vials containing 5 ml complete mediums (Dulbecco’s modified Eagle’s medium (DMEM) containing 10% fetal bovine serum (FBS, Gibco, UK), 100 U/ml penicillin, and 100 μg/ml streptomycin. All of the samples were immediately transferred to the lab for further procedures. To isolate of adipose tissue-derived hMSCs, the tissues were washed extensively with equal volumes of the phosphate-buffered saline (PBS) for three times and digested enzymatically for 30 min at 37°C using 0.075% (w/v) collagenase type I (Invitrogen, UK). Enzyme activity was neutralized with DMEM containing 10% (v/v) FBS, and centrifuged at 800×g for 5 min to obtain a pellet. The pellet was washed once with PBS and then re-suspended in complete medium and added into a 24 well plate followed by incubation at 37°C with 5% CO2 in a humidified environment for three days. The medium was changed every 3 days in each of the experiment. When the cells became confluent they were trypsinized with 0.25% trypsin (Gibco) and 1 mM EDTA (Invitrogen) and sub-cultured.
Cell proliferation assay
The tetrazolium-based colorimetric assay (MTT test) measures the mitochondrial activity of the cell culture, which reflects the number of viable cells. When each of early-passage MSCs cells grew to about 70% to 80% confluency, they were washed by PBS and trypsinized with Trypsin-EDTA. The cells were counted with hemocytometer and 4×103 cells were seeded in each well of a 96-well plate with a total volume of 200 μl of complete medium and incubated for 24 h at 37°C in a humidified environment with 5% CO2 to grow the cells in a monolayer. Then, L-carnitine was added to the wells at final concentrations of 0.1, 0.2 and 0.4 mM and incubated under the same culture conditions for 24, 48 and 72 hours. The control wells were prepared by addition of corresponding mediums. After the mentioned times, treatment medium was aspirated and then, the cells were incubated with stock MTT dye solution (5 mg/mL) for 4 h at 37°C. Following incubation time, the supernatant was removed and 100 μl of dimethyl sulfoxide (DMSO) was added. The optical density of each well was measured by ELISA Reader at a wavelength of 570 nm.
Phenotypical characterization of adipose tissue-derived hMSCs by Flow-cytometry
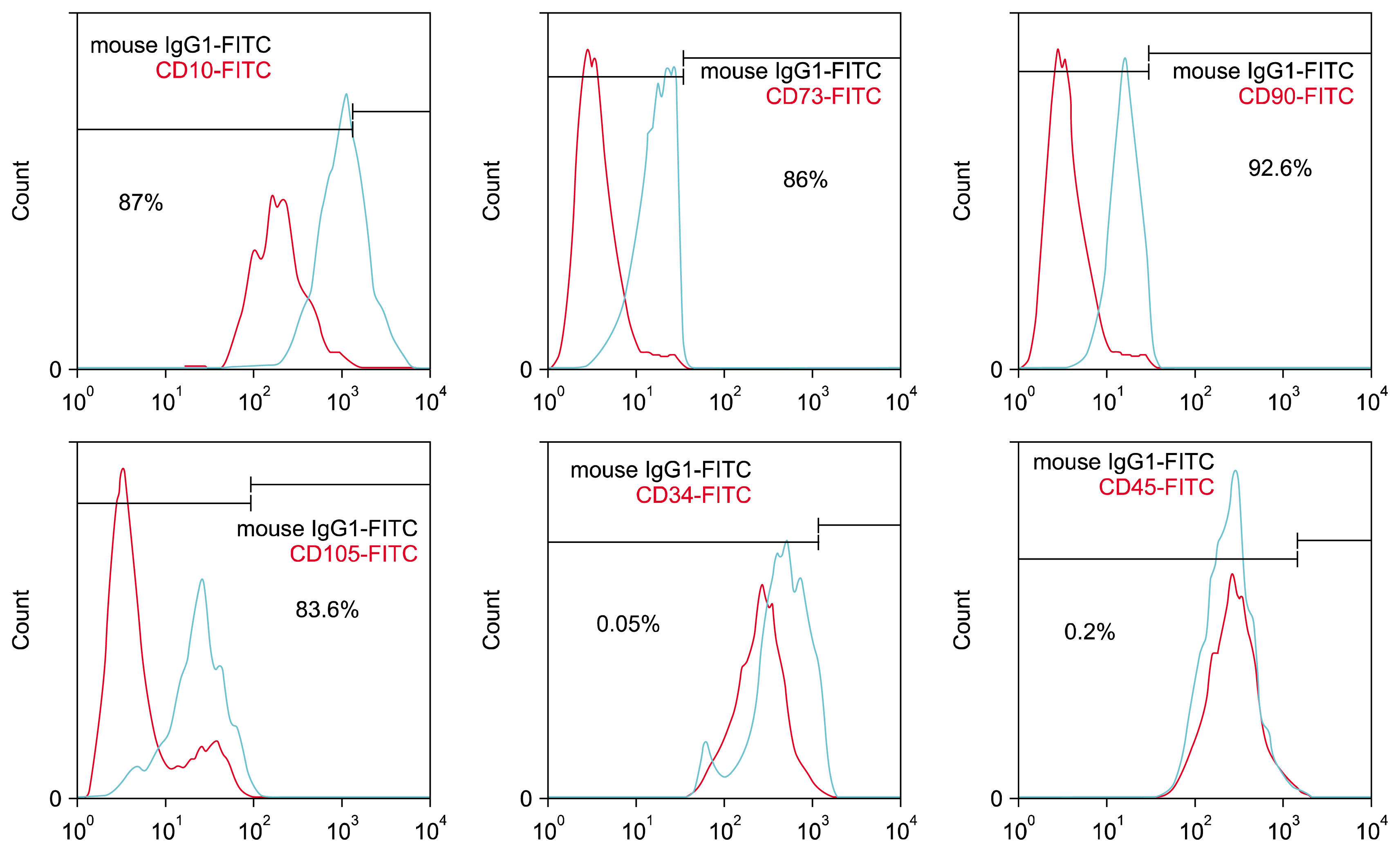
To analyze cell surface markers, approximately 106 cells of adipose tissue-derived hMSCs were harvested, washed in PBS and incubated with an appropriate amount (1 μg/106 cells) of fluorescein isothiocyanate (FITC)-conjugated mouse primary CD10, CD73, CD90, CD105, CD34 and CD45 antibodies (all from BD PharMingen) in PBS buffer supplemented with 1% FBS for 30 min on ice. These cells were washed in PBS and fixed in 10% formalin prepared in PBS containing 2% FBS. FACS (Fluorescence Activated Cell Sorter) analysis was also done using a FACScan (Becton Dickinson) and data were analyzed with FlowJo software.
Osteogenic differentiation and alizarin red staining
For osteogenic differentiation of adipose tissue-derived hMSCs, cells at passage 4 were cultured at an initial density of 50×103 cells in each well of 6-well plates to reach confluence before differentiation. To induce mineralization, cells were incubated in osteogenic medium containing 10% FBS, 10 nM dexamethasone, 100 U/mL penicillin, 100 μg/mL streptomycin, 10 mM beta-glycerophosphate, and 0.05 mM L-ascorbic acid-2-phosphate. Control cultures were fed only complete medium. After 21 days of culture, osteogenic differentiation of stem cells was confirmed by positive alizarin red S staining. The cells in 6-well plate were washed with PBS and fixed in 2% (v/v) formaldehyde. After 15~20 min, Alizarin red S 2% (pH 4.1) was added to each well. The plates were incubated at room temperature for 20 min and washed four times with PBS and then shaken for 5 min.
Adipogenic differentiation and oil red staining
To promote adipogenesis, subconfluent cells were incubated in adipogenic medium containing 0.5 mM 1-methyl-3 isobutylxanthine, 1 μM dexamethasone, 10 μg/ml insulin, and 200 μM indomethacin. After 21 days of culture to visualize lipid droplets, formalin-fixed cells were washed in 50% isopropanol and stained with Oil Red-O.
RNA extraction and cDNA synthesis
Approximately a 15×104 initial density of adipose tissue-derived hMSCs/well was seeded in 6-well plates and fed with complete medium as described. Three days after seeding, LC was added to the wells at final concentrations of 0.1, 0.2 and 0.4 mM for up to 48 h at 37°C in 5% CO2. The cells were collected and suspended in Trizol reagent (Invitrogen) in a clean RNase-free tube and were incubated for 5 min at 25°C. After incubation, 200 μl chloroform was added, shaken vigorously for 15 seconds, and incubated for another 5 min. The mixture was centrifuged at 12000 g for 15 min. The upper aqueous phase was carefully transferred to a clean 1.5 ml RNase-free tube. The total RNA was precipitated by adding 500 μl isopropanol and incubating for 15 min at 25°C. The pellet including total RNA was washed using 75% ethanol and centrifuged at 7500 g for 8 min. After drying the ethanol, the RNA pellet was dissolved in DEPC-treated water. Reverse transcription was carried out using the RevertAid™ first strand cDNA synthesis kit (Fermentas, Lithuania). For every reaction set, one sample was also prepared without the reverse transcriptase enzyme to provide a negative control in the subsequent PCR.
Real-time PCR amplification for hTERT expression
All PCR reactions were performed using the Corbett Rotor-Gene™ 6000 HRM (Corbett Research, Australia) in a total volume of 20 μl containing power SYBR Green master mix (TaKaRa), 0.1 μM of each primers, cDNA (40 ng/μl) and H2O. The specific primers used are listed in Table 1. Both β-actin and hTERT amplification were done in triplicate for each sample. The thermal cycling conditions were as follow: 2 min at 94°C, followed by 40 cycles, each at 94°C for 15 s and at 63°C for 1 min for both β-actin and hTERT. The data were normalized against the CT (threshold cycle) of the β-actin control. The CT values were calculated in relation to β-actin CT values by the 2−ΔΔCT method, in which ΔCt was the difference between the Ct value of hTERT and the Ct value of β-actin (19).
DNA isolation
Cells were plated in 6-well culture plates at the density of 15×104 cells/well and fed with complete medium as described. After 3 days, these cells fed with various concentrations (0.1, 0.2 and 0.4 mM) of LC up to 48 h. Genomic DNA was isolated from the cells using genomic DNA purification kit (Fermentas) and stored at 4°C or −20°C.
Oligomeres, telomere and single copy gene standard curves
Oligomer sequences for absolute telomere length (aTL) measurement are shown in Table 2. A standard curve is created by dilution of known quantities of a synthesized 84-mer oligonucleotide containing TTAGGG repeats 14 times with 84 bp in length and a molecular weight of 26667.2 Single copy gene (SCG) is used as a control for determining genome copies per sample. In this regard, 36B4 gene which encodes the acidic ribosomal phospho-protein (P0) was routinely used. SCG amplification is crucial for the accuracy and reliability of the results generated in the aTL assay. Genome copy number per reaction is calculated as previously described by O’Callaghan and Fenech (30).
Table 2
Oligomers used for aTL assay
![]()
Real-time PCR amplification for aTL measurement
All samples were run in a total volume of 20 μL containing Power SYBR Green master mix (2x), 0.1 μM of each primers and DNA (20 ng/μl). Cycling conditions (for both telomere and 36B4 amplicons) were as follows: 5 min at 95°C, followed by 40 cycles of 95°C for 15 sec, 64°C for 1 min, followed by a melt curve analysis.
Processing and analyzing data for measuring the telomere length
Real-time PCR data were analyzed as kb/reaction for telomere and genome copies/reaction for SCG. The kb/reaction value was then used to calculate total telomere length in kb per human diploid genome. The telomere kb per reaction value is divided by diploid genome copy number to give a total telomeric length in kb per human diploid genome. The obtained value can be further used to give a length per telomere by dividing by 92 which is the total number of telomeres on 23 pairs of chromosomes found in normal human cells.
Statistical analysis
The results were analyzed using the software programs GraphPad Prism 6.01. One-way and two-way ANOVA tests were used to determine the significant difference among groups. Statistical significance was determined at p<0.05.
Go to :
Results
Phenotypical characterization of adipose tissue-derived hMSCs
Adipose tissue-derived hMSCs had the capacity for attaching the culture flasks, and displayed as a fibroblast-like during the initial days of incubation. After 3 passages, red blood cells were seldom seen by light microscopy and the cells gradually grew into small colonies. Flow-cytometric analysis indicated that the cultured cells had high levels of expression of CD10 (87%), CD73 (86%), CD90 (92.6%) and CD105 (83.6) and also low levels of expression for CD34 (0.05%), and CD45 (0.2%) (Fig. 1).
Two lineage differentiation protocols
The osteogenic differentiation was evident in alizarin red staining. After mentioned staining, redness of the nodular aggregates indicating that calcium deposition has occurred as a result of the osteogenic treatment. Positive adipogenic differentiation was confirmed by oil red staining. Treated hMSCs with adipogenic differentiation media stained positive with oil red O staining. These results confirmed that the isolated cells have been adipose tissue-derived hMSCs (Fig. 2).
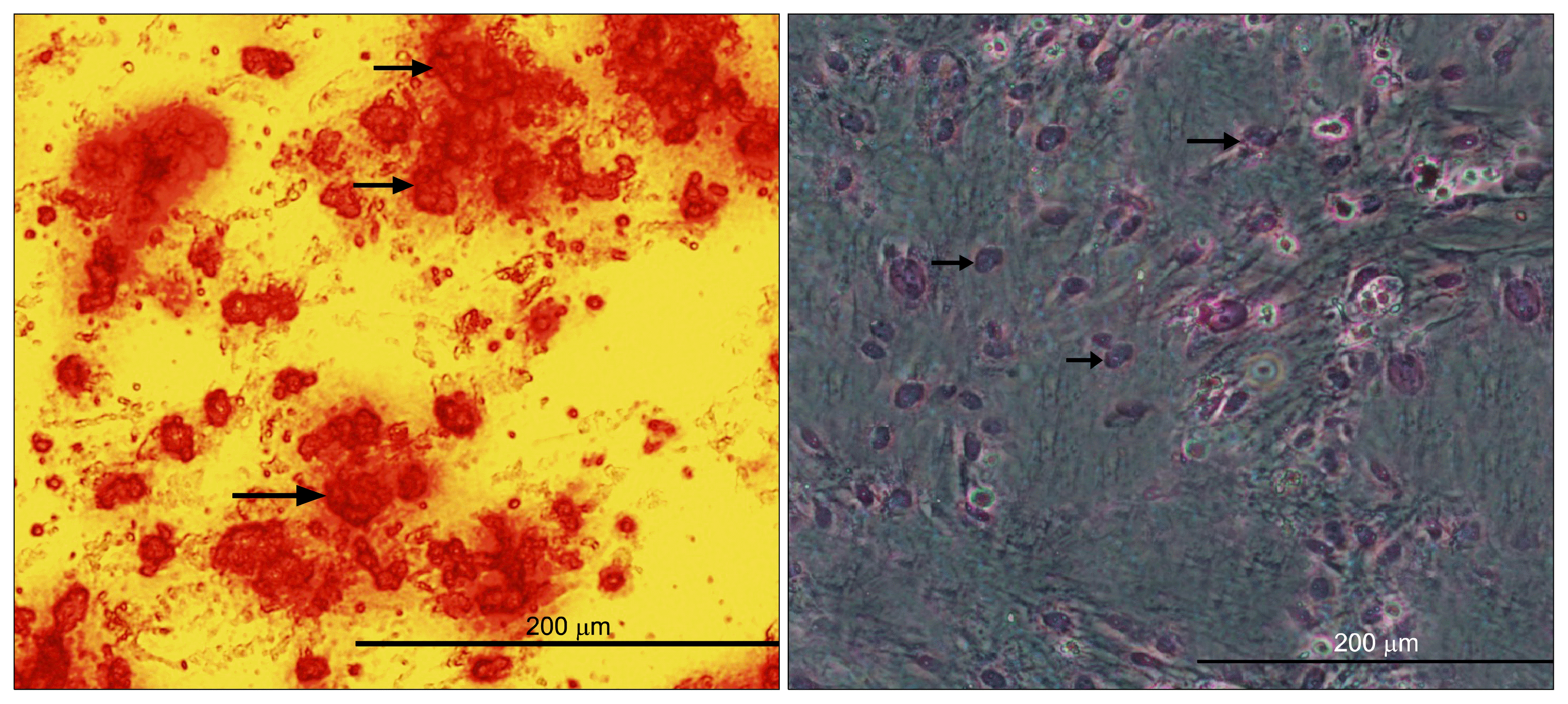 | Fig. 2Two lineage differentiation of ADSCs. (Left) Osteogenic differentiation and cell aggregates (were stained with alizarinred staining). Arrows show some of the mineralized cell aggregates (bar=200 μm). (Right) Differentiation into adipose cells. Arrows show lipid vacuoles generated after adipose differentiation (bar=200 μm).
|
The effect of LC on the adipose tissue-derived hMSCs proliferation
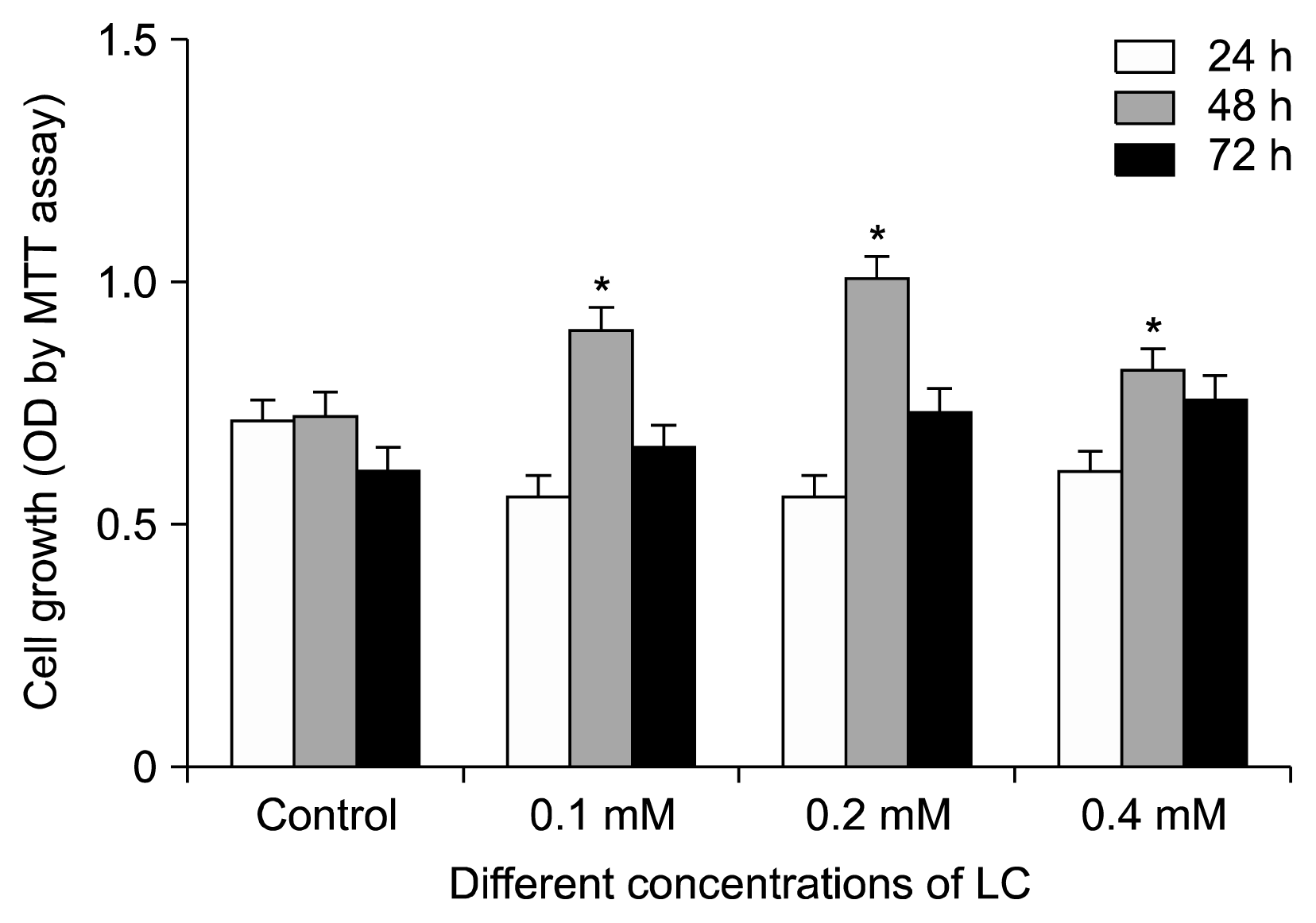
Adipose tissue-derived hMSCs were treated with different concentrations of the LC for 24, 48 and 72 hours and cell proliferation was examined by MTT assay. As shown in Fig. 3, there were significant differences in adipose tissue-derived hMSCs proliferation between the treated groups with 0.1, 0.2 and 0.4 mM of LC and the control group (p<0.05), especially after 48 hours. LC had no significant effect on the proliferation of treated cells compared to the control, after 24 and 72 hours. Therefore, the most suitable time for LC treatment of the adipose tissue-derived hMSCs seemed to be in the 48 hours of incubation.
The effect of LC on hTERT gene expression
Real-time PCR for the detection of hTERT expression in adipose tissue-derived hMSCs was carried out after 48 hours of culture in different concentrations of LC. The results showed that hTERT mRNA increased in the presence of 0.1 (44 fold), 0.2 (89.57 fold) and 0.4 (1.31 fold) mM LC compared to the control group. Also, as shown in Fig. 4, 0.2 mM LC remarkably increased the expression of hTERT after 48 hours of incubation (p<0.0001).
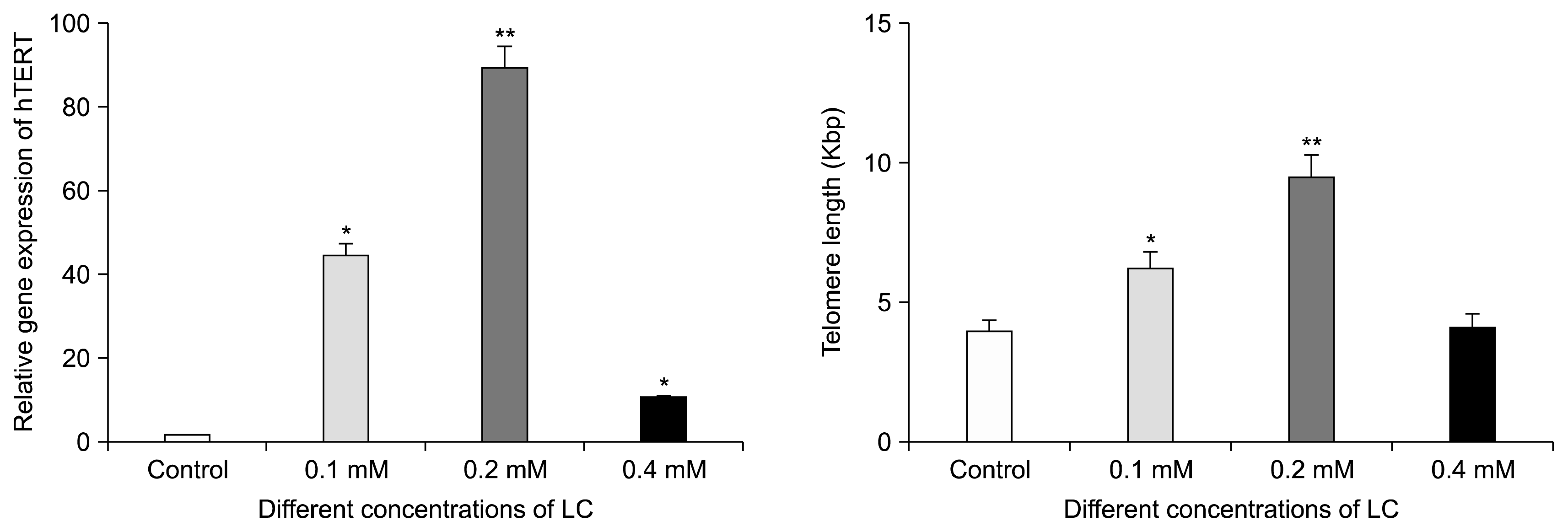 | Fig. 4Relative hTERT gene expression levels of adipose tissue-derived hMSCs in the presence of different concentration of LC for 48 hours of incubation (*p<0.05 and **p<0.0001 compared with the control group). (Right) Absolute telomere length measurement of tissue-derived hMSCs in the presence of different concentration of LC for 48 hours of incubation (*p<0.05 and **p<0.001 compared with the control group).
|
The effect of LC on telomere length
Real-time PCR was carried out after 48 h treatment of adipose tissue-derived hMSCs with 0.1, 0.2 and 0.4 mM LC. As shown in Fig. 4, telomere length increased significantly in the presence of 0.1 (6.19 Kbp) (p<0.05) and 0.2 (9.49 Kbp) (p<0.001) mM LC compared to the control group (3.89 Kbp).
Go to :
Discussion
Some studies claim that the hMSCs transplants from the aged donors are less effective (20). It has been reported that the aged hMSCs show a decline in differentiation potential as well as in proliferation rate (21). As reported by Stolzing et al., MSCs isolated from 56 week-old Wistar rats showed that the increased levels of apoptosis and reduced proliferation than that isolated from 8~12 week-old rats (22). Some molecular events such as changes in the telomere length and TERT expression are associated with the cell senescence. There is also a correlation between the proliferative capacity of the hMSCs and telomere length, both in culture and in the aged donor cells (23). This correlation indicated that the erosion of telomeres at each cell division of the hMSCs occurring until the telomeres reach to about 10 kb and consequently leading cell apoptosis (24). Some studies reported that the lower levels of the telomerase activity have been found in adult stem cells derived of the hematopoietic system (25). It is noticeable that there are a lot of reports indicated that no telomerase activity could be detectable in most of the normal somatic cells and even hMSCs (2, 25). In this regard, finding the ways to reduce the replicative senescence has definitely attractive in cell transplantation approach. According to the data reported by Böcker et al. the hTERT over-expression could prevent the hMSCs senescence and the cells showed significantly higher and unlimited proliferation capacities (5). Unlike the previous study, Britt-Compton et al. observed that hTERT over-expression alone could not prevent senescence despite preserving telomere length (26). The mechanisms by which hTERT over-expression may cause proliferation of stem cells, are still not clear. Other factors involved in the aging process are oxidative stress-mediated cellular damage via reactive oxygen species (ROS) (27). Recently, Liang et al. reported that aging increases the susceptibility of MSCs to ROS and impairs their therapeutic potency (20). Britt-Compton et al. indicated that more intense ROS-generating stressors can induce senescence via generation of telomere-independent DNA damage (26). On the other hands, it has been reported that antioxidant as an indicator of inhibiting the oxidative stress (28). Of all the antioxidants, exogenous / endogenous compounds, LC works as an endogenous with no adverse effects (29). Based on these effects of LC, we tried to plan a project to use LC to improve the lifespan of hMSCs. In this study, we found that the 0.2 mM of LC had the most effect on the cell proliferation in the time-dependent manner. The hTERT gene expression and telomere length significantly increased in LC-treated adipose tissue-derived hMSCs, as compared with untreated cells.
Go to :
Conclusion
This study showed for the first time that LC at the concentration of 0.2 mM could be used as a good candidate for extending the replicative life-spans of aged MSCs. Nevertheless, further in vitro and in vivo investigations need to be carried out to examine LC on aged adipose tissue-derived hMSCs and the conditions have to be optimized and established before cell therapy.
Go to :
 XML Download
XML Download