

 PDF
PDF Citation
Citation Print
Print


Introduction
The aim of regenerative medicine is currently underway to use the body’s own cells to repair diseased or damaged tissue. The use of stem cells from different tissues for regenerative medicine applications has increased over the past few years. Amniotic fluid (AF) represents rich sources of stem cells population deriving from either the fetus or the surrounding amniotic membrane that can be used for clinical therapeutic applications in the patients who develop organ failure that have resistance to current therapies. The AF was first studied at the beginning of the 20th century (1). Initial studies have been performed in order to detect fetal abnormalities during development the fetus (2). Isolation and identification of amniotic fluid- stem cells (AFSCs) dating back to the early 1990’s (3). The study of AFSCs has received significant attention of late for several reasons. First, AF is easily collected during the first trimester of pregnancy by scheduled amniocenteses for fetal karyotyping, prenatal diagnosis and detection a variety of genetic diseases (4). Second, expansion and storage of AFSCs is easy and achieved at minimal costs. Finally, these cells could be stored in cell banks and used in disease research, drug screening and genetic disorders.
Go to : 
Amniotic Fluid
Human AF is a protective and nourishing watery liquid that providing mechanical support during embryogenesis and is constituted of about 98% water (4). Other ingredients include electrolytes, pigments, sugars, fats, amino acids, proteins, carbohydrates, enzymes, growth factors and cells (5). This volume and composition varies throughout pregnancy (5, 6). After formation of the amniotic cavity, 7~8 days after fertilization, this fluid starts to gather immediately. AF volume increases progressively and is then completely surrounding the embryo after 4 weeks of pregnancy. The average volume is 270 ml at week 16 which increases to 400 ml at week 20 of pregnancy and 800 ml at birth (5). At the beginning of pregnancy, the amniotic osmolarity is similar to the fetal plasma. After keratinization of the fetal skin, which usually occurs at week 24 of pregnancy, amniotic osmolarity decreases relatively to maternal or fetal plasma, mainly due to the inflow of fetal urine (7). Additional investigations have been recently focused on the cellular and molecular properties of amniotic derived cells and their potential use in pre-clinical models and in cell therapies (4).
Go to :
Isolation and culturing of AFSCs
There are three major protocols for isolation of AFSCs from human amniotic fluid. The first one is based on single-stage method (8). In this method AF collected from second-trimester amniocentesis is centrifuged. The number of cells is counted by hemocytometer and mixed with an equal volume of culture medium, usually Dulbecco’s Modified Eagle Medium (DMEM) supplemented with Fetal Bovine Serum (FBS) and the cells allowed to adhere to a plastic culture plate at 104 cells/cm2 and incubated overnight at 37°C under 5% CO2 (9). Culture medium is changed after 3~5 days to remove non adherent cells and twice weekly thereafter. The primary cells are cultured for 4~5 days until they reached confluence and are defined as passage “0”. In this step the heterogeneous morphological cell population appears and after several sub-culturing the fibroblastic like cells was dominated. The cells typically reach confluence in 4 to 6 days and the remaining cells are cryopreserved in cryopreservation media (10% dimethylsulfoxide, 90% FBS), frozen at −80°C for 24 h, and stored in liquid nitrogen the next day. The second one is immunoselection based on surface antigens. Ditadi was the first researcher to show that the c-Kit population cells extracted from the AF do have hematopoietic potential (10). At this time, Atala et al. (2) and Schmidt et al. (11) isolated CD117 positive cells by c-kit (a rabbit polyclonal antibody to CD117) and CD133 positive cells by CD133 magnetic beads, respectively. Their study showed these cells can be easily expanded in cultures and sub-population of CD133 positive exhibited similar characteristics of mesenchymal progenitors cells (11). Following these studies, Arnhold et al. sorted CD117 positive cells by magnetic associated cell sorting. They indicated that the percentage of CD117 positive cells was 3.2±1.03% of the whole cell population and demonstrated that these cells could differentiate to adipogenic, osteogenic, myogenic and neurogenic lineage (12). The third one is the two-stage culture protocol established by Tsai et al., using nonadhering AF cells of the primary amniocytes culture to isolate AFSCs. In this protocol, nonadhering AF cells are collected from supernatant of AFCs that cultured in serum-free changes medium (first stage). Then collected cells are plated for AFSCs culturing after the completion of fetal chromosome analysis (second stage) (13). This method has some advantages over the others. Major advantage of this culture protocol comparing to the other two is that instead of the adhering cells derived in AF it isolates from the nonadhering cells, which is being left in the incubator without any added nutrition for 7~10 days (13). As mentioned two-stage method is more superior compared to other methods, the use of this method have been proposed.
Go to :
Characterization and application of AFSCs
The AFSCs are mainly composed of three heterogeneous groups of adherent cells, calcified based on their growth, morphological and biochemical characteristics that derived from the three germ layers (14). Epithelioid (E-type) cells that are cuboidal to columnar, derived from the fetal skin and urine, AF (AF-type) cells are originating from fetal membranes, and fibroblastic (F-type) cells are generated mainly from fibrous connective tissue. The percentage of these cells is 33.7%, 60.8% and 5.5%. Some studies have reported that AFSCs can be easily obtained from a small amount of AF (4, 15). Like other mesenchymal stem cells (MSCs), the AFSCs expressed CD73, CD90, CD105, CD29, CD166, CD49e, CD58, CD44 and HLA-ABC antigens and are negative for the hematopoietic markers such as CD14, CD34, CD45 and CD133, the endothelial marker such as CD31, and the HLA-DR antigen (Fig. 1) (4). These cells are able to differentiate along adipogenic, osteogenic, myogenic, endothelial, neurogenic and hepatic pathways (Table 1) (16–19). Additionally, the majority of these cells expressed the pluripotency markers such as the octamer binding protein 3/4 (Oct-3/4), the homebox transcription factor Nanog, and the stage-specific embryonic antigen 4 (SSEA-4) (20, 21). Similar to MSCs, AFSCs express MHC II at a very low level. Unlike human MSCs that are telomerase negative, low to moderate levels of the enzyme have been described in AFSCs (5, 22). Telomerase is an enzyme that maintains telomere sequences at chromosomal ends (23). Telomeres consist of TTAGGG repeats protect the ends of chromosomes from end-to-end fusion, recombination and deterioration (5). The presence of telomerase activity in both cultured and uncultured cells was found in 1999 (24). Another interesting finding have shown the presence of a population of Oct-4-positive cells in AF (25). AFSCs also express vimentin and alkaline phosphatase, which are markers of pluripotent stem cells (5, 26). Like other MSCs, AFSCs are attractive candidates for clinical applications, which were reviewed in Table 2. For instance some reports indicated that the AF can be a reliable and practical source of cells for the engineering of select fetal tissue constructs (27, 28). Another clinical application of AFSCs is the use as produce mineralized bioengineered constructs in vivo, functional repair of bone defects and bone engineering (2, 29–31), Neural tissue regeneration and nerve myelination (32–35), lung epithelial regeneration (36, 37), cardiac regeneration (38, 39) and kidney regeneration (21).
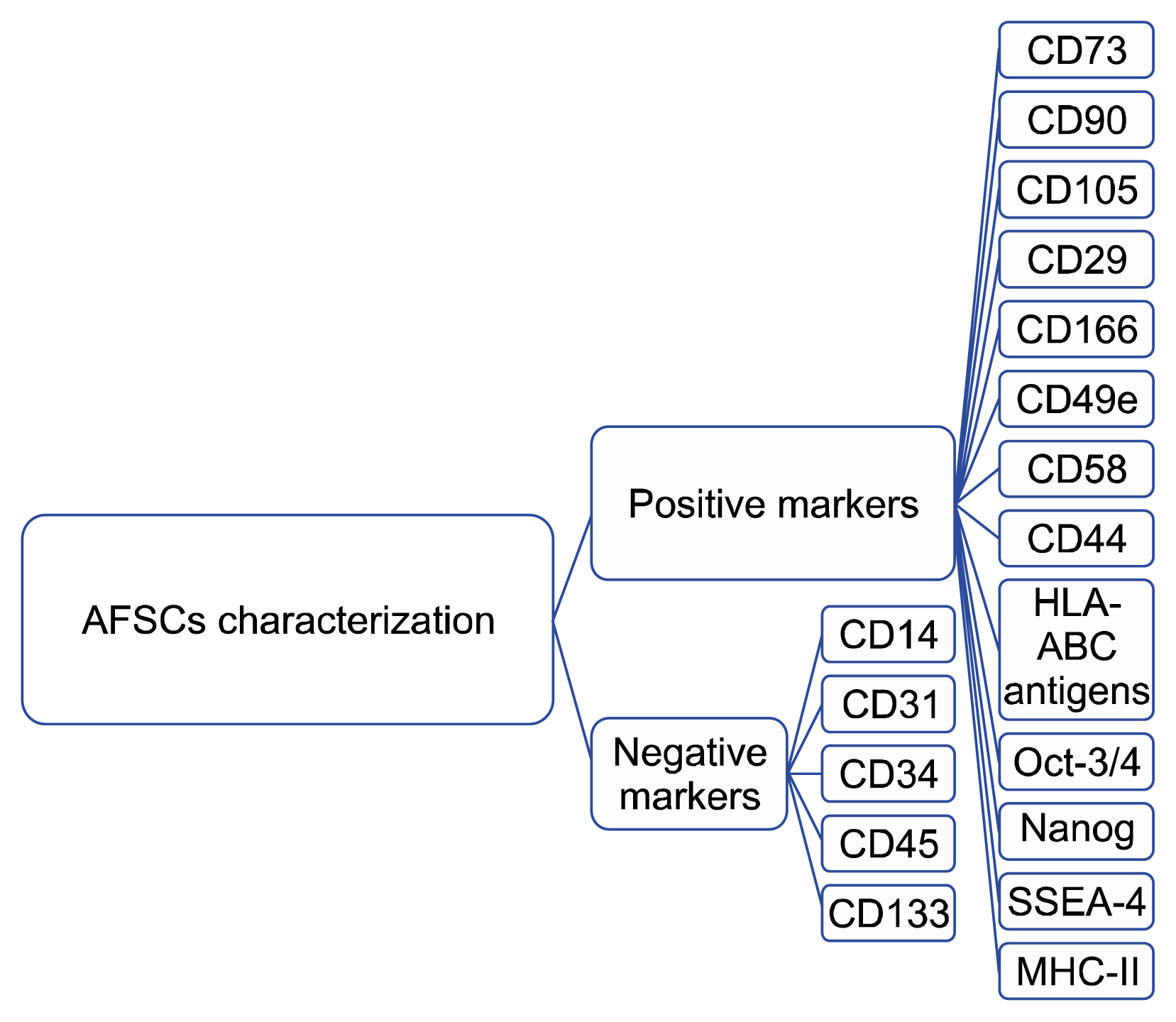
Table 1
Some researches on differentiation potential of AFSCS
| Author names | Year of publication | Title of publication | Type of differentiation | Main results |
|---|---|---|---|---|
| McLaughlin et al. | 2006 | Stable expression of a neuronal dopaminergic progenitor phenotype in cell lines derived from human AFSCs | Neural dupaminergic differentiation | They reported that AFSCs1 are primarily composed of a population of progenitors with a phenotype similar to that of committed dopaminergic neurons (18) |
| Perin et al. | 2007 | Renal differentiation of AFSCs | Renal differentiation | AFSCs may represent a potentially limitless source of ethically neutral, unmodified pluripotential cells for kidney regeneration (19) |
| Carraro et al. | 2008 | Human AFSCs can integrate and differentiate into epithelial lung lineages | Epithelial lung lineages differentiation | Human AFSCs can undergo lung-specific line-age differentiation and that these cells possess a certain level of plasticity in response to different types of lung damage (36) |
| Donaldson et al. | 2009 | Human AFSCs do not differentiate into dopamine neurons in vitro or after transplantation in vivo | Neural dupaminergic differentiation | AFSCs express specific markers of neural progenitors and immature dopamine neurons, but were unable to fully differentiate into dopamine neurons in vitro or in vivo (17) |
| Ditadi et al. | 2010 | Human and murine AF2cKit+Lin-cells display hematopoietic activity | Erythroid, myeloid, and lymphoid lineages | Under appropriate differentiation conditions, AFSCs were able to generate all the blood lineages (myeloid, erythroid and lymphoid colonies) (10) |
| Hauser et al. | 2010 | Stem cells derived from human AF contribute to acute kidney injury recovery | Renal differentiation | They reported that hAFSCs may provide an alternative source of stem cells for the treatment of acute kidney injury (16) |
| Peister et al. | 2011 | Cell sourcing for bone tissue engineering: AFSCs have a delayed, robust differentiation compared to MSCs | Osteogenic differentiation | Stem cell source can dramatically influence the magnitude and rate of osteogenic differentiation in vitro (27) |
![]()
Table 2
Some researches on application of AFSCs
| Author names | Year of publication | Title of publication | Application | Main results |
|---|---|---|---|---|
| Kaviani et al. | 2001 | The AF3 as a source of cells for fetal tissue engineering | Fetal tissue reconstruction | They seeded subpopulation of MSCs4 from the AF onto a polyglycolic acid polymer/poly-4-hydroxybutyrate scaffold and reported that these cells were able to attach firmly to the scaffolds and form confluent layers with no evidence of cell (23) |
| Kunisaki et al. | 2006 | Fetal cartilage engineering from amniotic mesenchymal progenitor cells | Tissue engineering | AF could be a good cell source for tissue engineered diaphragmatic reconstruction (24) |
| De Coppi et al. | 2007 | Isolation of amniotic stem cell lines with potential for therapy | Bone mineralization | Implantation of AFSCs5 into an immunodeficient mouse cause to production of mineralized tissue in vivo (2) |
| Cipriani et al. | 2007 | Mesenchymal cells from human amniotic fluid survive and migrate after transplantation into adult rat brain | Regeneration of neural tissue | Cipriani et al. noticed AFSCs grafted cells tended to migrate towards injured brain regions and differentiated into neurons. They suggested the amniotic fluid could be an alternative source for MSCs (28) |
| Pan et al. | 2007 | Post-injury regeneration in rat sciatic nerve facilitated by neurotrophic factors secreted by AF MSCs | Regeneration sciatic nerve | AFSCs could increase nerve degeneration due to the neurotrophic factors secretion (29) |
| Carraro et al. | 2008 | Human AFSCs can integrate and differentiate into epithelial lung lineages | Lung epithelial regeneration | AFSCs transplantation into an injured lung cause to pulmonary lineage differentiation (32) |
| Chenge et al. | 2010 | Enhancement of regeneration with glia cell line-derived neurotrophic factor-transduced human AF MSCs after sciatic nerve crush injury | Peripheral nerve regeneration | They embedded AFSCs and glial cells in matrigel and transplanted in to the injured sciatic nerve of rat and indicated that AFSCs promoted nerve regeneration (30) |
| Pan et al. | 2009 | Combination of G-CSF6 administration and human AF MSCs transplantation promotes peripheral nerve regeneration | Peripheral nerve regeneration | They embedded AFSCs in fibrin glue and delivered to the injured sciatic nerve. Increased nerve myelination and improved motor function were observed in AFS transplanted (31) |
| Yeh et al. | 2010 | Cellular cardiomyoplasty with human AFSCs: in vitro and in vivo studies | Cellular car diomyoplasty | AFSCs induce angiogenesis at the injured site, have cardiomyogenic potential, and may be used as a new cell source for cellular cardiomyoplasty (34) |
| Yeh et al. | 2010 | Cardiac repair with injectable cell sheet fragments of human AFSCs in an immune-suppressed rat model | Cardiac regeneration | Transplantation of AFSCs sheet fragments stimulated a significant increase in vascular density, improved wall thickness and a reduction in the infarct size (35) |
| Peterson et al. | 2010 | Tissue-engineered lungs for in vivo implantation | Lung regeneration | The results suggested that repopulation of lung matrix is a viable strategy for lung regeneration (33) |
| Perin et al. | 2010 | Protective effect of human AFSCs in an immunodeficient mouse model of acute tubular necrosis | Kidney regeneration | They found that injection of AFSCs into damaged kidney modulate the kidney immune milieu in renal failure (17) |
| Rosa et al. | 2010 | MSCs lead to bone differentiation when cocultured with dental pulp stem | Bone engineering | Combination of AFSCs with dental pulp stem cells may provide a rich source of soluble proteins useful for bone engineering purposes (25) |
| Maraldi et al. | 2011 | Human AFSCs seeded in fibroin scaffold produce in vivo mineralized matrix | Bone engineering | The results indicated the strong potential of AFSCs to produce mineralized bioengineered constructs in vivo (26) |
| Peister et al. | 2011 | Cell sourcing for bone tissue engineering: AFSCs have a delayed, robust differentiation compared to MSCs | Bone regeneration | They investigated the cells were cultured within the porous medical grade poly-epsiloncaprolactone (mPCL) scaffolds could differentiate to osteoblastic cells and concluded that the AFSCs were an effective source for functional repair of bone defects (27) |
![]()
Go to :
Conclusion
Like other MSCs, AFSCs have advantages such as rapid cell proliferation, low or negligible immunogenicity. Many of these cells seem to express some of the same pluripotency markers. All of these features make them valuable for potential therapy applications. Thus far they have been used in pre-clinical settings to treat a variety of diseases such as osteogenesis imperfecta, congenital diaphragmatic hernia, Parkinson’s disease and cancer with encouraging results. Finally, their usefulness for AFSCs is very likely to expand their future clinical use even further. Human AFSCs could be isolated by several methods including immunoselection method based on surface antigens, single-stage and two-stage methods. Two-stage method has some advantages over the others. For example this method doesn’t interfere with the normal culture process for fetal karyotyping and also illustrated their ability to successfully differentiate into osteocyte, adipocyte and etc in vitro. As two-stage method is more superior compared to other methods, the use of this method have been proposed.
Go to :
 XML Download
XML Download