

 PDF
PDF Citation
Citation Print
Print


Introduction
Approximately 1% of all men suffer from azoospermia in forms of obstructive or non-obstructive, and azoospermic men constitute approximately 10~15% of all infertile men (1). Much efforts were done to treat these infertile men, especially non-obstructive azoospermia, because their treatment is most difficult (2). In spite of hormonal or surgical treatments, in recent years, more attention has been centralized on the cell and stem cell therapy. Stem cell therapy has potential to develop more and being a choice for treatment of male infertility due to dysfunction of germ cells and their inability to differentiate or proliferate (3). This technique attracted significant interest for treatment of sperm deformity and azoospermia.
Moreover, stem cell therapy may help to reduce the effect of cancer therapeutics on reproductive failure in cancer patients. In addition, inter tubular injection of stem cells is used for production of transgenic mice (4) and rats (5). In-vitro previous studies showed germcell lineage can be differentiated from different kind of pluripotent stem cells including mesenchymal stem cells (MSCs) (6), including embryonic stem cells (ESCs) (7), induced pluripotent stem cells (8) and fetal stem cells (9). There are several reports on transplantation of MSCs to induce spermatogenesis in azoospermic models (10). They were isolated from different sources such as bone marrow (11), adipose tissue (12), endometrium (13), and dental pulp (14).
Lassalle et al. (15) injected BM-MSCs into testis to induce differentiation of these cells into germ cells but, they did not notice any differentiation. In another study in mouse and human, adult BM-MSCs grown in-vitro in the presence of retinoic acid were found to express germ cell markers, but there was still a failure to undergo spermatogenesis after transplantation into testes (16). Lue et al. (17) found different findings and showed that transplanting BM-MSCs into testis of a busulfan-treated infertile mouse model could differentiate into germ, sertoli and leydig cells. Cakici et al. (18) demonstrated that the treatment of busulfan-treated rat testes with adipose tissue derived MSCs resulted into morphologically normal spermatogenesis in some tubules after 12 weeks. Monsefi et al. (19) showed that transplanted BM-MSCs could differentiate into germinal cells in seminiferous tubules of Wistar rats. Although the possible mechanisms of healing are unclear, three mechanism might be responsible to recover testicular function during the tissue regeneration process by MSCs: (i) They may differentiate into the target cells via appropriate induction conditions (20); (ii) These cells secrete growth factors to stimulate the resident stem cells to restore the host cell function (21); and (iii) MSCs merged with the endogenous cells recover the injured tissue function (22). Therefore, this study was undertaken to use busulfan for induction of azoospermia and BM-MSCs for its cell therapy and further spermatogenesis in a hamster model.
Go to : 
Materials and Methods
Animals
Twelve albino male Syrian hamsters (Mesocricetus auratus) weighing 90~130 g were provided from Laboratory Animal Center, Shiraz University of Medical Sciences, Shiraz, Iran. They were housed in temperature-controlled rooms (20~22°C) under 12 h light/dark cycle. The hamsters were fed with standard commercial chow diet ad libitum. This project was approved by the Ethical Committee of Shiraz University of Medical Sciences (Shiraz, Iran). All efforts were made to minimize suffering during the experimental period.
Isolation and culture of hamster BM-MSCs
To isolate BM-MSCs, hamsters (n=2) were anesthetized by ketamine (intraperitoneal injection, 100 mg/kg, Woerden, Netherlands) and xylazine (intraperitoneal injection, 7 mg/kg, Alfazyne, Woerden, Netherlands) and then were euthanized by cervical dislocation. Incision was made on the skin and both femurs and humeruses and their muscular tissues were completely removed. After cutting the bones extremities, their bone marrows were washed and collected in falcon tube using a syringe full of Dulbecco’s modified Eagles medium (DMEM; Gibco, USA) and 1% penicillin and streptomycin (Sigma, USA).
The flushed marrow was centrifuged (5 min, 1500 rpm) and the supernatant was removed, and the pelleted fraction was resuspended in 5 ml DMEM. The suspension were transferred into 25 ml flasks containing DMEM, 10% fetal bovine serum (FBS; USA), and 1% penicillin and streptomycin. The flasks were in an incubator with 5% CO2 at 37°C and saturated humidity. This initial passage of the primary cell culture was referred to as “Passage 0”. Non-adherent cells with the medium were removed after 24 h and fresh medium was added again. The culture medium was changed every three days. When the cells reached 80~90% confluence, the cells were harvested using 0.25% trypsin (Gibco, USA), counted and passaged into new flasks.
To obtain a sufficient number of cells, they were sub-cultured two times using standard methods of trypsinization. Cells were collected in the third passage and were counted using a hemocytometer. They were kept in freezing media including 10% dimethyl sulfoxide (DMSO; MP Bio, France) and 90% FBS, at a density of 2×106 viable cells/ml and were aliquoted into sterile plastic cryovials that were labeled with the freezing serial number and the date. The vials were kept at −20°C for 60 min and then they were transferred to −70°C for 24 h, and finally transferred to liquid nitrogen for long-term storage.
Before surgical treatment, the cryovials were removed from the liquid nitrogen and quickly thawed in a 37°C water bath. When the ice clump was almost thawed, 1 ml of cell culture medium (89% DMEM, 10% FBS, and 1% penicillin and streptomycin) was added, and centrifuged at 1500 rpm for 5 min. The BM-MSCs were transferred into a flask with gently blown into uniform single cell suspension, and cultured at 37°C and 5% CO2. All BM-MSCs were subcultured just up to passage 4.
Busulfan treatment of hamsters and BM-MSCs transplantation
Five untreated male hamsters served as negative control and 5 male hamsters were used as a model of inducing azoospermia by injection of two dose of busulfan (10 mg/kg body weight, intraperitoneally, Busilvex®, Pierre Fabre Medicament, Boulogne, France) with 21 days interval. Thirty five days after the last busulfan injection, male hamsters were anaesthetized by ketamine and xylazine. Their abdominal area was surgically prepared in dorsal recumbency and 1 cm midline abdominal incision was made to expose the peritoneal cavity.
Under a stereomicroscope (Model SZN, Optika, Italy), the fat pad attached to the right epididymis and testis was gently pulled using an iris forceps. The testis was exteriorized and the testicular artery and epididymis were clearly visible. A thin sterile black plastic card with a 30 degree V-like cut was put underneath the testis as a holder. Suspension of BM-MSCs was mixed with sterile trypan blue (1:1, v/v) and was loaded into the polyethylene tube connected to a 1 ml syringe (Fig. 1A). A pulled pipette was attached into the tube. By applying pressure to the syringe, the cell suspension was gently forced into the pipette.
 | Fig. 1(A) Designed micro-injector consistent of a 1 mL syringe (without needle), tube of a 24 gage butterfly needle, and a pre-pulled glass pipette. (B) To make a good contrast between the translucent efferent duct and the fat tissue and the membrane around it, a triangular hard black plastic card was inserted underneath the duct. (C) Bone marrow-derived mesenchymal stem cells (BM-MSCs) transplantation (106 cells were mixed with trypan blue) into efferent duct of hamster testis. (D) Seminiferous tubules were filled with mixture of BM-MSCs and die.
|
The efferent duct that connects the testis to the epididymis was identified and fat tissue was gently removed around the ducts using a sharp needle. To make a good contrast between the translucent duct and the fat tissue and membrane around it, a triangular hard black plastic card was inserted underneath the duct (Fig. 1B). The pipette was carefully inserted into a duct in the bundle of efferent duct and gently threaded a few mm toward the testis (Fig. 1C). While avoiding moving the injection pipette, the plunger of the syringe was gently depressed and the blue suspension flowed into the rete testis. Then, almost all surface seminiferous tubules were filled with suspension (Fig. 1D). BM-MSCs’ mixture (106 cells in 100 μL) was completely injected. The testis was returned to the abdominal cavity and the abdominal wall and the skin were sutured. The untreated left testis was served as positive control.
BM-MSCs osteogenic and adipogenic differentiations
For in-vitro osteogenic differentiation, cells at 90% confluence were cultivated in DMEM, 15% FBS, 200 μM L-ascorbic acid, 10 mM glycerolphosphate, and 100 nM dexamethasone. The medium was changed twice a week for 3 weeks. After 21 days, osteogenic differentiation was evaluated using Alzarin red staining. In brief, BM-MSCs cultures were fixed with 4% paraformaldehyde for 10 minutes. Then cells were incubated for 20 minutes at room temperature in 1% Alizarin red and 1% ammonium hydroxide. Following incubation, cultures were washed 4 times, 5 minute each time with 1 ml dH2O replacing the water at each 5 minute interval and air-dried. Alizarin red dye binds to calcium ions present in mineralized deposits resulting in a brilliant red staining (all reagents from Sigma-Aldrich, USA).
For adipogenic differentiation, cells at 90% confluence were cultivated in DMEM, 15% FBS, 0.2 mM L-glutamine, 100 μM L-ascorbic acid, 200 μM indomethacin, and 100 nM dexamethasone. The medium was changed twice a week for 3 weeks. After 21 days, adipogenic differentiation was evaluated using Oil Red O staining. In brief, for evaluating the generation of oil droplets, the BM-MSCs were fixed in 10% formalin for 10 min at room temperature and washed twice with water. Oil red O working solution was prepared by adding 6 mL of stock solution (0.5 g oil red in 100 mL isopropanol) to 4 mL distilled water, mixed, and filtered through Whatman filter paper. Next, oil red O stain was added and incubated for 1 h at room temperature. Finally, the cells were rinsed several times with water and observed under an inverted microscope (all reagents from Sigma-Aldrich, USA).
Characterization of BM-MSCs by reverse transcription polymerase chain reaction (RT-PCR)
To quantify the expression of BM-MSCs specific markers (CD29 and CD73) and absence of hematopoietic stem cells specific marker (CD45), RT-PCR was performed. First of all, total RNA was extracted using the Column RNA Isolation Kit (Denazist-Asia, Iran) in accordance with the manufacturer’s instructions. Total RNA concentration was evaluated by spectrophotometer. After that, complementary DNA (cDNA) synthesis from RNA samples was done by using AccuPower Cycle Script RT PreMix Kit (Bioneer, Korea) according to the manufacturer’s instructions.
Briefly, 15 μL of total RNA was used for each reaction and the volume had been reached up to 20 μL with the DEPC water. Twelve thermal cycles was performed with the following way: 30 sec at 20°C for primer annealing, 4 min at 42°C for cDNA synthesis, 30 sec at 55°C for melting secondary structure and cDNA synthesis and 5 minutes at 95°C in order to heat inactivation. In the third step, 1 μL of template (cDNA) was mixed with other reagents consisting of PCR buffer, MgCl2, H2O, dNTPs, Taq DNA polymerase, forward and reverse primers (CD29, CD73, and CD45) (Table 1). Then, microfuge tubes containing of 20 μL above mixing were put in Thermocycler (Eppendorf Mastercycler Gradient, Eppendorf, Hamburg, Germany). Thirty amplification cycles were run, consisting of 30 sec denaturation at 95°C, 30 sec annealing at 64°C and 30 sec extension at 72°C with the 2 min at 95°C for primary denaturation and 5 min at 72°C for final extension. PCR products were evaluated for the presence of considered bands by gel electrophoresis with the aid of DNA safe stain in 1.5% agarose gel medium. Produced bands were visualized under UV radiation by Gel documentation system (UVtec, Cambridge, UK).
Table 1
Sequences of RT-PCR primers used to quantify the expression of bone marrow-derived mesenchymal stem cells specific markers (CD29 and CD73) and hematopoietic stem cells specific markers (CD45) in hamster
![]()
Histomorphometric analysis of testes
Thirty five days after cell injection, animals were euthanized with ether and their testes and epididymis tails were collected and fixed in a 10% formalin buffer solution. After fixation, segments were embedded in paraffin, and histological sections were made from each block. The 5 μm thickness hematoxylin-eosin stained sections were examined under light microscope for any spermatogenic activity.
For each testis five vertical sections from the polar and the equatorial regions were sampled. In one cross-section per animal, all tubules were evaluated for presence of spermatogonia, spermatocytes and spermatids. In 10 circular transverse sections of tubules, each from a different region of the testis and each homogeneous throughout with respect to cell associations and spermiogenesis, we measured inner, outer and total diameters, calculated areas of the cellular and luminal regions and cross sectional area of the tubule, and counted the number of late spermatids present.
The mean seminiferous tubule diameter (D) was derived by taking the average of two diameters, D1 and D2 at right angles. Cross-sectional area (Ac) of the seminiferous tubules was determined using the equation Ac=πD2/4, where π is equivalent to 3.142 and D the mean diameter of seminiferous tubules (23). A testis was rated for its spermatogenic potential (modified spermatogenic index) on a modified scale of 0 to 5 (23). The index was based on the appearance of the spermatogenic cells throughout the testis and included number of cell layers, types of cells, and the presence of late spermatids in the seminiferous tubules. The index and criteria were as follows: 0, no spermatogenic cells; 1, only spermatogonia present; 2, spermatogonia and spermatocytes present; 3, spermatogonia, spermatocytes and round (early) spermatids present with <50 late spermatids per tubule; 4, spermatogonia, spermatocytes, and round spermatids present; and up to 50~100 late spermatids per tubule; and 5, all cell types present and >100 late spermatids per tubule.
Statistical analysis
Means and standard error (SE) of the data of histomorphometric indices of seminiferous tubules were subjected to Kolmogorov-Smirnov test of normality and analyzed by one-way ANOVA (SPSS for Windows, version 11.5, SPSS Inc, Chicago, Illinois), and post-hoc test was performed by Tukey test. The spermatogenesis index of seminiferous tubules was compared using Mann-Whitney U test. The p value of less than 0.05 was considered to be statistically significant. Group means and their standard error were reported in the text and graphs (GraphPad Prism version 5.01 for Windows, GraphPad software Inc., San Diego, CA, USA).
Go to :
Results
Culture of BM-MSC
BM-MSCs showed a fibroblast-like, spindle-shaped morphology after they attached to the culture flasks. BM-MSCs proliferation started 3~4 days after incubation until reaching an 80% confluence (Fig. 2).
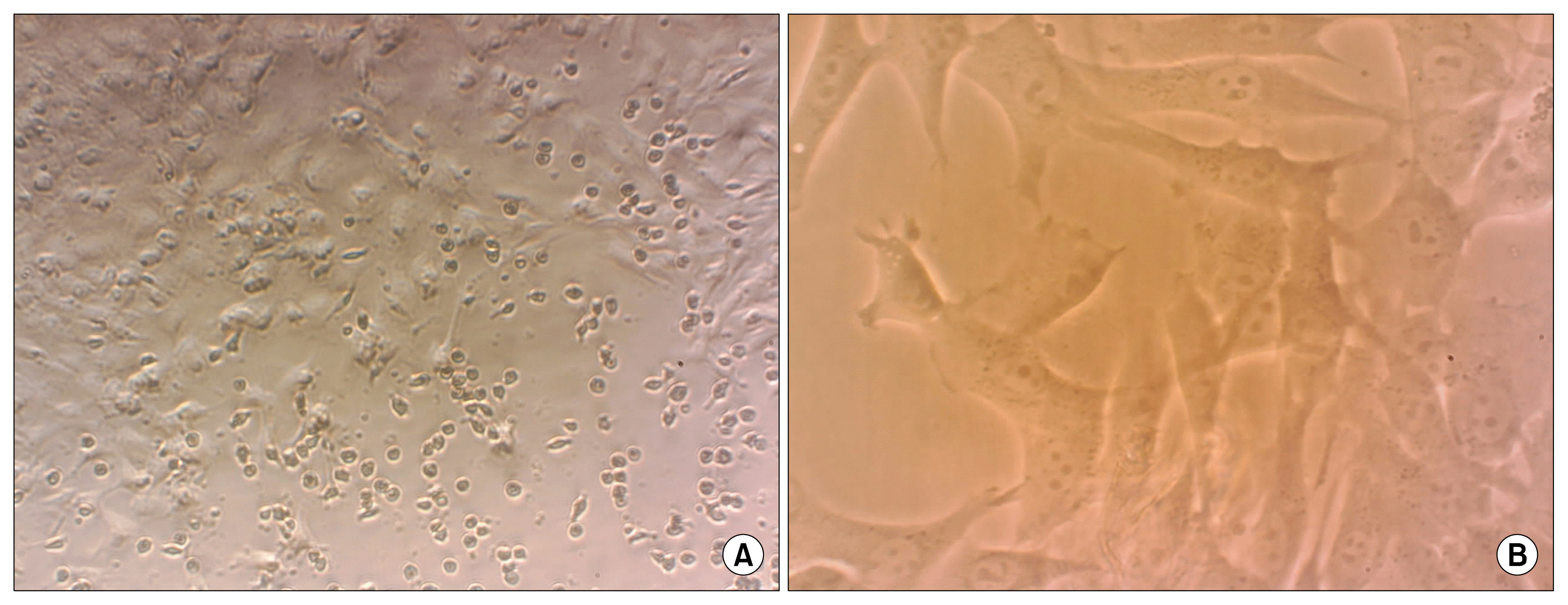 | Fig. 2Morphological and phenotypic characteristics of hamster bone marrow mesenchymal stem cells. (A) At passage 0, earlier stage. Diverse morphologies including attached spindle-shaped and flattened cells and round other bone marrow cells. (B) Stem cells exhibited large, flattened or fibroblast-like morphology in passage 3.
|
BM-MSCs osteogenic and adipogenic differentiations
To further confirm the differentiation capacity of BM-MSCs, osteogenic and adipogenic differentiations were induced. After culture of BM-MSCs in osteogenic differentiation medium, the cells differentiated toward osteoblasts as verified by positive staining with Alzarin Red staining (Fig. 3A). BM-MSCs treated by the adipogenic differentiation protocol showed the presence of intra-cellular lipid droplets, which were confirmed by Oil Red O staining (Fig. 3B). The BM-MSCs grown in culture medium alone (undifferentiation) did not show any osteoblast characters or lipid droplets at any of the time points examined, and maintained their typical fibroblast-like shape (Fig. 3C).
 | Fig. 3Bone marrow-derived mesenchymal stem cells of hamster cultivated in (A) osteogenic medium and stained with Alizarin Red (×100), (B) in adipogenic medium and were stained with Oil Red O at day 21 after induction (×200), and (C) in culture media without differentiation medium as control (×100). (D) Activation of specific mesenchymal marker (CD29 and CD73) compared with deactivation of specific hematopoietic marker (CD45) of hamster bone marrow-derived stem cells.
|
Characterization by RT-PCR
According to proof of the mesenchymal property of hamster BM-MSCs, these cells were expanded up to passage 3 and then they were analyzed using a RT-PCR assay. Fig. 3D displays positive expression for CD29 and CD73 (mesenchymal cell marker) and negative expression for CD45 (hematopoietic cell marker).
Histological assessment of spermatogenesis
After induction the azoospermia in the hamsters with double injection of 10 mg/kg of busulfan, the testes of the control hamsters were microscopically examined for any spontaneous spermatogenesis recovery and no sign was observed. After the treatment with busulfan, the seminiferous tubules of the testes were not treated with BM-MSCs were empty and their spermatogenesis process was disrupted (Fig. 4D). The examination of sections showed only sertoli cell appearance for samples without BM-MSCs therapy.
 | Fig. 4(A) Sections of seminiferous tubules of normal control hamster. Tubules have condensed spermatogenic epithelium. (B) Sections of seminiferous tubules of treated hamster with bone marrow mesenchymal stem cells. Most of tubules appeared to be filled up with spermatogenic cells. (C) Presence of different kind of germinal epithelial cells in stem cell treated seminiferous tubules. (D) Busulfan treated azoospermic control seminiferous tubules. The seminiferous tubules were partially empty and without vacuolated germinal layer cells indicating the absence of spermatogenesis. Hematoxylin and eosin staining.
|
Histological examinations of testes in busulfan treatment group after two months revealed some degenerative changes such as seminiferous tubular atrophy and germinal epitheliums degenerations in the most of tubules. The large vacuolated lumen occupied seminiferous tubules and the atrophic germinal epithelium covered peripheral zone of seminiferous tubules as thin band. Moreover, there were no spermatozoa in the epididymis of busulfan treated azoospermic group (Fig. 5B).
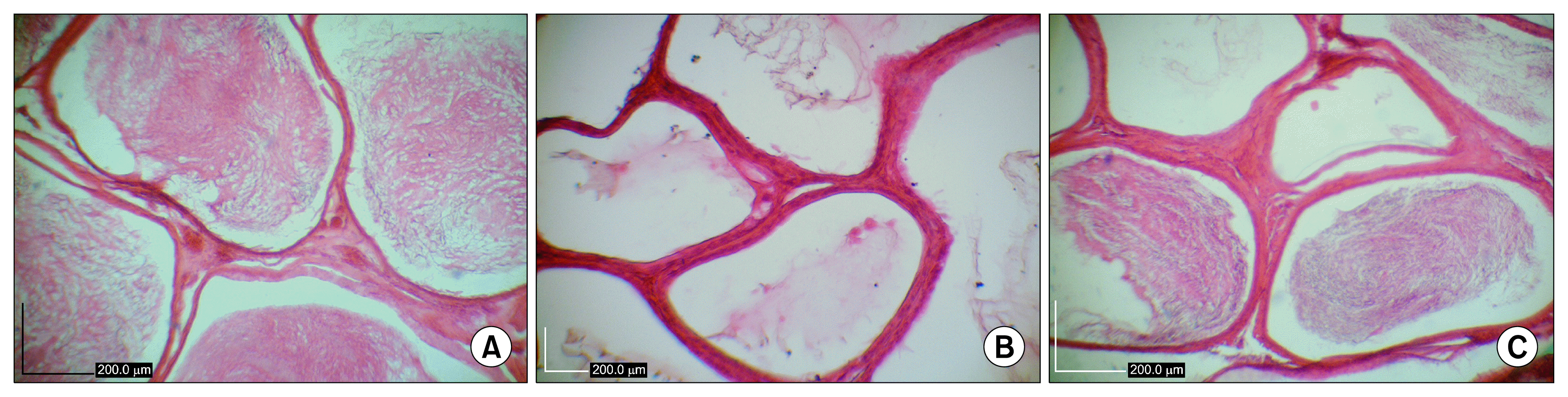
However, the presence of spermatogonia in the seminiferous tubules with BM-MSCs transplantation was observed (Fig. 4B). Moreover, the tubules appeared to be filled up with germinal cells (spermatogonia, primary spermatocytes, spermatids and sperms) in the sections of BM-MSCs-treated testis (Fig. 4C). Most of the epididymis tubes of cell treated groups have spermatozoa, but there was some tubes were empty (Fig. 5C). In comparison with treated group, normal intact hamsters had more condensed germinal epithelium (Fig. 4A) and all of epididymis tubes were filled by spermatozoa (Fig. 5A).
Histomorphometric measurements indicated the lumen diameter and luminal area of the seminiferous tubules in hamsters with stem cell transplantation were less than the azoospermic hamsters (p=0.001 and p=0.001, respectively) and more than normal hamster (p=0.001 and p=0.001, respectively; Figs. 6A and 6B). Cellular diameter and cellular area of the seminiferous tubules in hamsters with stem cell transplantation were more than azoospermic and normal control groups (p=0.001 and p=0.001, respectively; Figs. 6B and 6C). However, cellular diameter of normal control group was more than azoospermic hamsters (p=0.001), but cellular area of these two groups were not different (p=1.0).
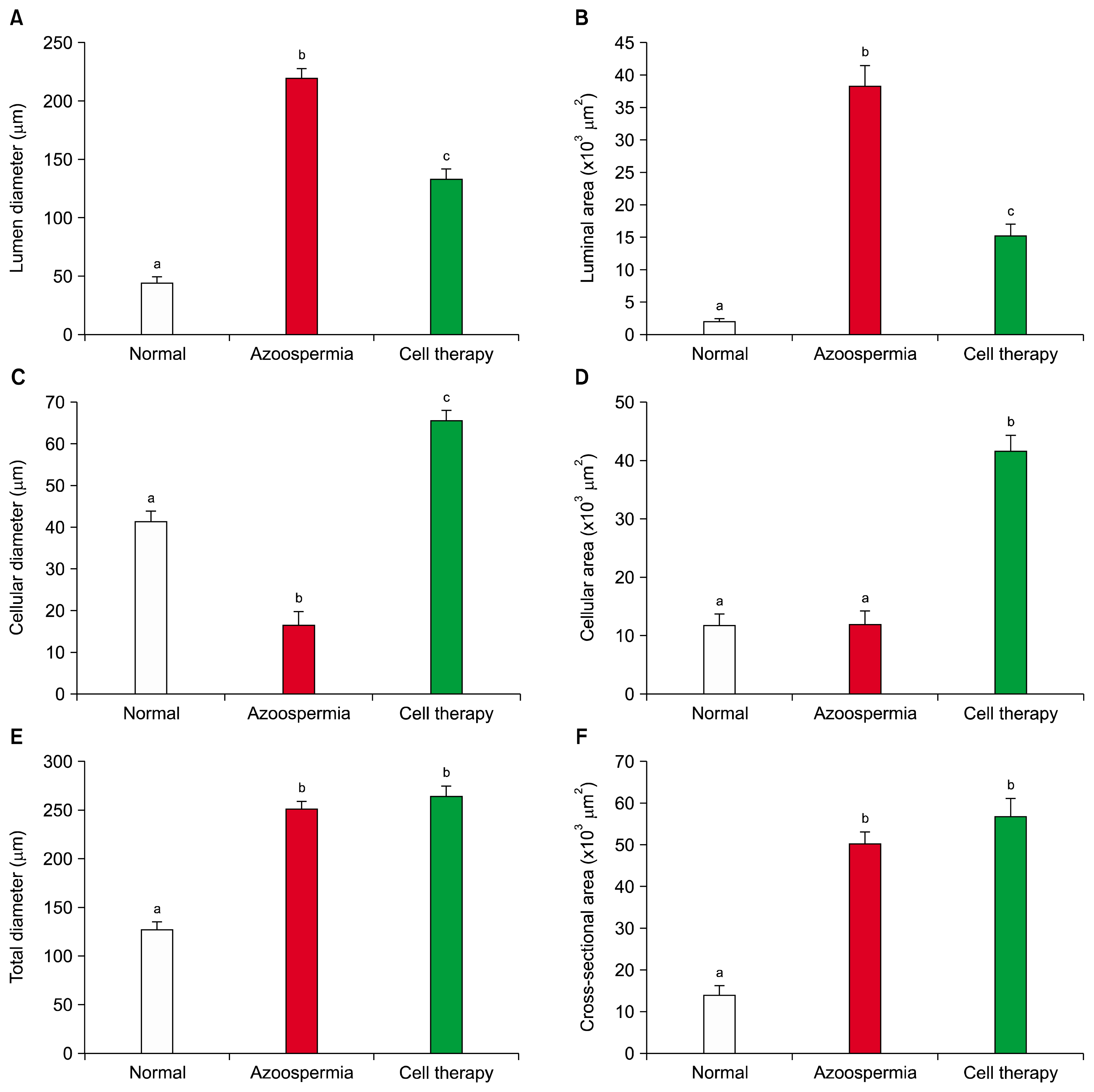 | Fig. 6Mean and standard error of histomorphometric indices of seminiferous tubules in busulfan-induced azoospermic testis treated with bone marrow mesenchymal stem cells (cell therapy) in comparison with busulfan treated testes (azoospermia) and intact normal testis (normal) in hamster. (A) Lumen diameter (μm), (B) Luminal area (μm2), (C) Cellular diameter (μm), (D) Cellular area (μm2), (E) Total diameters (μm), and (F) Cross sectional area of the tubule (μm2). a,b,cdifferent superscript letters show significant differences between groups (p<0.05).
|
Total diameter and cross sectional area of the seminiferous tubules in hamsters with stem cell transplantation were not different with azoospermic hamsters (p=0.7 and p=0.5, respectively; Figs. 6D and 6E) but both groups were more than the normal control (p=0.001, p=0.001, p=0.001 and p=0.001, respectively).
Spermatogenesis index of seminiferous tubules in hamsters with stem cell transplantation did not have difference with normal control hamsters (p=0.7), but both groups were more than the positive control hamsters (p=0.001 and p=0.001, respectively; Fig. 7).
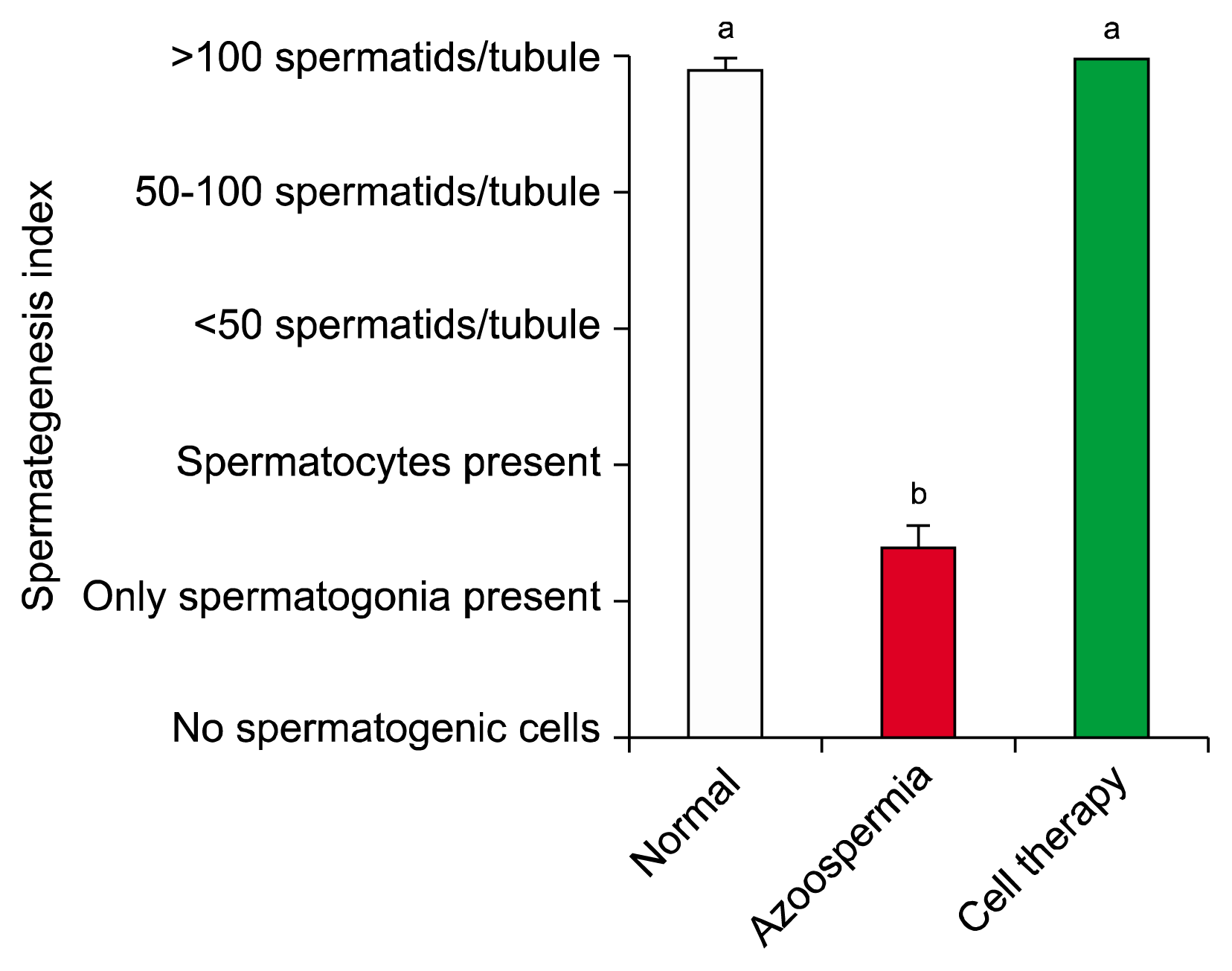 | Fig. 7Mean and standard error of spermatogenesis index of seminiferous tubules in busulfan-induced azoospermic testis treated with bone marrow mesenchymal stem cells (cell therapy) in comparison with busulfan treated testes (azoospermia) and intact normal testis (normal) in hamster. a,bdifferent superscript letters show significant differences between groups (p<0.05).
|
Go to :
Discussion
In the present study, BM-MSCs interactions in the hamster testis were evaluated functionally by transplantation of busulfan-induced azoospermic hamster testis. Injected BM-MSCs induced spermatogenesis. However, the damaged somatic environment of the testis is one of the most problem and limiting step in this method of infertility treatment. Damaged testis is not receptive to spermatogonial stem cells transplantation and thereby not able to treat patient azoospermia (24). Interestingly, BM-MSCs are not only hypo-immunogenic but also produce immunosurveillance or immunosuppression upon transplantation, therefore they are suitable for allogeneic transplantation (25). Intravenously bone BM-MSCs transfusion showed immunomodulatory effects on antisperm antibody production in allogeneic settings in mice after testis rupture (26). Moreover, sertoli cells are immune tolerant cells (27) and they can cause survival of the donor BM-MSCs and protect the transplanted allogeneic cells from post transplantation immune or inflammatory reaction.
In the present study, spermatogenesis index of seminiferous tubules in hamsters with stem cell transplantation did not have difference with normal control hamsters. Seminiferous tubules provide cyclic and dynamic regulation of spermatogenesis, and sertoli cells prepare a microenvironment that induces spermatogonial proliferation and differentiation. Our results also showed that BM-MSCs can induce reconstitution of the tubular microenvironment which helps remained inactivated germinal cells to proliferate in the host seminiferous tubules.
In some studies combination of in-vitro differentiation and in-vivo transplantation was performed to obtain advanced differentiated spermatozoa (28). Several in-vivo studies have been performed to evaluate the differentiation potential of mesenchymal stem cells into spermatozoa in rat and mice animal models. However, Van Saen et al. (29) and Lassalle et al. (15) showed that BM-MSCs could not differentiate into sperm in mice, but in the other study, transplanted mouse BM-MSCs have been demonstrated to form germ cells in-vivo (17). Moreover, BM-MSCs was differentiation into germ cells and spermatozoa in-vitro (30). Nayernia et al. (16) demonstrated that murine BM-MSCs are able to differentiate into early germ cells in-vitro and in-vivo in mice.
Allogeneic BM-MSCs trans-differentiated into spermatogenic-like-cells and enhanced endogenous fertility recovery in rat by injection into seminiferous tubules of recipient with busulfan induced (2, 19, 31) and testicular torsion (32) azoospermia rat. Mehrabani et al. (33) and Cakici et al. (18) demonstrated that adipose-tissue-derived MSCs can leading to recovery of fertility in the busulfan-treated azoospermia rat model. Recently, Chen et al. (34) showed that sperm differentiation potential of transplantation of human umbilical cord MSCs into immunodeficient mouse seminiferous tubule. Therefore, success in MSCs therapy of azoospermia in animal models may be species dependent.
Various conditions can cause non-obstructive azoospermia, including congenital or genetic abnormalities, exposure to gonadotoxins, infectious issues, varicocele, trauma, medications such as chemotherapy reagents, endocrine disorders, and idiopathic causes. Current medical therapy, including hormone or surgical methods, shows little benefit to non-obstructive azoospermia (35). Busulfan is a chemotherapeutic agent that most often use as low dose in a long time manner to treat chronic myeloid leukaemia (36). Before allogeneic transplantation of hematopoietic cell, busulfan is used as a myeloablative agent (37). Despite of its beneficial effects, cells with high division activities such as germ cells are more susceptible to busulfan side effects.
Spermatogonial stem cells of different species can be destroyed by busulfan (23, 38). But it has no effects on DNA synthesis. However, when busulfan intoxicates the cells in the G1 phase, inhibits the next mitosis (39). Busulfan-treated hamsters are excellent and well-established recipients for evaluating stem cell activity in hamster testis cell populations (38). They were used as recipients in this study.
On the other hand, morphometric findings of the present study showed a new feature of azoospermia cell therapies. Total diameter and cross sectional area of the seminiferous tubules in hamsters before and after transplantation were more than the normal control. Moreover, the lumen diameter and luminal area of the seminiferous tubules in hamsters with stem cell transplantation were more than the azoospermic hamsters and less than normal hamster. These morphostructural change can be explained that after induction of azoospermia some tubes collapsed and even after cell therapy, the tubular structures of seminiferous which were treated in BM-MSC treated groups and untreated ones in azoospermia groups had more space to grown by the intratubular hydrostatic pressure of fluid and cell grown.
The intratubular hydrostatic pressure of hamster seminiferous tubules are 2~3 mm Hg (40). Increase of total diameter may reduce the contraction ability of peritubular layer of contractile myofibroblast cells which may resulted in observed decrease of spermatozoa in epididymis of BM-MSCs treated hamsters. In addition, cellular diameter and cellular area of the seminiferous tubules in hamsters with stem cell transplantation were more than azoospermic and normal control groups. These were caused by increasing total diameter of treated animals’ seminiferous tubules and increase of space between sertoli cells resulted in more places for cell proliferation.
Interestingly, however, cellular diameter of normal control group was more than azoospermic hamsters, but cellular area of those two groups were not different. Increase of cross sectional area of tubules in azoospermic hamster and moreover, greater space for non-busulfan affected Sertoli cells helps the cells to cover a more two-dimensional area than a normal groups.
Go to :
Conclusion
Cell therapy could help the fast repair of pathological changes in testicular seminiferous tubules. It seems BM-MSCs were found effective in treating animal model of azoospermia. The fertile status of busulfan induced azoospermic hamsters was recovered by transplantation of BM-MSCs. This finding raises the possibility of using BM-MSCs to treat azoospermia in human. But, more similar studies are need in different animal models to approve using of this technique in treatment of azoospermia and infertility in human.
Go to :
 XML Download
XML Download