

 PDF
PDF Citation
Citation Print
Print


Introduction
Direct conversion is a process by which somatic cells are reprogrammed into tissue-specific cells, such as induced neural (iN) cells, spinal motor neurons, and dopaminergic (DA) neurons, using a combination of tissue-related factors and mitogens (1–3). Recent studies have described the direct conversion of somatic cells into induced neural precursor cells (iNPCs) and induced dopaminergic neurons (iDA) (4–6); however, the conversion efficiency to iDA in these studies was low. Caiazzo et al. (2) and Pfisterer et al. (3) showed that mouse and human fibroblasts can be directly converted to DA neurons using transcription factors, such as Nurr1, Ascl1, Lmx1a and Foxa2. These studies did not supplement culture media with fibroblast growth factor 8 (FGF8), but other research groups [e.g., Kim (7) and Liu (8)] have reported successful production of iDA neurons by adding FGF8 to media.
FGF8 has been implicated in early vertebrate brain patterning, and inhibition of its expression is associated with absence of the entire mesencephalic and cerebellar primordia and early embryonic death (9, 10). FGF8 is capable of binding to all four FGF receptors, with different affinities (11). When the growth factor binds to its receptor, phosphorylation of extracellular signal-regulated kinase 1/2 (ERK1/2) usually occurs and in turn activates the RAS-MAPK intracellular pathway (12, 13). Several neurotrophic factors, including sonic hedgehog (shh) and FGF8, are critical to the specification and differentiation of midbrain DA neuron development (14). Additionally, FGF8 immunoreactivity levels in the substantia nigra have been found to be significantly lower among Parkinson’s disease (PD) patients than in age-matched controls (15), although there is no evidence that this is a primary event in PD. It is therefore reasonable to assume that FGF8 may have dual roles in survival of iNPCs and maturation of iNPC-derived DA neurons.
Kim et al. (7) suggested that FGF8 is critical to promote iDA neuron generation from fibroblasts; however, their process directly reprogrammed fibroblasts into iDA neurons without passing through the NPC stage. Thus, we would like to evaluate whether FGF8 is in fact essential for iNPC conversion process and DA neuron generations passing through the NPC stage. To address this, we investigated the generation of iNPCs and DA neurons in the presence or absence of FGF8. Here, we demonstrate that early and continuous exposure of FGF8 to fibroblasts and iNPCs promotes maturation of iNPC-derived DA neurons.
Materials and Methods
Fibroblast isolation
Animals were housed and treated according to the Institutional Animal Care and Use Committee (IACUC) guidelines of Hanyang University. Embryonic fibroblasts (REF) from Sprague-Dawley rats (DaeHan BioLink, Seoul, Korea) were isolated from embryonic day 14 embryos after removing the head, spinal cord, and all internal organs, including the gonads. Fibroblasts were cultured in high-glucose Dulbecco’s modified Eagle’s medium (DMEM; Invitrogen, Carlsbad, CA, USA) supplemented with 10% fetal bovine serum (FBS; Invitrogen), 0.1 mM nonessential amino acids (Invitrogen), 0.1 mM β-mercaptoethanol (Sigma-Aldrich, St. Louis, MO, USA), 2 mM L-glutamine (Sigma-Aldrich), and 1% penicillin-streptomycin (Invitrogen).
Retroviral production
Retroviral constructs for Brn2 (B), Ascl1 (A), Myt1l (M), Bcl-xL (X), Nurr1 (N), and Foxa2 (F) were constructed by engineering the appropriate DNA fragments into the retroviral vector, pCL (16). The retroviral vectors were transfected into 293GPG packaging cells using Lipofectamine 2000 (Invitrogen). Supernatants containing viral particles were harvested 72 hours after transfection.
iNPCs generation and isolation
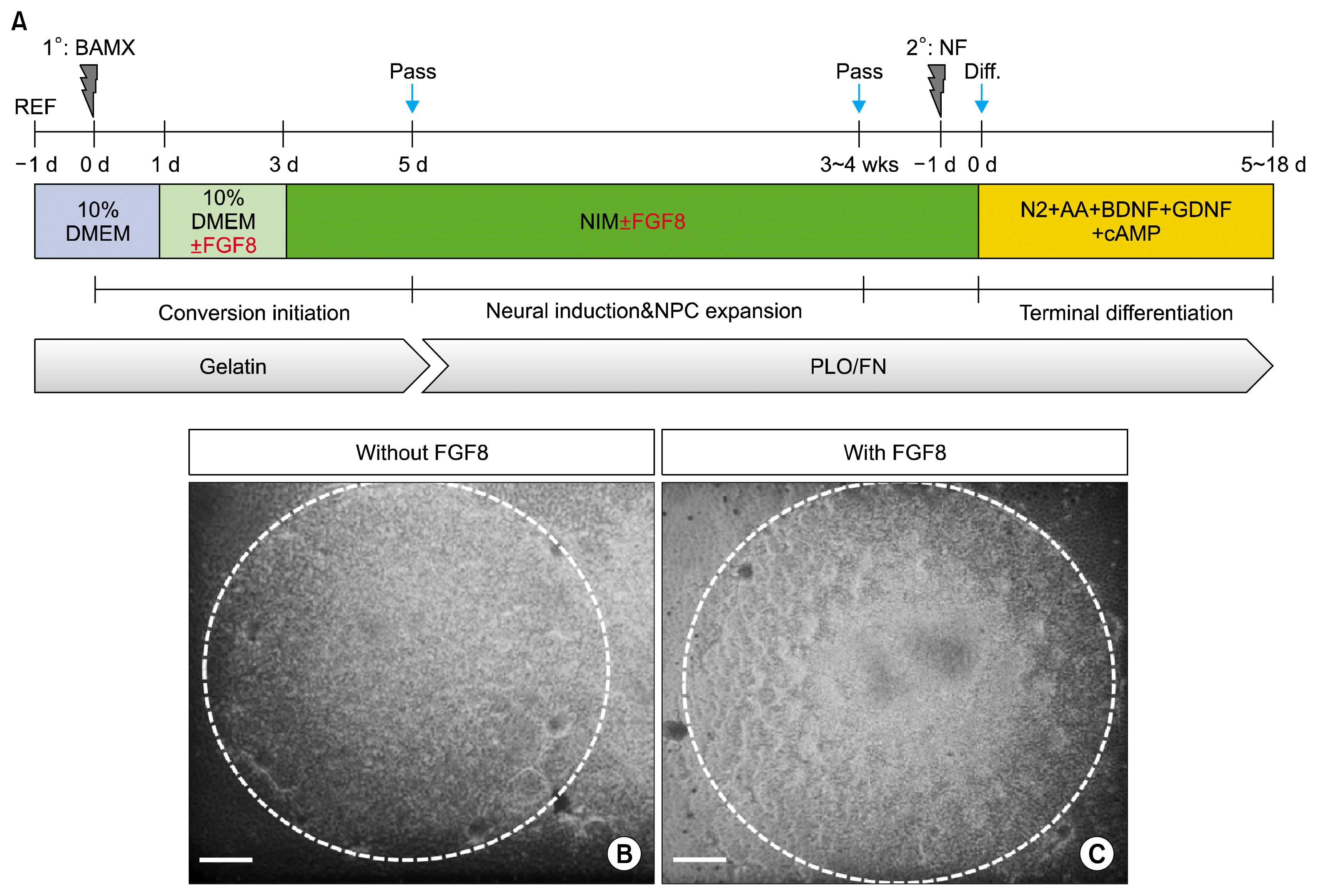
iNPCs generation was performed as previously described (17). Briefly, for generation of iNPCs, fibroblasts were seeded on gelatin-coated culture dishes (0.5~1×106/100 cm dish). The next day, the cells were transduced with four retroviruses (BAMX). After 16~20 hours, the culture media were changed to two different media: 1) fresh fibroblast media containing 100 ng/mL human fibroblast growth factor 8 (FGF8; PeproTech, Rocky Hill, NJ, USA) and 2) fresh media that did not contain FGF8. After two days, transduced cells were placed into neural induction media [NIM: N2 medium supplemented with 20 ng/mL basic fibroblast growth factor (bFGF; R&D Systems, Minneapolis, MN, USA), 100 U/mL recombinant human leukemia inhibitory factor (EMD Millipore, Billerica, MA, USA), 2 μg/mL doxycycline (Sigma-Aldrich) and with/without 100 ng/mL FGF8] and the culture media were changed every other day thereafter. Two days after initiating NIM culture conditions, cells were transferred to six-well culture dishes pre-coated with 15 μg/mL poly-L-ornithine (PLO; Sigma-Aldrich) and 1 μg/mL fibronectin (FN; Sigma-Aldrich) and maintained in NIM. After approximately two to three weeks, cell morphologies changed to neural stem-like cells, as evidenced by small cell size and bipolar morphology. Formation of cell clusters was also observed.
Dopamine neuron generation from fibroblast-derived iNPCs
To generate dopaminergic neurons, iNPCs were transferred onto coverslips (Bellco Glass, Vineland, NJ, USA) pre-coated with PLO/FN. After one day, iNPCs were transduced for two hours with dopaminergic neuron-related factors, Nurr1 and Foxa2 retroviruses, which were cultured overnight in NIM and differentiated the following day in media containing 0.2 mM ascorbic acid (AA; Sigma-Aldrich), 20 ng/mL brained-derived neurotrophic factor (BDNF; R&D Systems), 20 ng/mL glial cell-line-derived neurotrophic factor (GDNF; R&D Systems), and 250 μg/mL dibutyryl cAMP (Sigma-Aldrich) in N2 medium.
Immunostaining of cultured cells
Immunostaining of cultured cells was performed as previously described (18). Cells were photographed using an epifluorescence microscope (Leica Microsystems, Wetzlar, Germany). Measurement of tyrosine hydroxylase (TH) fiber length was computed with the Leica Application Suite (LAS) image analysis package. Primary antibody information is summarized in Table 1.
Cell counting and statistical analyses
Cell counting was performed on a random selection of 5~10 uniform microscopic fields/wells, with 3~4 wells per experimental condition. All values were confirmed with at least three independent experiments. Data were expressed as mean±SEM. When more than two groups were compared, a paired t-test was performed using SigmaPlot for Windows, version 10.0 (Systat Software GmbH, Erkrath, Germany).
Results
FGF8 did not influence the proliferation efficiency in cultured iNPCs
Recently, our study has shown that NPCs can be successfully generated from fibroblasts transduced with BAMX (17). Briefly, after 16~20 hours of transduction, all media were changed to 10% DMEM (−FGF8) or 10% DMEM (+FGF8); two days later, NIM (−FGF8) or NIM (+FGF8) media were applied for neural induction and NPC expansion, after which rat fibroblasts transformed into small and slender cells. Cells with morphologies consistent with a neural lineage were generated within two weeks, and they actively proliferated into cluster-forming cells after another two weeks of growth in culture conditions, as schematized in Fig. 1A. There were no significant differences in the formation of NPC-like cluster cells without or with FGF8 (Fig. 1B and C).
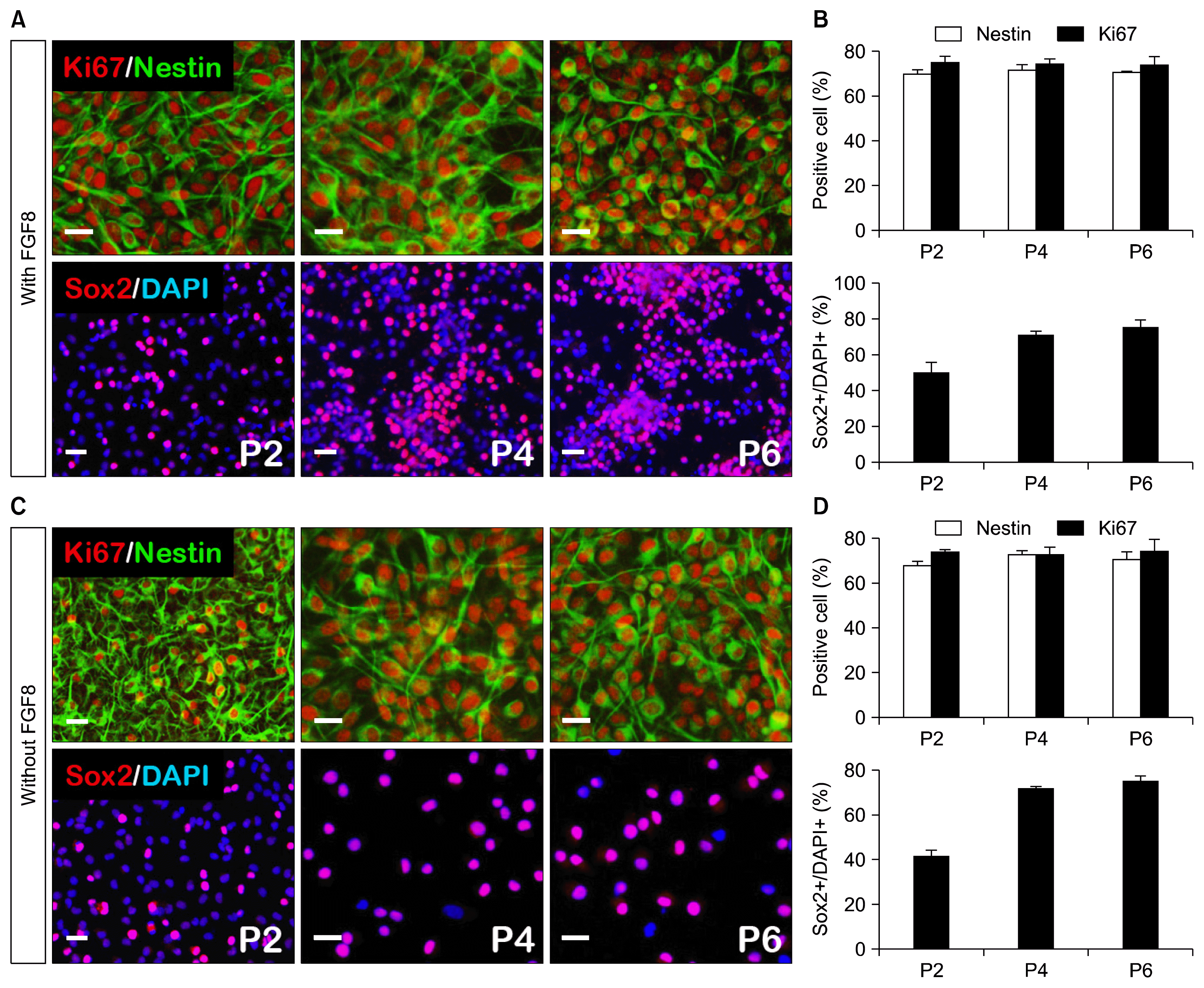
To investigate whether FGF8 has a positive effect on iNPCs proliferation, we compared the characteristics of iNPCs derived from with or without FGF8. After dissociating the mechanically selected clusters into individual cells, they were transferred to PLO/FN-coated dishes and expanded by passaging them every 5~7 days. The iNPC proliferation rate did not differ between these two groups [Fig. 2: with FGF8 and without FGF8, nestin >70%, sox2 >70%, ki67 >70%, each passage (p2, p4, p6)]. Collectively, these findings suggest that FGF8 does not contribute to iNPC proliferation efficiency.
FGF8 is essential for induction and maturation of DA neurons
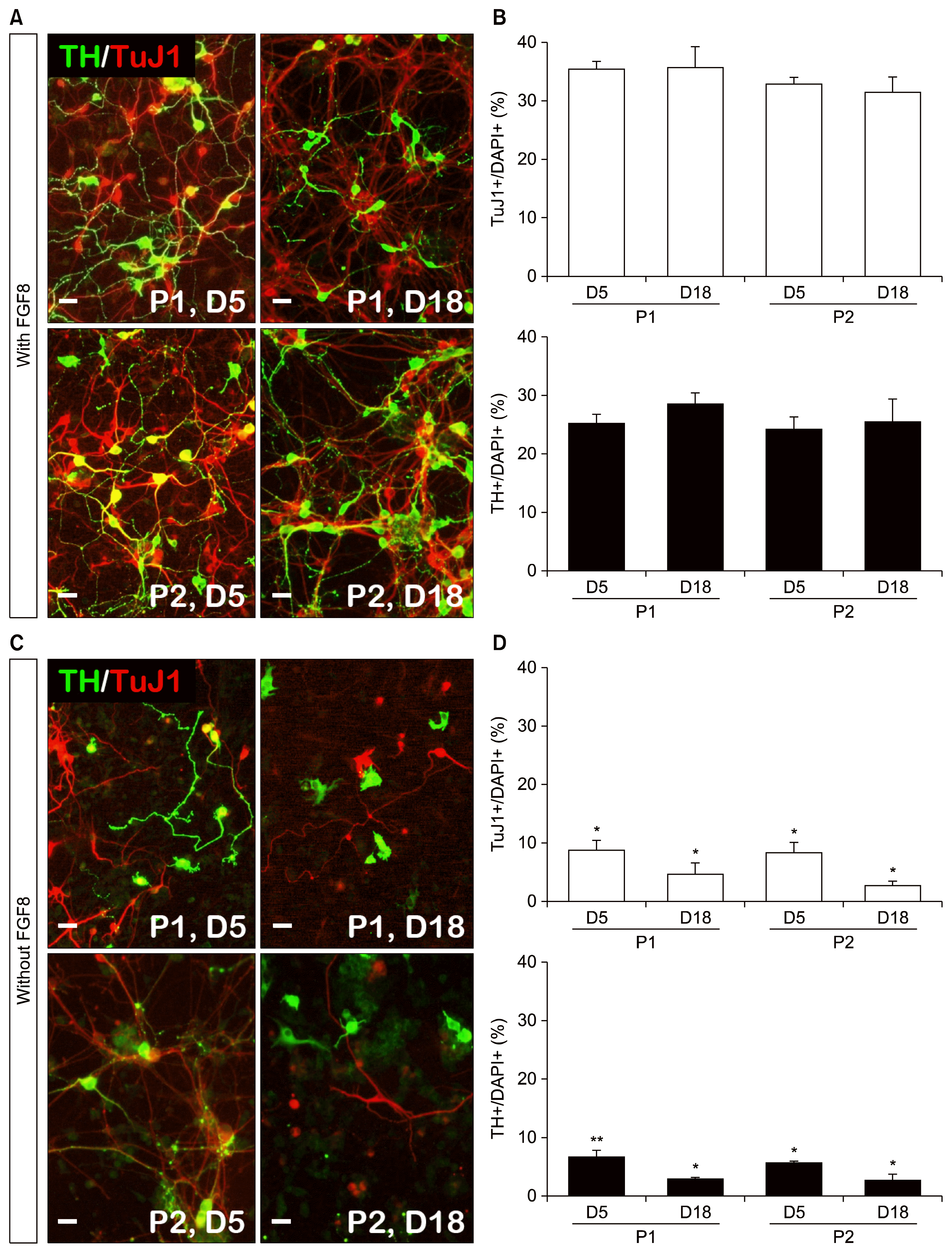
Next, we evaluated the ability of iNPCs to differentiate into DA neurons. To this end, we transduced iNPCs (derived from with FGF8 and without FGF8) with DA neuron-specific factors, Nurr1 and Foxa2. After differentiation 5 and 18 days, passaged p1 and p2 cells revealed that the numbers of TuJ1+ cells and TH+ cells increased in the presence of FGF8 by immunocytochemistry analysis (Fig. 3A and B: TuJ1+ cells: P1-D5 vs. P1-D18; 35.3±1.3% vs. 35.6±3.6%; P2-D5 vs. P2-D18; 32.6±1.3% vs. 31.3±2.7%; TH+ cells: P1-D5 vs. P1-D18; 25.3±1.3% vs. 28.6±1.8%; P2-D5 vs. P2-D18; 24.3±2.0% vs. 25.6±3.7%) compared to the absence of FGF8 (Fig. 3C and D: TuJ1+ cells: P1-D5 vs. P1-D18; 8.6±1.8% vs. 4.6±2.0%; P2-D5 vs. P2-D18; 8.3±1.8% vs. 2.6±1.0%; TH+ cells: P1-D5 vs. P1-D18; 6.3±1.3% vs. 2.6±0.5%; P2-D5 vs. P2-D18; 5.3±0.5% vs. 2.3±1.3%) and a larger number of longer TH fiber lengths were observed (data not shown). These results indicate that mature DA neurons were efficiently generated from the FGF8 that was supplied during iNPC generation stages.
Discussion
Alternative DA neurons can be ideal cell sources for cell replacement therapy for Parkinson’s disease. To obtain DA neurons in vitro, various cell types and growth protocols have been developed. Recently, direct conversion methods have become one of the most popular approaches because they sidestep the issue of pluripotent stem cells, which is the main problem for cell therapy. On the other hand, reprogramming efficiency of direct conversions is lower than that from other cell sources. In order to improve DA neuron generation efficiency, we treated the culture media with FGF8 during the direct conversion and expansion period. Early period of cluster-forming and iNPC proliferation stages, did not experience an improved proliferation rate when grown in the presence of FGF8; one explanation for this is that FGF2, not FGF8, is actually the significant influencing factor. FGF2 is known to promote the survival and proliferation of neural precursor cells (19). However, the DA neuron differentiation rate with FGF8 contrasted greatly with the rate of without FGF8 conditions. FGF8-treated groups highly expressed TuJ1+, TH+, and TuJ1+TH+/TH+ cells after differentiation. In particular, DA neuron maturation was confirmed by TH fiber lengths (data not shown). In our work, FGF8 is a morphogenetic factor that was used to understand patterning and maturation of midbrain-type DA neurons.
Unfortunately, other functional markers for DA neurons were not statistically different between the FGF8+ and FGF8− groups. To explain this, we considered the possibility that absence/presence of shh influences the role of FGF8 in DA neuron generation. Shh is required for numerous developmental processes, including specification of ventral cell types in the central nervous system, such as midbrain DA neurons. shh and FGF8, which are soluble neurotrophic factors, have been shown to regulate the initial stages of DA neuronal development and to specify DA progenitors in the ventral area of the midbrain (20). Here our data strongly implicates FGF8 signaling in the regulation of maturation of iNPC-derived DA neurons in vitro. Further to improve the functionality of DA neurons, we will evaluate the effect of a combination of FGF8 and shh in culture.
 XML Download
XML Download