

 PDF
PDF Citation
Citation Print
Print


Introduction
Stem cell therapy is a promising approach for treating degenerative diseases by replacing dead or vulnerable neurons. Parkinson’s disease (PD) may benefit from this approach where motor function could be restored by replacing degenerated nigrostriatal dopaminergic neurons (1). The proof of principle for such an approach has been established in animal models of hemi-Parkinsonism where transplantation of embryonic stem (ES) or induced pluri-potent stem (iPS) cell-derived dopaminergic precursors integrated and differentiated within host’s striatum with significant recovery of motor functions (1–3). However, a practical clinical adaptation of the stem cell approach requires that stem cells are [1] available in sufficient quantities, [2] non-immunogenic, and [3] non-tumorigenic. While both ES and iPS cells are self-renewing thus available in sufficient number to support practical clinical applications, only the latter, generated from patients’ cells would qualify on the criterion of immunological compatibility. However given their pluripotency, both suffer from the proclivity to form tumor. In addition, iPS cells generated by the nucleic acid approaches carry the risk of insertional mutagenesis (4). These barriers necessitate the identification of accessible, renewable, and safe sources of stem cells that can sustain an autologous stem cell therapy for Parkinson’s disease. One such source could be mesenchymal stem cells (MSCs) (5). However, these cells may not be a practical solution in the current state given the low efficiency of dopaminergic conversion of naïve MSCs and issues related to insertional mutagenesis where the conversion efficiency was enhanced by the expression of exogenous genes (6).
A heterologous source of accessible adult cells for dopaminergic neurons could be the adult limbus, a regenerative ocular tissue (7). These cells, safely isolated through the biopsy without affecting the vision, and efficiently expandable in number in vitro, have been successfully used in autologous stem cell approaches to treat the damaged cornea with the recovery of vision (8). We have observed that limbal progenitors, when cultured in reduced BMP4 signaling, acquire neural properties (9, 10). Molecular screening of these cells revealed that they also expressed Nurr1, a transcription factor involved in the differentiation and survival of dopaminergic neurons (11), suggesting that, given the conducive culture conditions, they might differentiate along the mesencephalic dopaminergic lineage. Here, we examined this notion and first demonstrated that the limbus-derived neural progenitors, under the influence of embryonic hippocampal cells, differentiated into generic functional neurons. Second, when subjected to protocol for the directed differentiation of ES cells into dopaminergic neurons, the limbus-derived neural progenitors activated regulators and acquired phenotype and functional characteristics of dopaminergic neurons, which included ability to synthesize dopamine. Their intrastriatal transplantation in the rat model of hemi-Parkinsonism was associated with a reduction in the amphetamine-induced rotations. No tumor formation was observed 6 weeks post-transplantation. These preliminary observations thus posit limbus-derived neural progenitors as an accessible and immune compatible source from which dopaminergic neurons can be derived non cell-autonomously for a safe autologous ex-vivo stem cell approach to Parkinson’s disease.
Go to : 
Materials and Methods
The study was approved by the Institutional Animal Care and Use Committee (IACUC), at University of Nebraska Medical Center, and animals were housed in the Department of Comparative Medicine at University of Nebraska Medical Center.
Neurosphere assay
Neurosphere assay to derive neural progenitors from the limbus was carried out as previously described (10). Briefly, following the enucleation of eyes, the entire cornea was removed by a circular incision below the limbus. The limbal region was carefully dissected away from the central cornea and cut into small pieces. Tissues were incubated first in 0.05% trypsin (Sigma) and then in 78 U/ml of collagenase (Sigma) plus 38 U/ml of hyaluronidase (Sigma) at 37°C for 27 and 30 min, respectively and dissociated into single cells by trituration. Dissociated cells were cultured in DMEM: F12 supplemented with 1x N2 supplement (Gibco), 20 ng/ml of EGF (Collaborative Research), 10 ng/ml of FGF2 (Collaborative Research) and 100 ng/ml of Noggin (R&D Systems) at a density of 105 cells/cm2. After a week in culture, resulting neurospheres of cells were plated on poly-D-lysine and laminin coated glass coverslips. BrdU (10 μM) was added to the last 8~12 hours of culture to tag proliferating cells. Medium was changed every other day.
Pan neuronal differentiation
To promote pan neuronal differentiation, neurospheres were co-cultured with embryonic day 18 (E18) hippocampal cells using a Millicel CM insert (pore size, 0.4 mm, Millipore) in the medium defined above, except it lacked mitogens and was supplemented with 1% FBS and 1ng/ml of BDNF (Collaborative Research). Neurospheres were plated in 24-well tissue culture plates and allowed to settle. Inserts containing the hippocampal cells were slowly and carefully lowered into wells to avoid any spill. Medium was changed without removing the insert. Controls consisted of neurospheres cultured in the mitogen-free medium supplemented with serum and factors as above.
Dopaminergic differentiation
The directed differentiation of limbus-derived neural progenitors along the dopaminergic lineage was achieved by using the Mouse Dopaminergic Neurons Differentiation Kit (R&D Systems) as per vendor’s instructions. Following differentiation, cells were collected for RT-PCR analysis or fixed with 4% paraformaldehyde for 15 min at 4°C for immunocytochemical analysis. Briefly, limbal neurospheres were cultured in the presence of ITSFn for neural induction (Stage I), differentiation of dopaminergic neurons under the influence of FGF8 and Shh (Stage II), followed by selection of dopaminergic neurons in the presence of ascorbic acid (Stage III).
Immunofluorescence analysis
Immunofluorescence analyses were carried out as previously described (10). Briefly, cells or cryosections were incubated with specific antibodies [βIII-tubulin (Covance), Nestin (DSHB), TH (Cell-Signalling), and EN1 (Hybridoma Bank)] in blocking serum overnight at 4°C. Following incubation in species-specific IgG antibodies conjugated to CY3, incorporation of BrdU was detected by using FITC conjugated anti-BrdU (Sigma). Images were captured by a CCD cool digital camera (Princeton Instruments) using image-capturing software (OpenLab).
RT-PCR analysis
Isolation of total RNA from cells and synthesis of cDNA were carried out as previously described (6). cDNA (2 μl) was amplified by using the forward and reverse gene-specific primers using step cycles (denaturing for 30s at 94°C; annealing for 40s at primer-specific annealing temperature, extension for 45s at 72°C for 30 cycles). PCR products were resolved on 1.5% agarose gel. The gene-specific forward (F) and reverse (R) primer pairs were as follows:
Nurr1: AGTACCTTTATGGACAACTACAGCA (F), CGTAGTGGCCACGTAGTTCTGGT (R)
PTX3: CTCTCTGAAGAAGAAGCAGCA (F), CCGA GGGCACCATGGAGGCAAC (R)
TH: TCCTGCACTCCCTGTCAGAG (F), CCAAGA GCAGCCCATCAAAGG (R)
DAT: CAGAGAGGTGGAGCTCATC (F), GGCAGA TCTTCCAGACACC (R)
Calbinidin: GCAGTCATCTCTGATCACAGC (F), G AGGTCTGTGTACTCTGCTAG (R)
L-AADC: CCTACTGGCTGCTCGGACTAA (F), GC GTACCAGGGACTCAAACTC (R)
β-actin: GTGGGGCGCCCCAGGCACCA (F), CTC CTTAATGTCACGCACGATTTC (R)
Electrophysiological analysis
Electrophysiological analysis was carried out as previously described (10). Briefly, cells were plated on cover-slips, placed in a chamber, and perfused on the stage of an upright, fixed-stage Olympus (BHWI) microscope with an oxygenated solution containing NaCl, 140 mM; KCl, 5 mM; CaCl2, 2 mM; MgCl2, 1 mM; HEPES, 10 mM; glucose, 10 mM (pH 7.4). Experiments were performed at room temperature. For whole-cell recording, patch pipettes were pulled on a vertical puller (model PB-7; Narishige, Tokyo, Japan) from borosilicate glass pipettes and had tips of 1~2 μl outer diameter with tip resistances of 6~12 MΩ. Pipettes were filled with a bathing solution containing KCH3SO4, 98 mM; KCl, 44 mM; NaCl, 3 mM; HEPES, 5 mM; EGTA, 3 mM; MgCl2, 3 mM; CaCl2, 1 mM; glucose, 2 mM; Mg-adenosine triphosphate (ATP)6, 1 mM; guanosine triphosphate (GTP), 1 mM; and reduced glutathione, 1 mM (pH 7.2). Cells were voltage clamped and voltage dependent currents were measured with 20 mV steps (150 ms).
Reverse phase HPLC
Dopamine and its metabolite, dihydroxyphenylacetic acid (DOPAC), were analyzed by reverse-phase HPLC with electrochemical detection (12). Media from induced and un-induced cell culture (100 μl) were removed, placed in eppendorf microcentrifuge tubes, and 100 μl of 0.1 M perchloric acid was added to the samples. All samples were vortexed for 30 sec. and centrifuged at 14,000 RPM at 4°C. A total of 100 μl was withdrawn and placed into auto-sampler vials with spring glass inserts. Duplicate or triplicate 20 μl aliquots were injected for HPLC-EC analysis. An HR-80 catecholamine column (ESA, Bedford, MA) was used. The electrochemical detector was a Coulochem III (ESA, Inc.) that was set at 200 mV-guard cell potential and −150 and 350 mV-analytical cell potentials. Mobile phase consisted of 90% 50 mM sodium phosphate/0.2 mM EDTA/1.2 mM heptanesulfonic acid (pH=3.5) solution and 10% methanol mM. The analytical equipment included a pump running at 0.9 ml/min, refrigerated autosampler set at 4°C, and a mixing and degassing unit (all from Shimadzu, Inc., Columbia, MD). Peak area were integrated using chromatographic software (EZStart, Shimadzu, Inc.) and compared to a known standard curve of neurotransmitters. Neurotransmitter levels are reported as pg/ml.
Generation of hemi-Parkinsonian animals and rotation tests
Adult male Sprague-Dawley rats purchased from SASCO were anesthetized using a combination of Ketamine (5 mg/kg) and Xylazine (40 mg/kg) and were then placed in a Kopf stereotaxic frame (Tujunga, California) with the incisor bar set at 33 mm above the interaural line. Unilateral lesions of the left nigrostriatal dopaminergic pathway were produced by injection of 4 mg of 6-hydroxy dopamine (6-OHDA)9, dissolved in 1x PBS containing 0.1% ascorbic acid (2 μl), into the substantia nigra according to the following coordinates: 2.2 mm anterior to bregma, 1.5 mm lateral (right) of bregma and 7.9 mm ventral to the dura. The control group of animals were received an injection of 2 μl of 0.1% ascorbic acid in PBS. All injections were made at a rate of 0.5 μl/min and the needle remained in position for a further 3 minutes to prevent reflux of the toxin along the injection tract. The animals were allowed to recover and monitored daily until pre-operative weights were restored. Animals received an IP injection of 1 mg/ml apomorphine (300 mg/kg body weight, Sigma) and kept in 60 cm diameter plastic container. Animals’ behavior was observed for 30 minutes and rotations (over 360°), contralateral to the neurotoxin-damaged striatum, were recorded.
Transplantation
Neural progenitors derived from the limbus of GFP rats were subjected to dopaminergic differentiation as described above. Following the completion of induction program and confirmation of dopaminergic differentiation on a parallel set of cultured cells by RT-PCR 105 cells in 2 μl of PBS were transplanted into the striatum of each hemi-Parkinsonian rats (0 mm anteroposterior, 3 mm mediolateral and 5 mm dorsoventral of bregma) and the meninges (n=10). 2 μl PBS were injected into sham animals (n=10). All animals were subjected to rotations tests once a week after transplantation until they were sacrificed 4 weeks later. Animals (n=4) were examined 4 and 6 weeks post transplantation for tumor formation. The engrafted brains were fixed in 4% paraformaldehyde, sectioned, and subjected to immunohistochemical analysis.
Statistical Analysis
Values are expressed as mean fold change±SEM from three different experiments. Statistical analysis was done using Student’s t-test to determine the significance between different conditions.
Go to :
Results
First, we determined the fidelity of neuronal differentiation of limbus-derived neural progenitors. We cultured cell dissociates from the limbus in the presence FGF2 and Noggin for 5 days, which led to the generation of clonal neurospheres (Fig. 1A, B). Examination of the cellular phenotype of neurospheres revealed that they contained cells, which had incorporated BrdU and expressed neuroectodermal stem cell marker, Nestin (Fig. 1B). This observation suggested that the proliferating limbal cells had acquired neural progenitor properties in reduced BMP signaling conditions in vitro. To test whether or not limbus-derived neural progenitors in neurospheres can differentiate along neuronal lineage, we cultured neurospheres on poly-D-lysine and laminin coated glass cover slips in the presence of embryonic hippocampal cells across a semipermeable membrane in the absence of FGF2 for 5 days (Fig. 1A). Neurospheres spread within 48 hours in culture, and cells therein elaborated processes. Immunocytochemical analysis of the cells at the end of five days in culture revealed the expression of early and mature neuronal markers, βIII tubulin and respectively, suggesting their differentiation into generic neurons (Fig. 1D). To determine whether or not the differentiated cells had functional attributes of neurons we carried out whole cell patch recordings of cells with neuronal morphology (Fig. 1E and F). When voltage dependent currents were measured by a series of voltage steps (150 ms; −100 mV to +30 mV), applied from a steady holding potential of −70 mV, cells displayed rapidly activating inward currents above −60 mV, likely due to the voltage-gated sodium channels. Delayed but sustained outward currents were observed above −30 mV, characteristic of outwardly rectifying potassium channels. These observations suggested that cells differentiated from limbus-derived neural progenitors possessed electrophysiological properties characteristic of functional neurons.
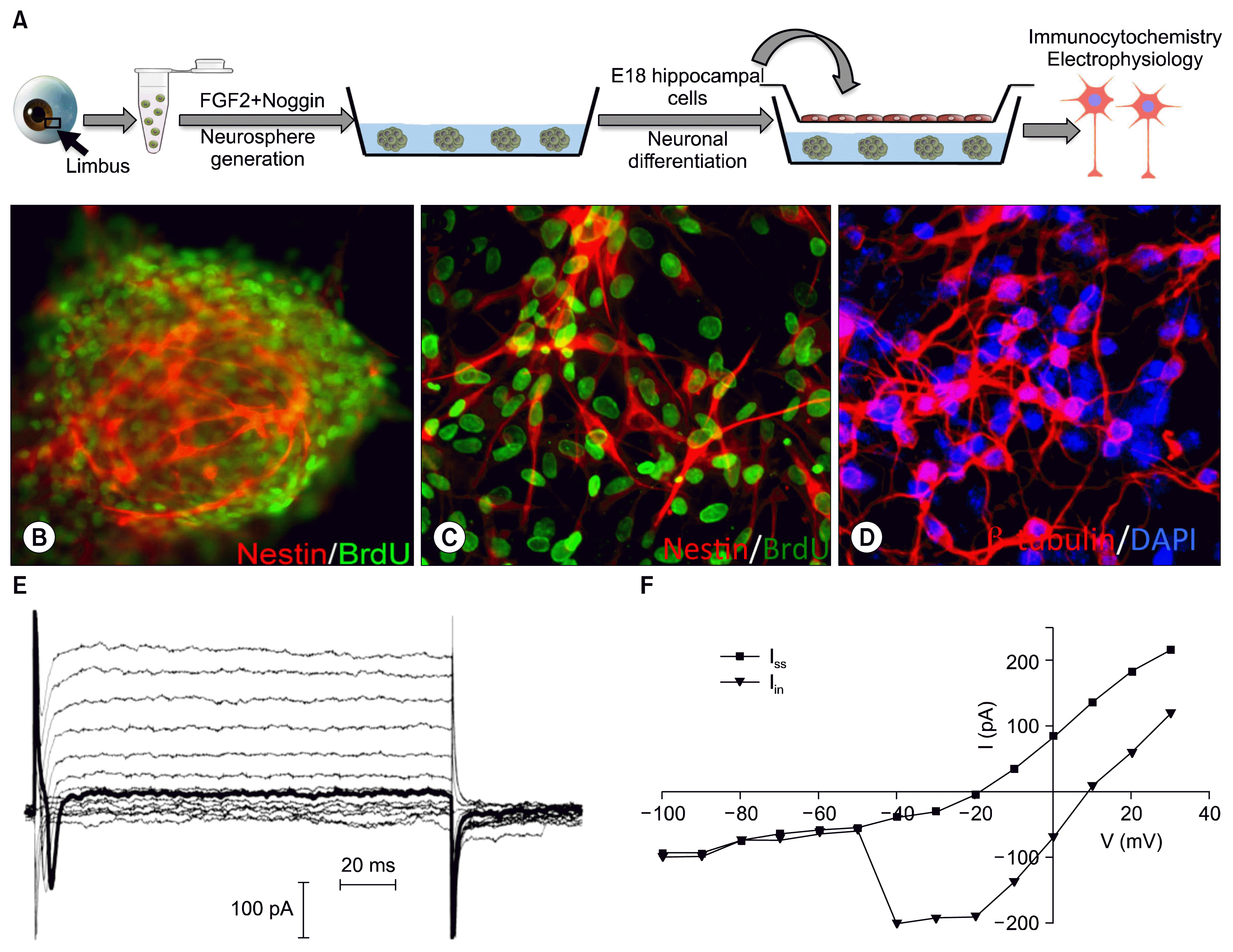 | Fig. 1Generation and differentiation of limbus-derived neural progenitors. A schematic representation of the differentiation protocol is provided in A. Cell dissociates from limbus when cultured in the presence of FGF2 and Noggin generated neurospheres with BrdU+ and Nestin+ cells (B; C, dissociated neurospheres). BrdU-tagged neurospheres, when co-cultured with E18 hippocampal cells spread out and cells acquired neuronal morphology, co-expressing neuronal markers β-tubulin/DAPI (D). Whole cell voltage clamped recordings of differentiated cells with neuronal morphology revealed fast inward currents and sustained outward currents (E). The current-voltage (I~V) curve (F) exhibited a typical I-V relationship of fast inward currents due to the voltage gated Na+ channels and sustained outward currents due to the outwardly rectifying K+ channels.
|
Next, we examined whether or not limbus-derived neural progenitors could differentiate into mesencephalic dopaminergic neurons. We subjected the BrdU-tagged limbus-derived neurospheres to a well-established method developed for the directed differentiation of ES cells into dopaminergic neurons, where the efficiency of differentiation is significantly increased in the presence of Shh and FGF8 (Fig. 2A) (13). We observed a progressive change in the morphology of cells in the neurospheres; by stage III, cells that were round at stage I, had elaborated processes and acquired the morphology of typical neurons (Fig. 2B~E). The stage III cells were examined for the expression of molecular and biochemical properties of dopaminergic neurons. Q-PCR analysis revealed that both un-induced and induced cells expressed transcripts corresponding to Nurr1, however, the levels were higher in the former, compared to the latter (Fig. 3A). The accentuation of Nurr1 expression following induction suggested the recruitment of intrinsic regulatory mechanism for dopaminergic differentiation. Pursuant to this notion Q-PCR analysis revealed the induction of Pitx3, a regulator downstream to Nurr1 in the regulatory hierarchy of dopaminergic differentiation (14, 15). The induction in the expression of Nurr1 and Pitx3 was accompanied by the onset of the expression of transcripts corresponding to calbindin, a marker of dopaminergic neurons, tyrosine hydroxylase (TH) and L-aromatic amino decarboxylase (L-AADC), enzymes for the biosynthesis of dopamine and its transporter, dopamine transporter (DAT). These transcripts were undetectable in the un-induced cells. Next, to corroborate dopaminergic differentiation at the biochemical levels, we carried out immunocytochemical analysis on stage III cells that were derived from BrdU-tagged cells in the neurospheres. BrdU-positive cells were observed expressing immunoreactivities corresponding to TH and mesencephalic transcription factor, En1 demonstrating that limbus-derived proliferating neural progenitors were induced along the dopaminergic lineage (Fig. 3B~F). Together these observations suggested that limbus-derived neural progenitors acquired the molecular and biochemical phenotypes of mesencephalic dopaminergic neurons non-cell autonomously under the directed differentiation protocol.
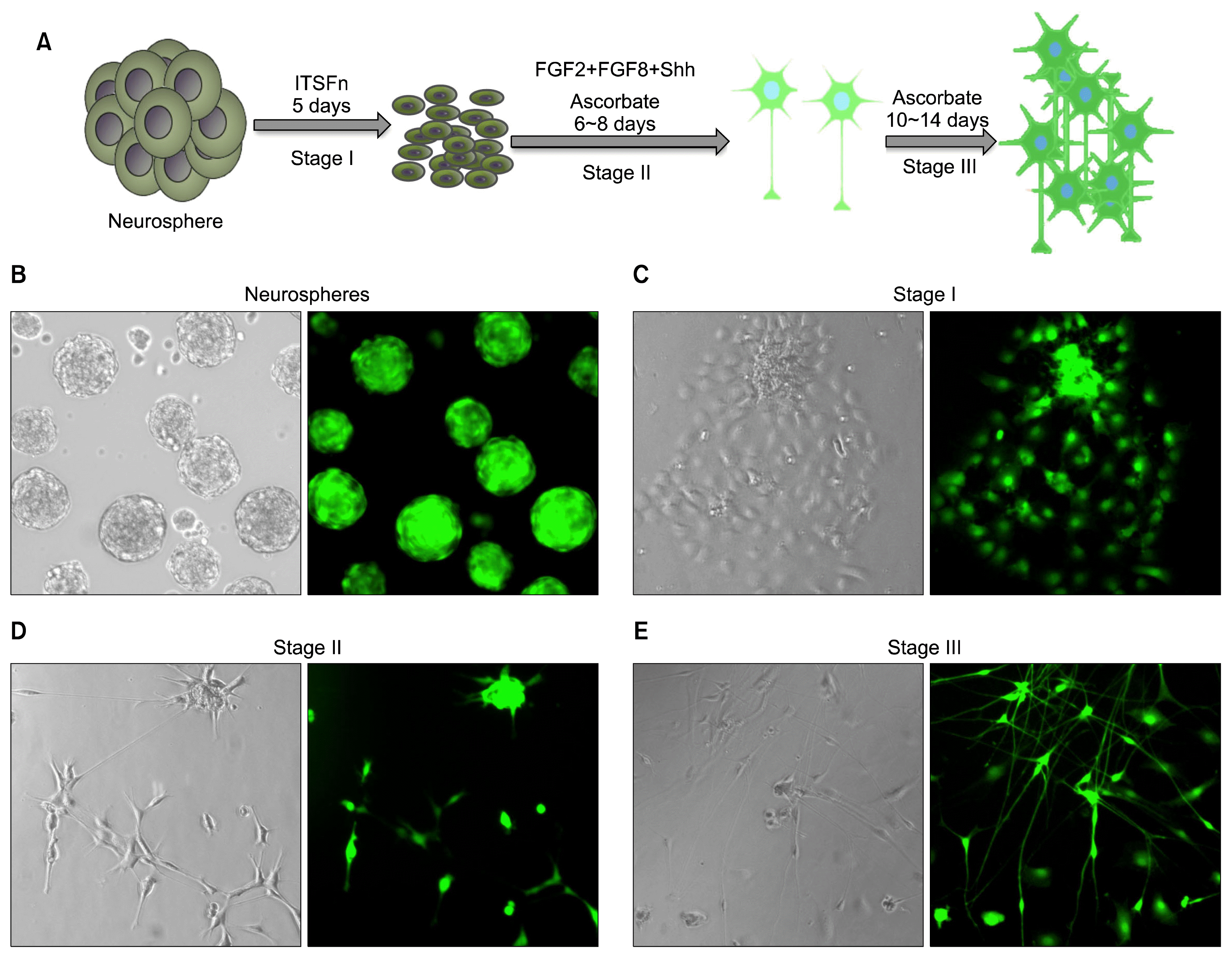 | Fig. 2Differentiation of limbus-derived neural progenitors along the dopaminergic cell lineage. Limbus-derived neurospheres were subjected to three stages directed dopaminergic neuron differentiation protocol, using Shh and FGF8 (A). Cell dissociates from GFP-rat limbus when cultured in the presence of FGF2 and Noggin generated neurospheres with GFP+ neural progenitors. Neurospheres spread under the influence of ITSFn (Stage1) and cells therein acquired neuronal morphology under the influence of Shh and FGF8 (Stage II) and increased in number with ascorbate (Stage III).
|
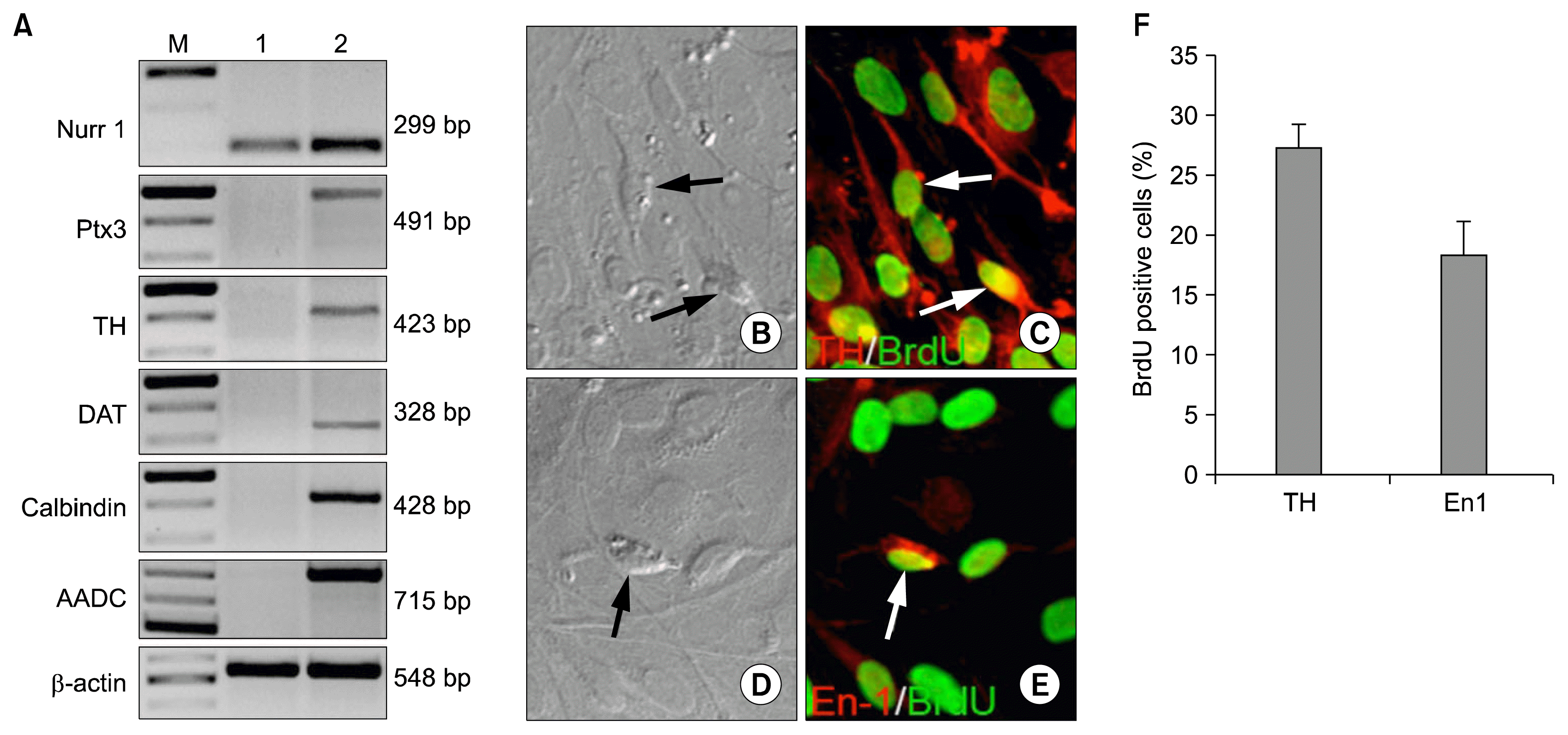 | Fig. 3Molecular and biochemical properties of limbus-derived dopaminergic neurons. RT-PCR analysis of cells in neurospheres (lane 1) and cells at the end of Stage III of differentiation (lane 2) demonstrated the induction of transcripts corresponding to dopaminergic neuron regulator (Nurr1 and Ptx3) and phenotypic and functional markers (Calbindin, TH, DAT, and AADC, compared to the former (A). Immunocytochemical analysis of BrdU-tagged Stage III cells revealed BrdU+ cells, expressing dopaminergic neuron markers, TH (B, C, F) and En1 (D, E, F).
|
Next, we examined the functional attributes of limbus-derived neural progenitors differentiated along the dopaminergic lineage (Fig. 4A). First, we examined the ability of the induced cells to synthesize and evoke-release dopamine in the culture medium. Reverse-phase high performance liquid chromatography (HPLC) did not detect dopamine in the culture medium of un-induced cells. However, exposure of induced cells to 56 mM KCL, evoked a release of DA and its metabolite DOPAC in the culture medium to a mean concentration of 250 pg and 1650 pg, respectively (Fig. 4B). Second, to test whether or not limbus-derived dopaminergic neurons could integrate functionally in vivo we transplanted GFP expressing stage III cells into the striatum of 6-OHDA-lesioned rats, a functional and behavioral model for Parkinson’s disease (16). Behavioral tests of these animals revealed that the transplantation of induced cells led to a decrease in the amphetamine-induced contralateral rotations near to the levels of PBS non-lesioned controls for two weeks after the surgery. Retrospective immunohistochemical analysis of brains undergone transplantation revealed the presence of GFP positive cells in the host’s striatum, expressing TH, suggesting the survival and integration of grafted cells at the site of the lesion. Examination of brains, four and six weeks after the transplantation, did not reveal the presence of tumor. Together, these observations suggested that limbus-derived neural progenitors were capable of generating differentiated neurons with biochemical and functional attributes of mesencephalic dopaminergic cells.
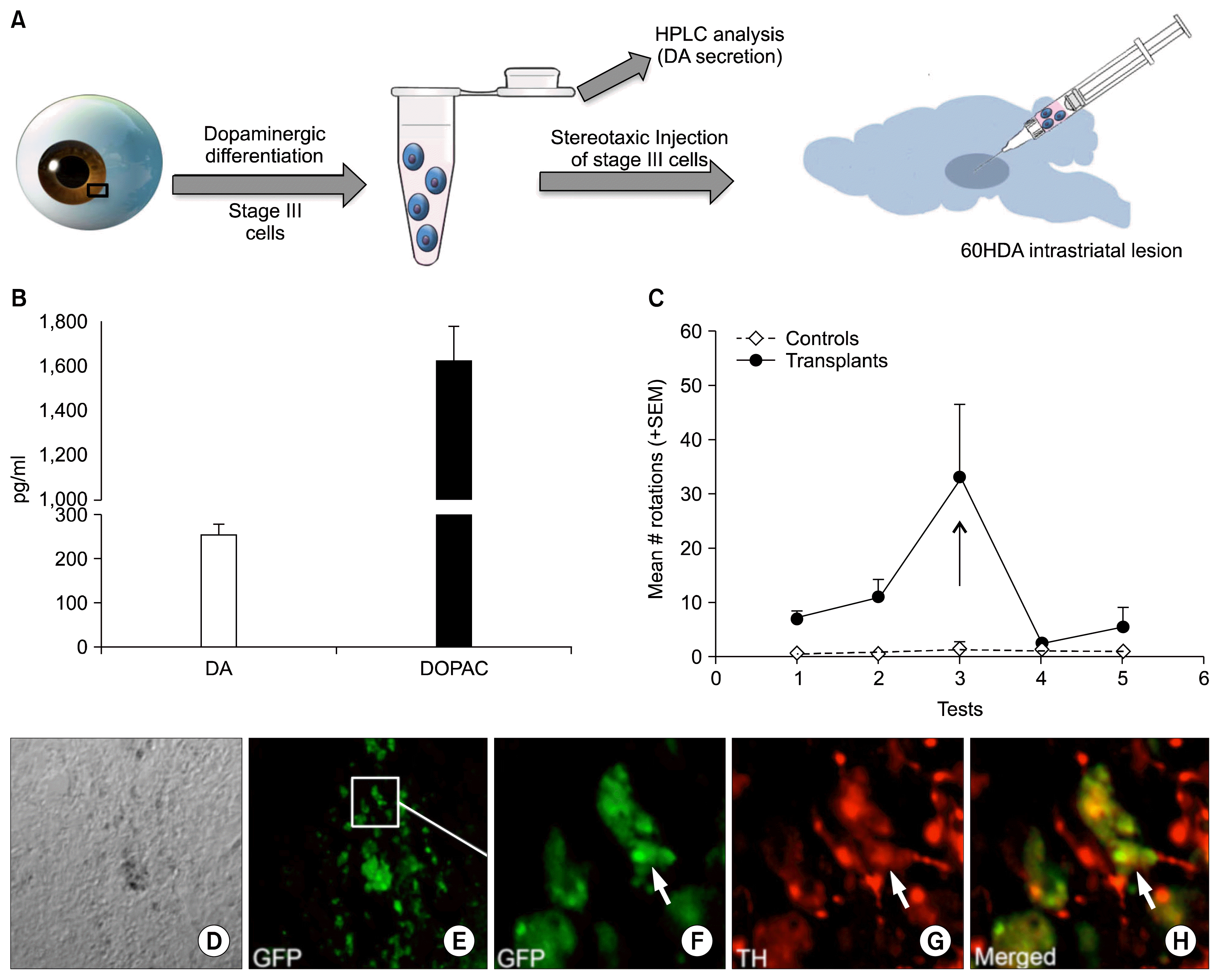 | Fig. 4Functional analysis of limbus-derived dopaminergic neurons. The schematic representation of the functional analysis of non-cell autonomously derived dopaminergic neurons is given in A. the HPLC-EC analysis revealed that limbus-derived Stage III cells release DA and DOPAC in the culture medium in response to KCL (B). Un-induced cells were negative for DA and DOPAC. Transplantation of limbus-derived GFP+ stage III cells in the striatum of 6OHDA-lesioned rats reduced the amphetamine-induced rotations to sham-lesioned controls one-week post-translation (C). Retrospective immunohistochemical analysis of sections through the striatum of transplanted rats revealed GFP+ limbus-derived stage III cells expressing TH in the host’s striatum (D~H).
|
Go to :
Discussion
We have recently demonstrated that cells from the regenerative tissue limbus (8) could be reprogrammed to pluripotency by both nucleic acid and non nucleic acid method (17). However, the acquisition of pluripotency by somatic cells lend themselves the risk of generating teratomas, a serious barrier to their practical clinical use. Here, we have tested the hypothesis that limbus-derived neural progenitors can be directly differentiated non-cell autonomously into midbrain dopaminergic neurons, without an intervening pluripotent stage. This hypothesis is based on our observations that the limbus-derived neural progenitors express Nurr1, a nuclear orphan receptor that plays an important role in the development of midbrain dopaminergic neurons. For example, it has been observed that dopaminergic neurons are depleted in mice lacking Nurr1 (11) and that Nurr1 can induce dopaminergic differentiation in progenitors from different developing CNS regions (18). The limbus-derived neural progenitors responded to culture conditions defined by the presence of Shh and FGF8 for the directed differentiation by changing morphologically and biochemically; at the end of stage III of the differentiation protocol, these cells had acquired neuronal morphology with long processes and up regulated the expression of Nurr1. However, the expression of Nurr1 does not conclusively identify dopaminergic neurons, as it is found in areas in the developing midbrain beyond dopaminergic neurons (14). Therefore, the acquisition of the expression of Pitx3, whose expression follows that of Nurr1 in vivo and confined to midbrain dopaminergic neurons suggested that the differentiation of limbus-derived neural progenitors along the midbrain dopaminergic lineage was likely acquired through the recruitment of normal mechanism, thus predicting the stability of the differentiated phenotype (15). The differentiated phenotype of induced limbus-derived neural progenitors betrayed mature and functional dopaminergic neurons. The induced cells expressed immunoreactivities corresponding to En1, an early regulator of dopaminergic neuron cell fate, whose expression is continued in matured cells (15, 19) and the calcium binding protein, Calbindin, which is expressed in a large subset of dopaminergic neurons in the pars compacta of the substantia nigra (20, 21). More importantly, from the functional viewpoint, the induced cells expressed transcripts corresponding to enzymes (TH and L-AADC) essential for dopamine biosynthesis. That the induced cells can synthesize and release dopamine and partially ameliorate motor dysfunction in the rat model of hemi-parkinsonism suggested that these cells had acquired the functional attributes of midbrain dopaminergic neurons.
Taken together, our observations suggest that limbus-derived neural progenitors represent a potential source of dopaminergic neurons along with ES cells, iPS cells, and MSCs for stem cell approach to Parkinson’s disease (1, 2, 3, 5). Like MSCs, it addresses the issue of teratoma formation associated with the pluripotent nature of both ES and iPS cells. These cells may be relatively safe than MSCs, whose efficacious differentiation along the dopaminergic lineage necessitates the expression of exogenous genes. For example, the efficiency of dopaminergic differentiation of MSCs was increased to 40% when they were genetically engineered to constitutively express Notch intracellular domain (NICD) followed by induction mediated by growth factors (6). Though dopaminergic neurons derived from NICD expressing MSCs were successful in restoring motor functions in animal models of Parkinson’s disease the risk of insertional mutagenesis remains a concern, in addition to the fact that the constitutive activation of Notch signaling may be detrimental, as it may lead to undesirable cell proliferation (22). Therefore, a facile non-cell autonomous method presented here, consisting of defined culture conditions for directed differentiation where normal mechanism of dopaminergic differentiation is recapitulated, regardless of the cell source, would constitute a relatively safe stem cell approach to address the progressive loss of the dopaminergic neurons to Parkinson’s disease. The cells in the limbus, which are accessible and expandable in vitro, are amenable to these conditions for testing the proof of principle of autologous stem cell approach to repair the nigrostriatal degeneration in Parkinson’s disease.
Go to :
 XML Download
XML Download