

 PDF
PDF Citation
Citation Print
Print


Introduction
Unrelated umbilical cord blood (UCB) transplantation has become one of the standard therapeutic options for pediatric patients with hematological malignancies (1–6). The success of UCB transplantations has been demonstrated in more than 10,000 cases so far for various genetic, hematological or immunological disorders (7). Compared to unrelated bone marrow, UCB has the advantages of rapid availability (8) and relatively lower risk of causing severe acute graft-versus-host disease (GVHD) despite some donor-versus-recipient HLA disparity (1–5, 8, 9).
UCB transplantation, although successful in children, poses some challenges for adults. One of the major disadvantages of UCB is attributed to limited number of CD34+ HSCs resulting in higher graft failure rates compared to bone marrow transplantation (10). While the dosage that is deliberated in various conferences is in the range of 20∼30×106 total nucleated cells /kg body weight of the recipient, the average content of UCB grafts is in the range of 800∼4,000 million cells, most of them having counts below the median value (11). The next important challenge is to obtain a suitable HLA matched graft containing adequate cell dose of CD34+ HSCs. Use of multiple cord blood units is one of the recent approaches tried by few groups for increasing the CD34+ stem cell counts for transplantation into adult patients (8, 12). However this strategy also faces its own limitations (11). Hence, it is best to expand CD34+ stem cells of the desired HLA type to get the desirable cell numbers for transplantation (13, 14).
Ex vivo expansion of CD34+ cells, however also have its own attendant problems, mainly that of maintenance of stemness without allowing differentiation of CD34+ cells to the cells of hematopoietic lineage (15). These expanded cells could lose their stem cell potential, a requisite for successful transplantation and engraftment of HSCs. Bone marrow derived mesenchymal stem cells (BMMSCs) have been reported to be employed as feeders for expansion of HSCs in vitro (16, 17). However, using one’s own BMMSCs, though the best known choice so far, may not be always available due to several reasons. In addition, BMMSCs can be expanded to only a few passages. Hence it is imperative to look out for alternative sources of MSCs that can support ex vivo expansion of HSCs (18, 19).
In this study, we have analyzed the suitability of umbilical cord derived mesenchymal stem cells (UCMSCs) for fresh or stored CD34+ cell expansion and maintenance of their stem cell potential. We have also compared the results of using MSCs derived from the same cord as that of cord blood (autologous) and those derived from an unrelated cord (allogeneic). We observed that both autologous and allogeneic UCMSCs have equally supported the expansion of UCB HSCs in vitro. Expanded cells have been characterized and their functionality confirmed by traditional in vitro clonogenic assays. Expanded HSCs have been cryopreserved using human serum albumin and DMSO under optimal conditions. These findings have extreme significance in transplant settings and are likely to address a major unmet medical need in the sense that the expanded HSCs may alleviate the problem of graft failure due to insufficient cell number used for transplantation.
Go to : 
Materials and Methods
Collection of biological samples
The umbilical cord and cord blood units were collected immediately after the delivery as per the usual practice after the approval from the local ethics committee. Informed consents were obtained from the mothers. Bone marrow samples were collected from healthy volunteers with the approval of the institutional review board. The samples were sent to the laboratory in ambient temperatures (∼25°C) and were processed within 48 hours post collection.
Derivation of mesenchymal stem cells from human umbilical cord tissue
The MSCs were isolated from the cords using an ‘in-house’ established protocol (20). Briefly, the cord was cleaned with PBS (Invitrogen, USA) containing antibiotics. The blood clots were removed and the cord was dissected into smaller explants and placed on tissue culture dishes in expansion media {DMEM/F12 (Invitrogen, USA) with 10% serum (Hyclone, USA) and 2 ng/ml basic FGF (Sigma, USA)}. The cells were allowed to grow out from the explants, expanded in monolayer at 37°C and 5% CO2 and supplemented with fresh media every alternate day. The cells were routinely sub-cultured every 5∼6 days and characterized at every passage as described below. Umbilical cord derived mesenchymal stem cells (UCMSCs) from passage 2 to 6 were used for all the expansion experiments.
Derivation of mesenchymal stem cells from human bone marrow
The BMMSCs were isolated and expanded according to the previously reported methods (20, 21). Briefly, the bone marrow cells were washed and layered on ficoll (Sigma, USA) and centrifuged at 400g for 20 minutes. The mononuclear cells were collected from the interface and plated onto culture flasks in DMEM/F12 media with 10% serum and bFGF. MSCs grew as colonies at passage 0 (P0) initially and gradually formed a monolayer of cells. These cells were harvested on confluence and subcultured every 7∼10 days and characterized as described below.
Processing of umbilical cord blood
All the processes were carried out in our clean room facilities. The volume of the cord blood was determined and equal volume of hespan was added to the collection bags and mixed well. The collection bag was kept standing for half an hour after proper mixing of the cells with Hespan. The RBCs aggregated together due to hespan and increased in weight. RBCs settled down and were collected and discarded. The bag containing plasma and the stem cell fraction was then centrifuged at 1800 rpm for 10 minutes to pellet down all the nucleated cells. The plasma was expelled out using a plasma expressor. The final volume of the concentrated cells was adjusted to 20 ml. To this 20 ml of cell concentrate, 5 ml of freezing media was added and mixed well. The cells were suspended in the freezing medium and then transferred to the freezing bag. This was then frozen using a controlled rate freezer. Bags containing cryopreserved stem cells were transferred to the liquid nitrogen freezer for long term storage.
Isolation of mononuclear cells from UCB
From fresh cord blood: Cord blood was collected in 50 ml centrifuge tubes, and centrifuged at 1,500 rpm at 4°C. The supernatant was discarded. To the cell pellet, equal volume of 3% Dextran was added and mixed well following which the tubes were kept standing for half an hour. The contents were then mixed properly and allowed to stand further for next 30 minutes. The leukocyte rich plasma (LRP) was collected in fresh centrifuge tubes and centrifuged at 1,500 rpm for 5 minutes to pellet down all the nucleated cells. The cells were washed once with 1xPBS (Invitrogen, USA) and were then resuspended in 20 ml of 1xPBS.
From long term frozen processed cord blood: The MNCs were also isolated from the processed cord blood stored at liquid nitrogen temperatures in the cord blood repository for long term. Designated units were retrieved from liquid nitrogen, thawed at 37°C with intermittent shaking for uniform thawing. The contents were collected in the sterile 50 ml centrifuge tube. To this equal volume of thawing media (IMDM basal medium with DNase) was added and mixed well to prevent clumping of cells. The cells were washed and resuspended. They were counted using the cell counter (Beckman Coulter counter) and viable cell count was determined using Viaprobe on Flow cytometer (FACS Calibur, BD).
The cell suspension from either the fresh or thawed sample was layered on equal volume of Ficoll (Histopaque, Sigma, USA). The tubes were then centrifuged at 400 g for 20 minutes to obtain the mononuclear cells by density gradient centrifugation. The MNCs were collected in fresh centrifuge tubes and were washed once with the thawing medium. The viable cells were counted.
Sorting of CD34+ cells from MNCs
The CD34+ cells were sorted out from the MNCs using the sterile magnetic beads (Miltenyi Biotech, Germany) following the manufacturer’s protocol. Briefly, for every 100×106 cells, 100 μl of FcR blocking reagent was added to the MNC cell pellet and resuspended in 300 μl of MACS Buffer (1×PBS with 4% serum) and mixed well. Also 100 μl of CD34 microbeads was added to the cell suspension and the tube was incubated at 40C for 30∼40 minutes. Next, the cells were washed twice with 10 times the volume of the cell suspension. The cells were then re-suspended in 1 ml for MS columns (<2×108 cells) or in 3 ml for LS columns (2×108∼2×1010 cells) of MACS buffer depending on the cell count. This cell suspension was then passed through the column and the positive fraction of CD34+ cells was collected by depletion strategy in a fresh sterile centrifuge tube. To obtain a higher purity, the positive fraction cells were passed through a second column. The cells were then collected, counted and characterized before they are taken up for expansion.
Immunophenotyping
The sorted, cryopreserved and expanded CD34+ HSCs were characterized for cell surface marker expression. The cell purity was confirmed by labeling the sorted cells (50,000 cells) with PE labeled antiCD34 antibody (BD Pharmingen, USA). Along with this, the cells were also stained with FITC or PE labeled antibodies against CD31, CD133, CD33, CD38, CD44 VLA-4 and LFA-1 (CD11a) (all antibodies were obtained from BD Pharmingen, USA).
The expanded UCMSCs and the BMMSCs were also characterized by immunophenotypic analysis. About 50,000 cells were stained with one or two of the following monoclonal antibodies directly conjugated with FITC or PE to CD73, CD105, CD45, SSEA4, HLA-ABC, HLA-DR, CD31, CD29, CD44, CD14, and vWF (all antibodies were obtained from BD Pharmingen, USA)as described previously. The stained cells were acquired on FACS Calibur and analyzed using Cell Quest software (BD, CA, USA).
Co-culture of CD34+ cells with MSCs
Several experiments were conducted to come up with the most optimum culture conditions that give stable dependable expansion of sorted CD34+ cells. Five sets of experiments were conducted using different expansion protocols. We also used two types of expansion media for CD34+ cell expansion.
The ‘Liquid culture’ (Table 1) is basically suspension culture of 1×106 CD34+ cells in tissue culture dishes in medium containing cytokines to support maintenance and expansion, whereas, the ‘Co-culture’ pertains to culturing of 1×106 CD34+ HSCs along with 0.4×106 MSCs in the tissue culture flasks. Here the MSCs grew as adherent cells, while the CD34+ HSCs were maintained as suspension cultures in the same culture dishes. All the expansion cultures were incubated at 37°C in presence of 5% CO2 for a period of 14 days with an intermediate passaging on day (7).
Table 1.
Types of expansion medium used for expansion
![]()
The experimental protocols were as given Table 2.
Functional assays
The isolated CD34+ HSCs were also characterized for their clonogenic potential by in vitro clonogenic assays. Twenty thousand cells were taken per 35mm petridish and mixed with 1.5 ml of methyl cellulose (Stem cell technologies, USA). This cell suspension was then plated in triplicates. The plates were then incubated in humidified chambers at 37°C and 5% CO2 for 14 days. The type and the number of colonies generated after 14 days were enumerated using the phase contrast microscope. Each colony was defined as clusters consisting of ≥40 cells. Evaluation was done by two individuals independently.
Cytokine release assay
Cytokine release from the UCMSCs was estimated using an assay kit from Raybiotech (Norcross, GA). This was performed to determine the nature of cytokines produced by UCMSCs in order to support proliferation and maintenance of CD34+ HSCs. Raybiotech cytokine assay kit provided a simple array format to detect multiple cytokine expression from serum free conditioned media of UCMSCs. The UCMSCs were cultured up to 90% confluence in regular medium and subsequently grown in serum free media for a period of 48 hours. The conditioned medium was collected, concentrated 20 times and the assay was performed as per manufacturer’s protocol. The membrane provided with the kit was incubated with the conditioned media concentrate overnight at 4°C. This was then incubated with the primary antibody, followed by biotin conjugated antibodies and HRP conjugated streptavidin with intermediate washes between each step. The detection reagent was used for signal amplification and developed used an x-ray film.
Stability studies
Storage: The CD34+ cells expanded with UCMSCs were cryopreserved using in-house freezing medium (containing Human serum albumin, DMSO-Dextran and Saline) for a period of 6 months, 1 year and 2 years (n = 3). The expanded CD34+ cells were frozen at a concentration of 10×106 cells/ml in 1.8ml cryovial. At each time point, the cells were retrieved from liquid nitrogen container and thawed using the routine thawing medium. The cells were checked under the microscope for morphology, cell surface marker expression and viability.
Statistical analysis
The data was analyzed using student’s paired 2 tailed t-test to determine statistical significance. Values are given as mean±SD (standard deviation).
Go to :
Results
Phenotypic characterization
CD34+ HSCs were characterized for the expression of cell surface markers known to be associated with the maturation status of these cells. The cells sorted using magnetic beads gave 98% pure population of CD34+ cells. These cells were positive for CD33 (21%±2.1), CD38 (27%±1.9), CD133 (57%±1.32), CD31 (99%±0.21), CD44 (99%±0.16), VLA-4 (91%±1.23) and CD11a (63%±1.98) markers as shown in Fig. 1 (n=10). Under the microscope, CD34+ HSCs were seen as refractile round cells. Post sorting, the average viability was more than 97%.
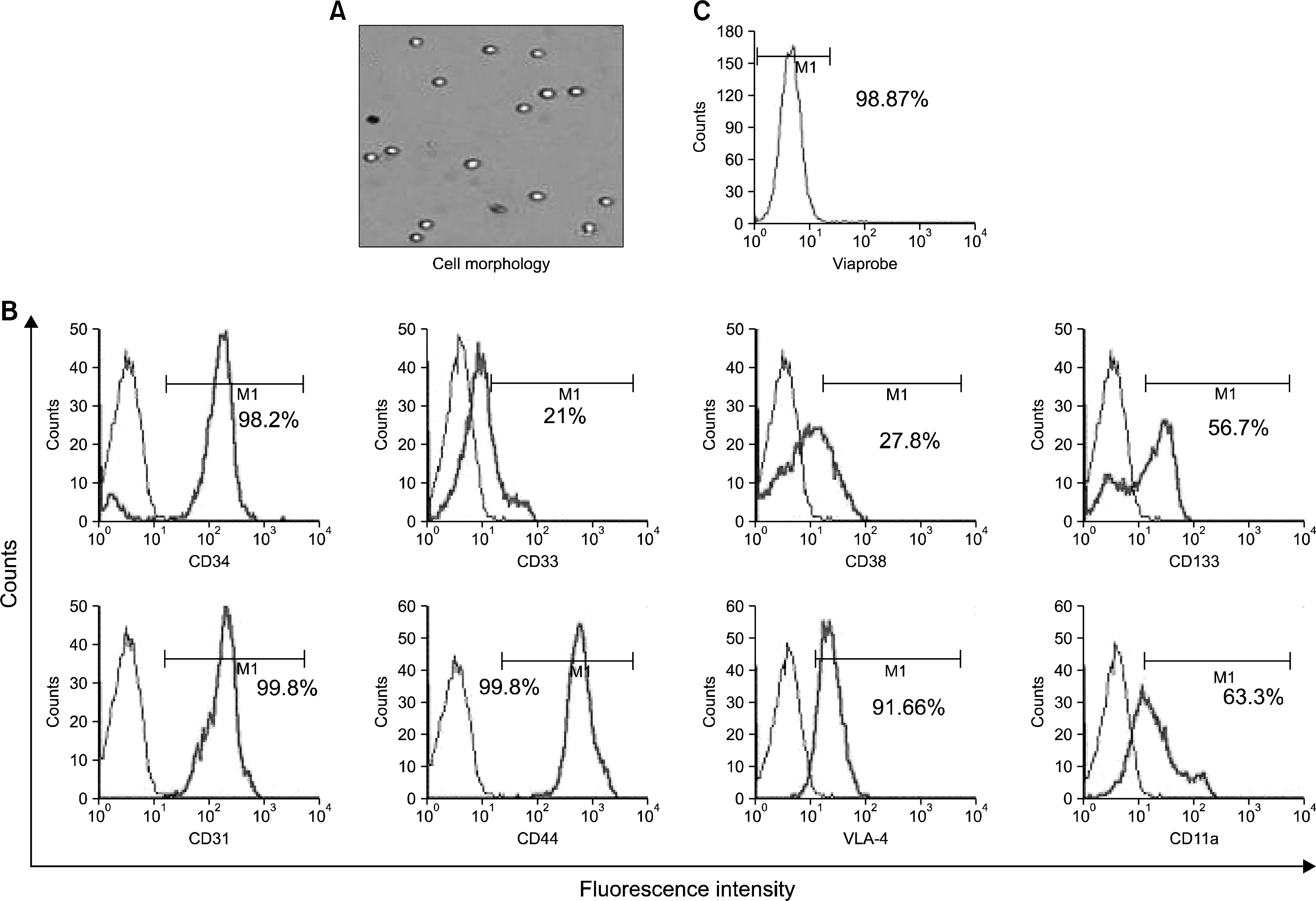 | Fig. 1.Morphology and Characterization of CD34+ HSCs and HPCs isolated from hUCB (n=10). (A) Shows the morphology of the isolated CD34+ HSCs and HPCs after isolation from UCB. These cells are shiny and round in shape in culture. (B) Shows the characterization of the isolated cells. These cells are 98% positive for CD34. They show low expression of CD33, CD38 and CD133, but very high expression of cell adhesion molecules CD31, CD44 and VLA-4. (C) Their viability is dependant on many factors. Mainly the transport conditions and the time taken. For all samples that were received and processed within 48 hours of collection showed very good viability, above 90%.
|
Similarly, the UCMSCs were also characterized using flow cytometer. All the UCMSC lines (n=10) expressed markers similar to those expressed by bone marrow derived MSCs (BMMSCs). These cells were positive for CD73, CD105, CD90, SSEA-4, CD29, CD44, HLA-ABC but were negative for hematopoietic markers CD45, CD31, CD14 and other markers like CD271, vWF and HLA class II (Fig. 2). Average viability of UCMSC samples (n=10) was observed to be 99%.
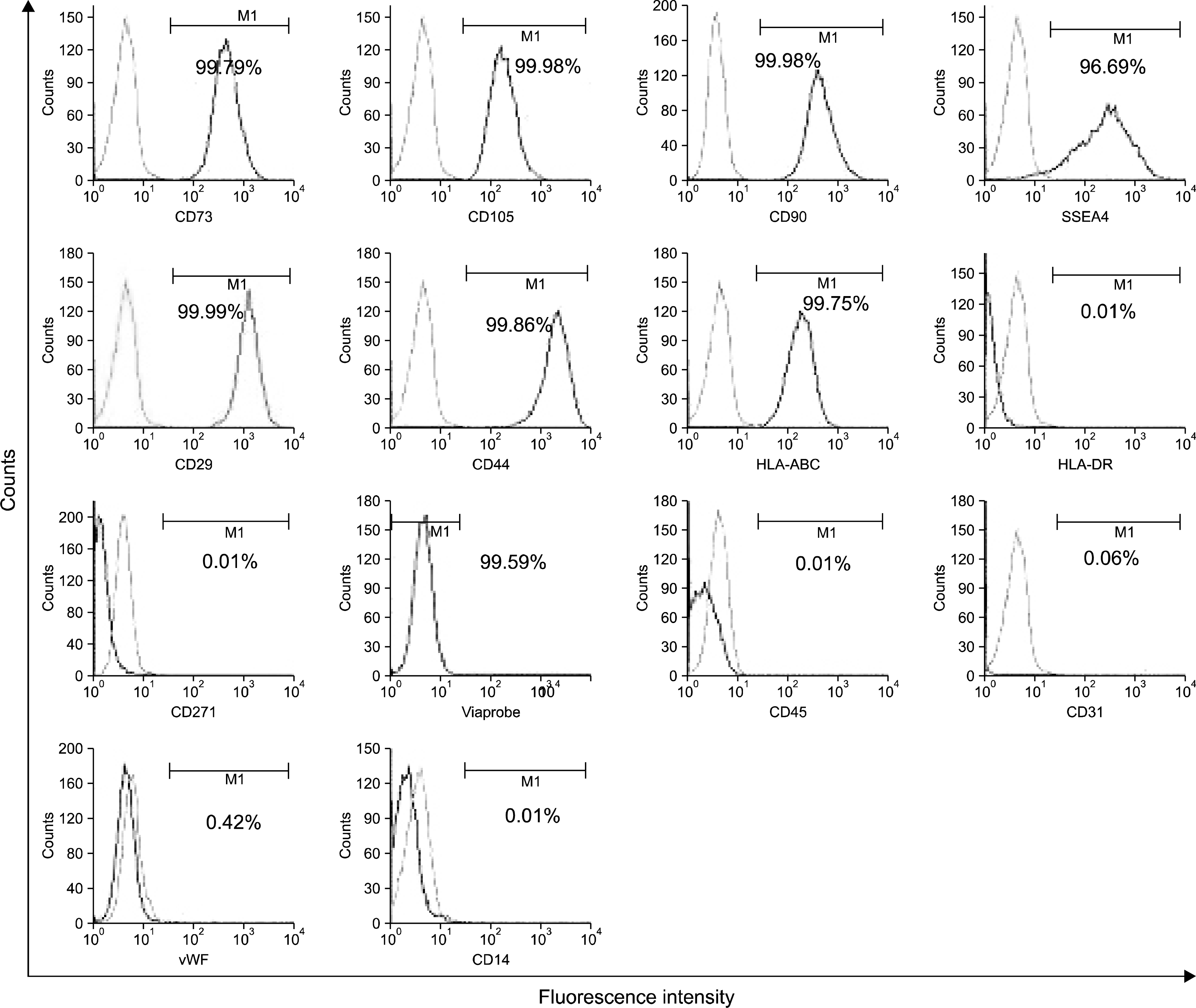 | Fig. 2.Characterization of UCMSCs. The umbilical cord tissue derived MSCs were characterized at every passage after harvest and before storage or use for other experiments. The cells were checked for the expression of mesenchymal stem cell markers, hematopoietic markers, cell adhesion molecules and the viability. It was observed that UCMSCs were 98∼99% positive for all the MSC markers and cell adhesion molecules and were negative for hematopoietic markers. They showed a typical MSC phenotype - CD73+/CD105+/CD45-/SSEA-4+ /HLA-DR-/HLA-ABC+/CD31-/ CD29+/CD44+/CD14-/vWF-/CD90+/CD271-.The cells maintained >95% viability at all passages. This is a representative figure for the 10 UCMSC samples.
|
In vitro expansion of CD34+ HSCs
The cells cultured in expansion medium-1 showed a 10 fold increase in TNCs and about 5 fold increase in CD34+ cells. CD34+ cells cultured in expansion medium-2 were collected on the day of passaging (day 7) and on the day of harvest (day 14). The expanded cells maintained their round shiny morphology during the culture period (Fig. 3A). The cell counts were assessed using a cell counter. The flow cytometric analysis of the expanded cells showed about 10% positivity for CD34 marker (Fig. 4A) after harvest on day 14. At the end of the culture period, we obtained a 6 fold increase in the total number of nucleated cells (Fig. 5A), but CD34+ HSCs and/or HPCs showed less than 1 fold increase (Fig. 5C). The expanded CD34+ cells started losing their stem cell markers and differentiated to lineage committed cells when cultured beyond 14 days. Hence, the culture period was optimized to 14 days and upto passage 2 only for all the expansion experiments.
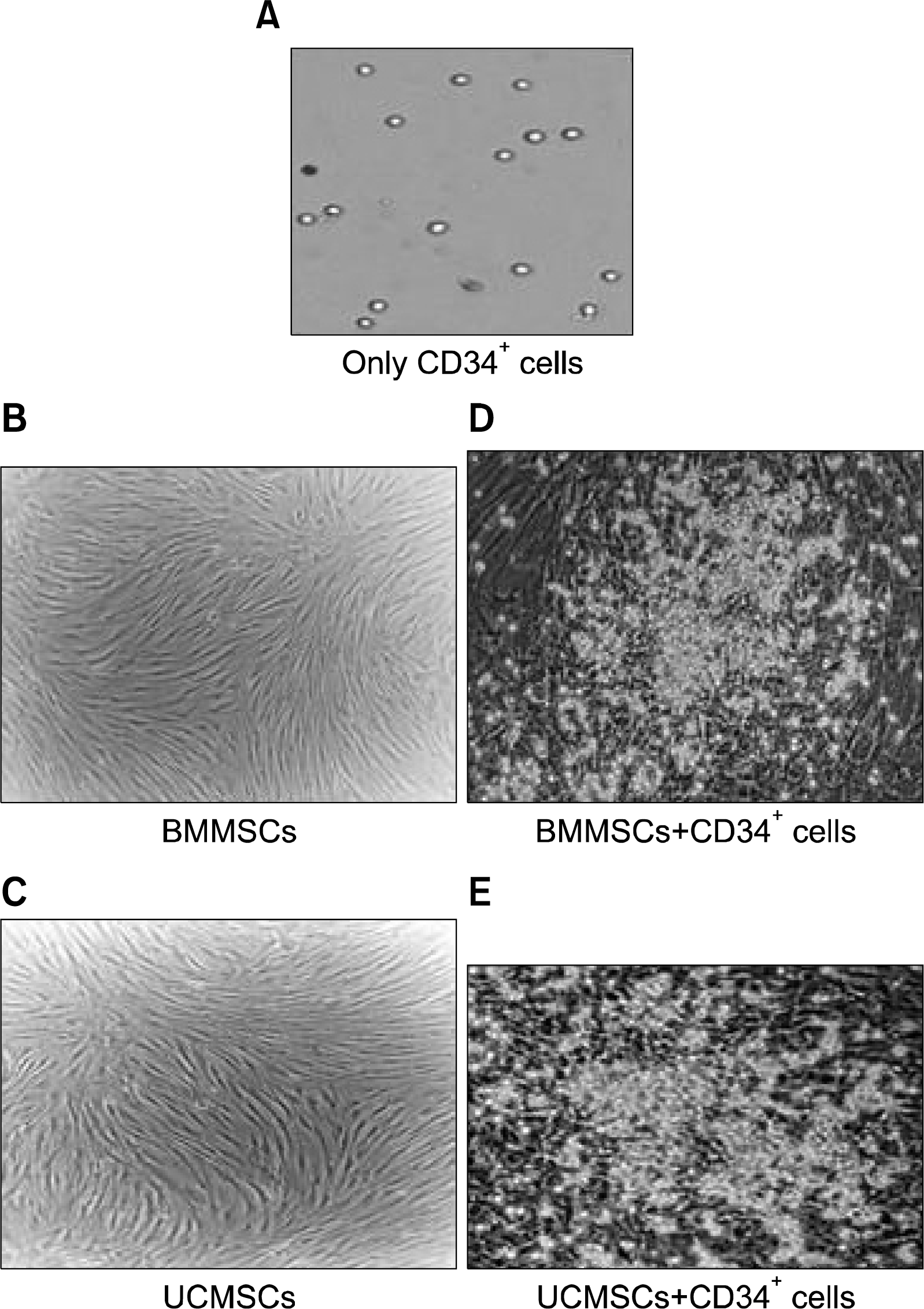 | Fig. 3.Morphology of cells. (A) Only CD34+ cells in culture appear as round shiny refractile cells. They grow in suspension culture. (B) Shows spindle shaped morphology of confluent BMMSCs. (C) Shows the round CD34+ cells attached to the basal BMMSCs during the culture period. (D) Shows spindle shaped morphology of confluent UCMSCs. (E) Shows the round CD34+ cells attached to the basal UCMSCs during the culture period. All the cells in the picture are as observed in 10X magnification.
|
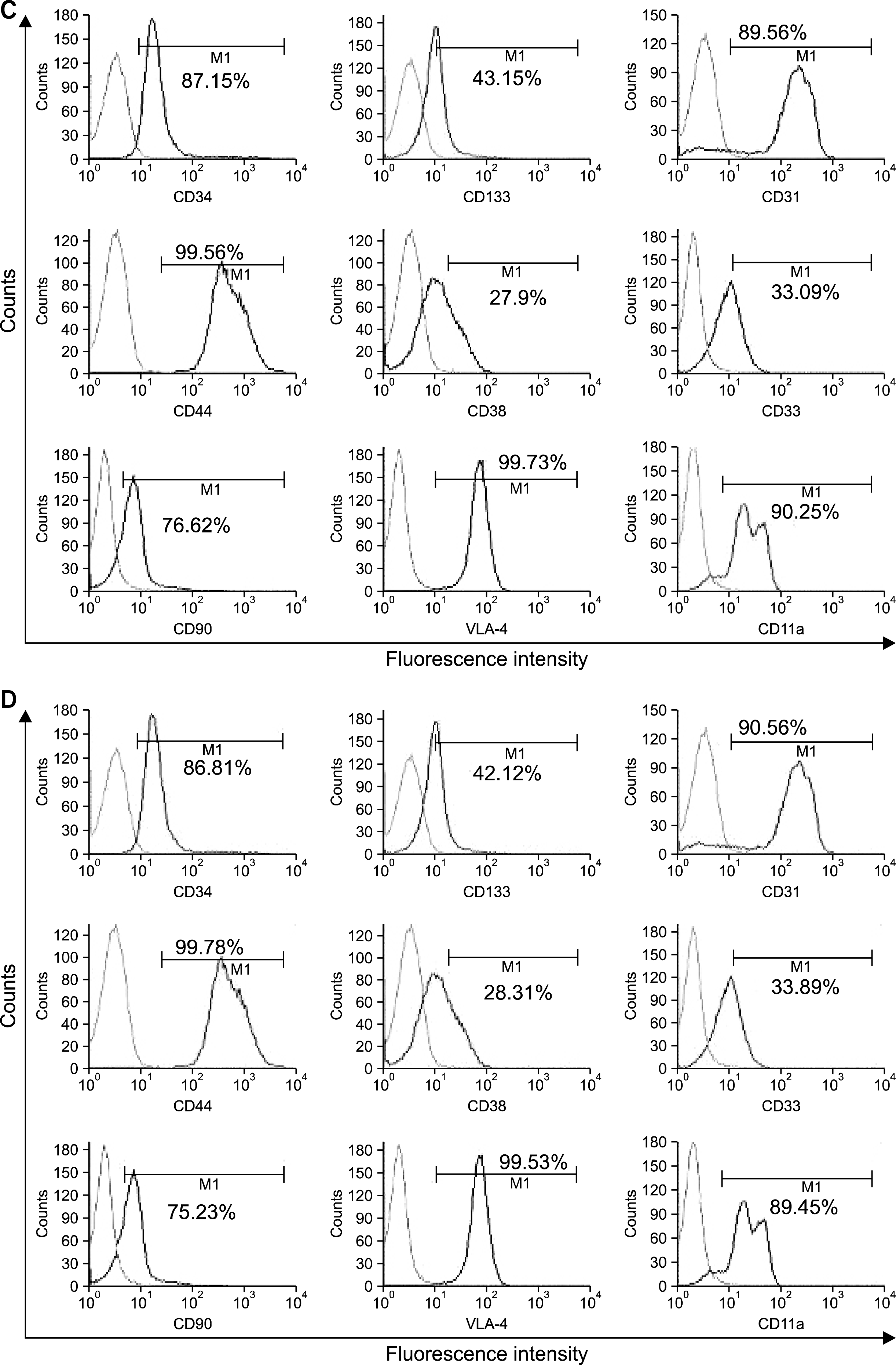 | Fig. 4.Expansion of CD34+ cells. The CD34+ cells were cultured in static liquid cultures or as co-cultures with either BMMSCs, or autologous or allogeneic UCMSCs. (A) Static liquid culture caused expansion of TNCs without expansion of HSCs. The cells that were expanded were negative for CD34, CD90, VLA-4 and CD11a which are the characteristic markers for HSCs. They were, however, positive for CD33, CD38, CD133 and the adhesion molecules- CD31and CD44. (B) The CD34+ cells expanded with BMMSCs were 63% positive for CD34, 74% positive for CD90, 99% positive for VLA-4 and 89% positive for CD11a They were, also positive for CD33, CD38, CD133 and the adhesion molecules- CD31and CD44. (C) The cells that were expanded with UCMSCs from the same cord were 87% positive for CD34, 76% positive for CD90, 99% positive for VLA-4 and 90% positive for CD11a (LFA-1) and also positive for CD33, CD38, CD133 and the adhesion molecules- CD31and CD44. (D) The CD34+ cells expanded with allogeneic UCMSCs showed a similar immunophenotype as that of the ones expanded with autologous UCMSCs. Thus, the UCMSCs, both, autologous as well as allogeneic, supported ex vivo expansion of CD34+ HSCs. All the plots are representative figures for the 5 independent experiments carried out.
|
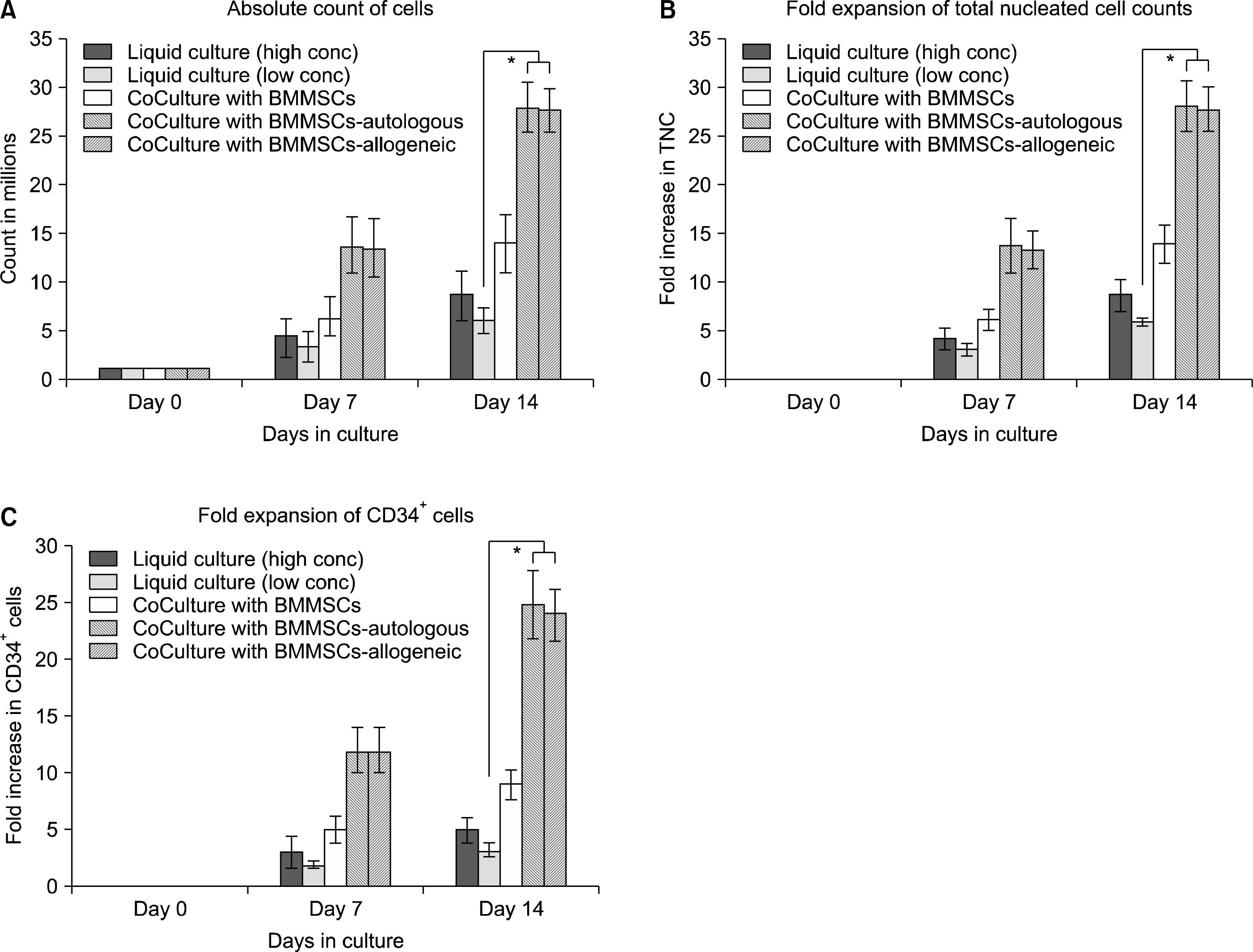 | Fig. 5.Comparative studies. (A) Comparison of absolute count of TNCs as obtained at different time-points under different culture conditions. When the CD34+ cells were cultured alone for a period of 14 days with a starting count of 1 × 106 cells, they increase to 10×106 (±1×106) cells in case of cytokine mediated expansion at high concentration of growth factors and 6×106 (±1×106) cells at low concentration of growth factors. About 14×106 (±1.6×106) cells were obtained when cultured with BMMSCs, while 28×106 (±2.1×106) cells were obtained when the CD34+ cells were cultured with either the autologous or the allogeneic UCMSCs as the supporting cells (*p<0.001). (B) Comparison of fold expansion of TNCs. The CD34+ cells cultured alone showed 10 (±1.2) fold increase in case of cytokine mediated expansion at high concentration and 6 (±0.8) fold increase at low concentration of growth factors. Fourteen (±1.4) fold increase was seen when cultured with BMMSCs, while 28 (±2.1) fold increase was observed when the CD34+ cells were cultured with UCMSCs, either autologous or allogeneic (*p<0.001). (C) Comparison of fold expansion of CD34+ HSCs. When the CD34+ cells were cultured alone for a period of 14 days, 5 (±0.92) fold increase was observed in case of cytokine mediated expansion a high concentration of growth factors and 3 (±0.84) fold increase at low concentration of growth factors. But we could obtain a 9 (±1.6) fold increase with BMMSCs, while 24 (±2.14) fold increase was observed when the CD34+ cells were cultured with UCMSCs, either allogeneic or autologous (*p<0.001).
|
The CD34+ cells, when cultured with allogeneic BMMSCs as feeder cells, maintained their round shiny morphology during the culture period and appeared very healthy (Fig. 3D). About 63% of expanded cells were positive for CD34 expression (Fig. 4B). By the end of 14 day culture period, we could obtain a 14 fold increase in the total nucleated cell count (Fig. 5B) and a 9 fold increase of CD34+ HSCs/HPCs (Fig. 5C). When these cells were plated in in vitro methylcellulose clonogenic assays, they could form as high as 300 colonies per 10 million cells as compared to 100 colonies per 10 million cells obtained with the unexpanded CD34+ cells (Fig. 6B). We could obtain a variety of colonies like CFU-GEMM representing the very primitive population of stem cells, CFU-GM as well as CFU-E, showing presence of progenitors for all cell types belonging to the hematopoietic lineage (Fig. 6A). This was uniformly observed in all the experiments conducted (n=5).
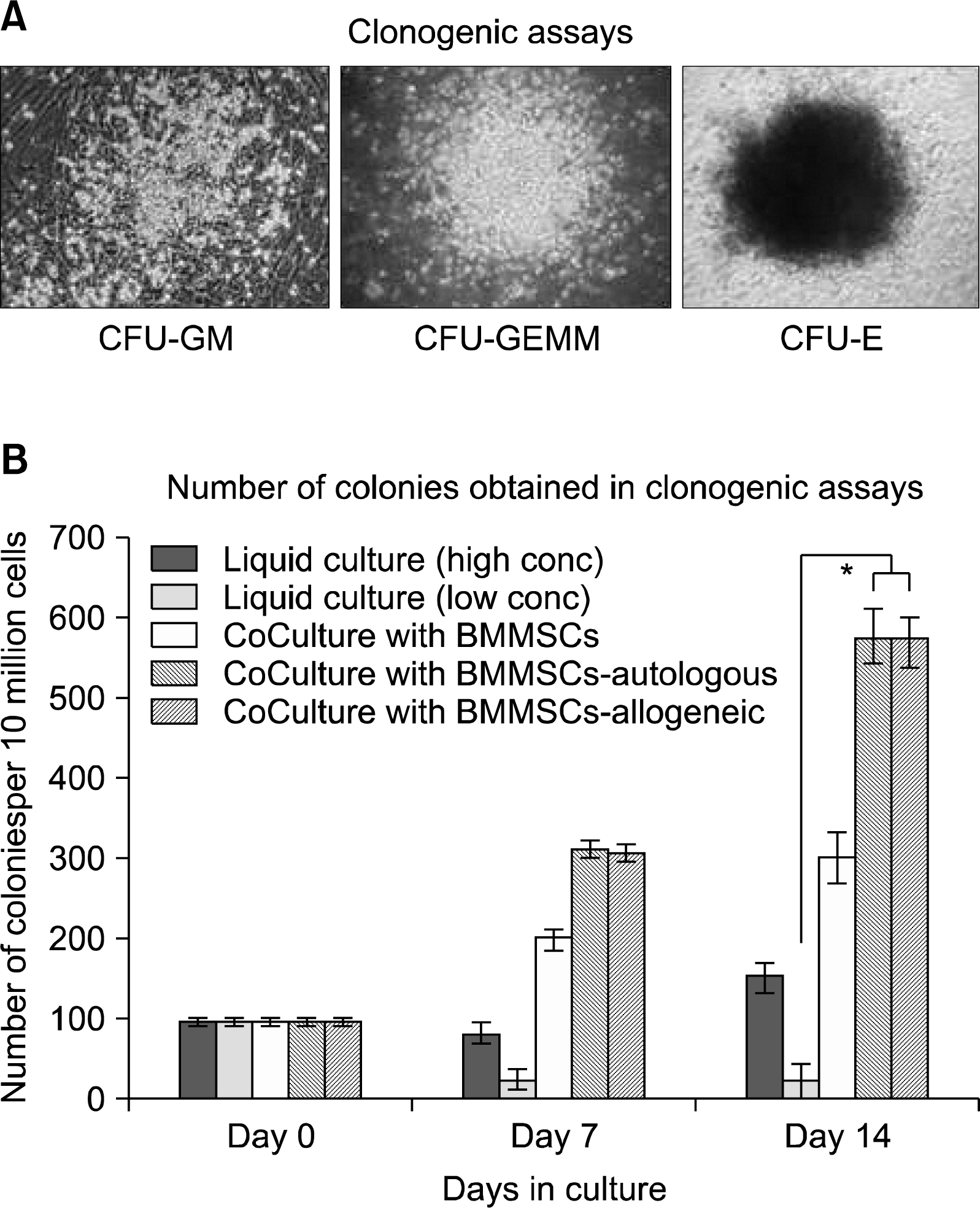 | Fig. 6.Functional characterization of expanded CD34+ cells by in vitro clonogenic assays. (A) Morphology of colonies formed by expanded CD34+ cells. The expanded CD34+ cells were plated in semisolid methylcellulose agar plates and incubated for a period of 14 days at 37°C and 5% CO2 in a humidifies chamber. At the end of the incubation period, the colonies were counted to determine their functionality, i.e. their ability to form different colonies. We could obtain all types of colonies, like colonies formed by early progenitor cells -a. CFU-GEMM, b. CFU-GM and c. CFU-E. (B) Comparative analysis. Since the assay was done in triplicates, the results were calculated as an average of the plates from the same starting number of cells. Compared to about 100 colonies with unexpanded CD34+ cells, we could obtain on an average 45 colonies with CD34+ cells expanded at a low concentration of growth factors, 300 colonies with BMMSCs and more than 550 colonies when cultured with UCMSCs, both autologous as well as allogeneic. The data is a compilation of 5 different experiments (*p<0.001).
|
When CD34+ cells were expanded with autologous or allogeneic UCMSCs as feeders, the expanded cells appeared to be healthy. They maintained their refractile morphology and viability (Fig. 3E). The fold expansion of CD34+ cells was compared with that of cytokine mediated liquid expansion. We could obtain a ∼28 fold increase in the total nucleated cell count (TNC) with autologous UCMSCs and ∼27 fold increase with allogeneic UCMSCs (Fig. 5B). The fold expansion obtained with UCMSCs was significantly higher as compared to that obtained in cytokine mediated liquid cultures (p-value<0.001) (Fig. 5C). Twenty four fold increase of CD34+ HSCs / HPC was observed with autologous as well as allogeneic UCMSCs as shown in Table 3. When these cells were plated in in vitro methylcellulose clonogenic assays, they could form about 550∼600 colonies per 10 million cells in both the cases (Fig. 6). These values were significantly higher (p-value <0.001) as compared to the number of colonies obtained with same number of cells expanded in cytokine mediated culture system (Fig. 6B).
Table 3.
Comparative study of CD34+ cell expansion using different strategies in expansion medium -2. The comparative analysis shows that UCMSCs were able to give more number of functional expanded cells as studied by flow cytometry and in vitro clonogenic assays
![]()
Expression of cell surface markers by expanded CD34+ cells
The CD34+ cells cultured by liquid culture system in expansion medium-2, were analyzed for expression of other cell surface markers. It was observed that more than 90% cells were positive for the lineage determining markers CD33 and CD38. The expanded cells were negative for the cell adhesion molecules VLA-4 (Very late antigen-4) and LFA-1 (CD11a, lymphocyte function associated antigen -1) which are known to help maintain these cells in stem cell stage. The cells were however, also positive for CD31 and CD44 (Fig. 4A).
More than 60% CD34+ cells expanded with allogeneic BMMSCs as the feeders in co-culture system, were positive for CD34, while 70∼80% cells were positive for CD33 and CD38. These cells were 99% positive for VLA-4 and more than 85% positive for LFA-1 (CD11a). They were also very strongly positive for CD90, CD31 (PECAM) and CD44 (Fig. 4B), indicating their stemness status and good potential for engraftment.
With autologous UCMSCs as supporting cells in co-culture system, we observed more than 85% of the cells positive for CD34 and more than 75% cells maintained CD90, an important stem cell marker. These cells were strongly positive for cell adhesion molecules, VLA-4, LFA-1 (CD11a), CD31, and CD44 which are responsible for engraftment. Only about 30% cells were positive for CD33 and CD38, however these cells co-expressed CD34 antigen on their surface (Fig. 4C). This shows that the cells expanded with autologous UCMSCs, maintained their stemness and did not get differentiated into lineage specific cells during the 14 day culture period.
Interestingly, our data also shows that allogeneic UCMSCs were equally capable of supporting the growth and expansion of CD34+ HSCs. At the end of expansion period, more than 85% of the cells were CD34+ and 70% (±3%) cells maintained CD90 expression. These cells were also strongly positive for cell adhesion molecules, VLA-4, LFA-1 (CD11a), CD31, and CD44 confirming their suitability for engraftment. Similar to the data presented above, even with allogeneic UCMSCs as feeders, about 30% of HSCs were positive for CD33 and CD38, and majority of these cells co-expressed the CD34+ marker (Fig. 4D) confirming their stemness.
UCMSCs assist in increasing the colony forming units per sample of UCB
The clonogenic potential of the stem cells is of paramount importance for successful transplantation and long term engraftment. More than 250 clonogenic units (CFUs) per 10 million cells, which constitute erythroid, myeloid, granulocytic colonies, is a requirement to obtain good engraftment. Our results show that the CD34+ HSCs expanded in liquid cultures could form only about 150 CFUs (Fig. 6B). However when HSCs were expanded with BMMSCs, we obtained on an average of 300 colony forming units per 10 million cells (Fig. 6B). In this study, we report an average of 550∼580 colony forming units per 10 million cells when these sorted CD34+ HSCs were co-cultured with UCMSCs as. These results were obtained from five independent experiments performed in our laboratory.
Allogeneic expansion of CD34+ cells is comparable to that obtained with autologous UCMSCs
The use of UCMSCs as feeder cells, irrespective of whether it is derived from autologous or allogeneic sources, for expansion and maintenance of CD34+ HSCs did not matter. The expansion using either of them was comparable to the other.
Karyotypic analysis of expanded CD34+ cells
The expanded CD34+ cells were analyzed for karyotypic abnormalities, at passage 2. In all instances, these cells were found to be of normal karyotype (Fig. 7).
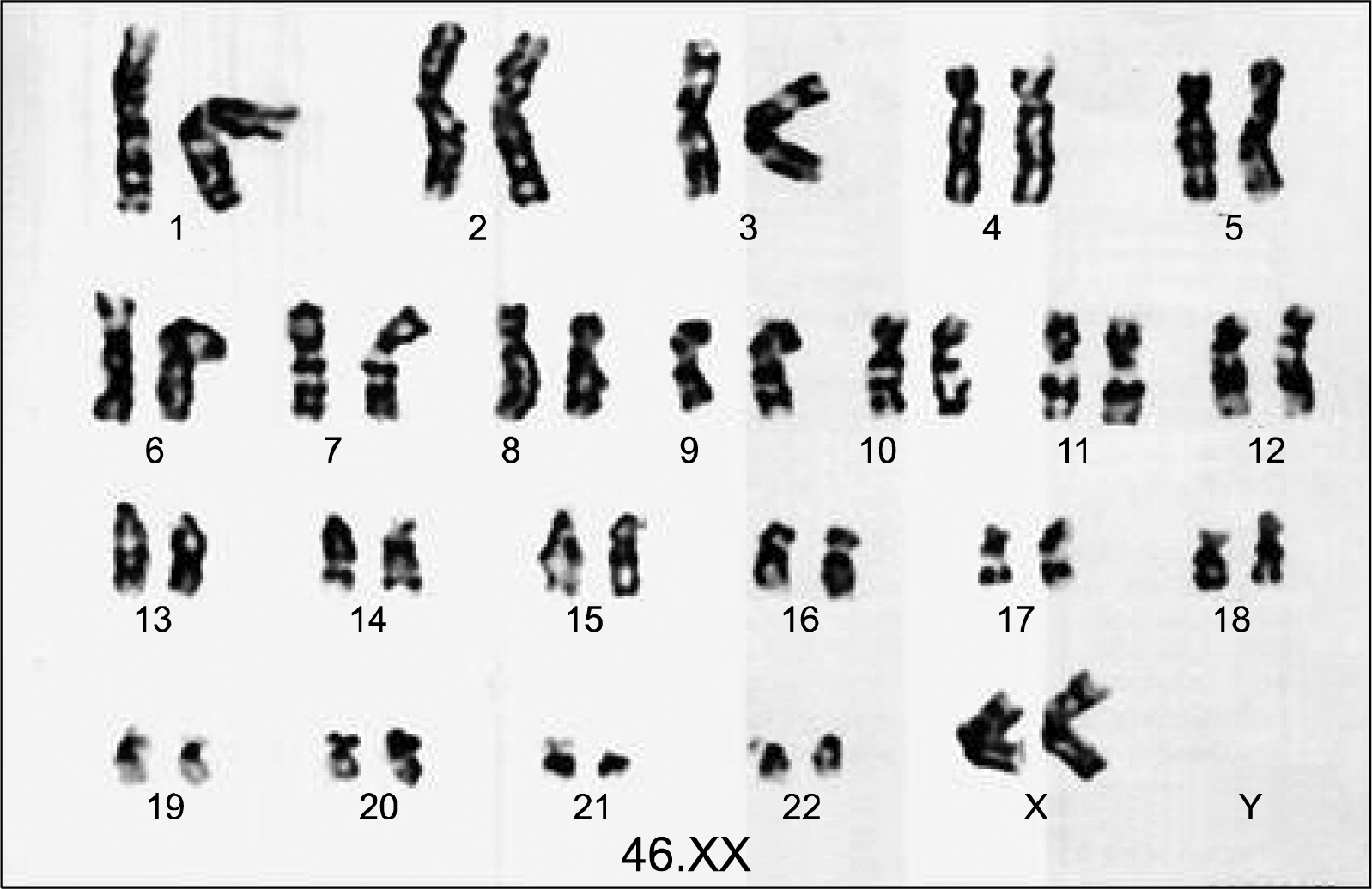 | Fig. 7.Karyotypic analysis of the expanded CD34+ cells at passage no. 2 (P2). The in vitro expanded CD34+ cells were checked for their karyotypic stability. It was observed that their not no abnormality induced by the culture systems. Hence, the culture conditions were safe with respect to maintenance of their karyotype.
|
Stability of expanded CD34+ cells
The post thaw viability of expanded CD34+ cells was determined to be more than 95% (Fig. 8). Ninety percent of these cells were positive for CD34 and they also expressed CD90, CD31, CD44, VLA-4 and LFA-1 (CD11a) antigens. About 30% of the cells were positive for CD33 and CD38, while about 40% cells were positive for CD133. These data collectively showed that the HSC immunophenotype was maintained and the cells were stable and functional even after cryopreservation and thawing.
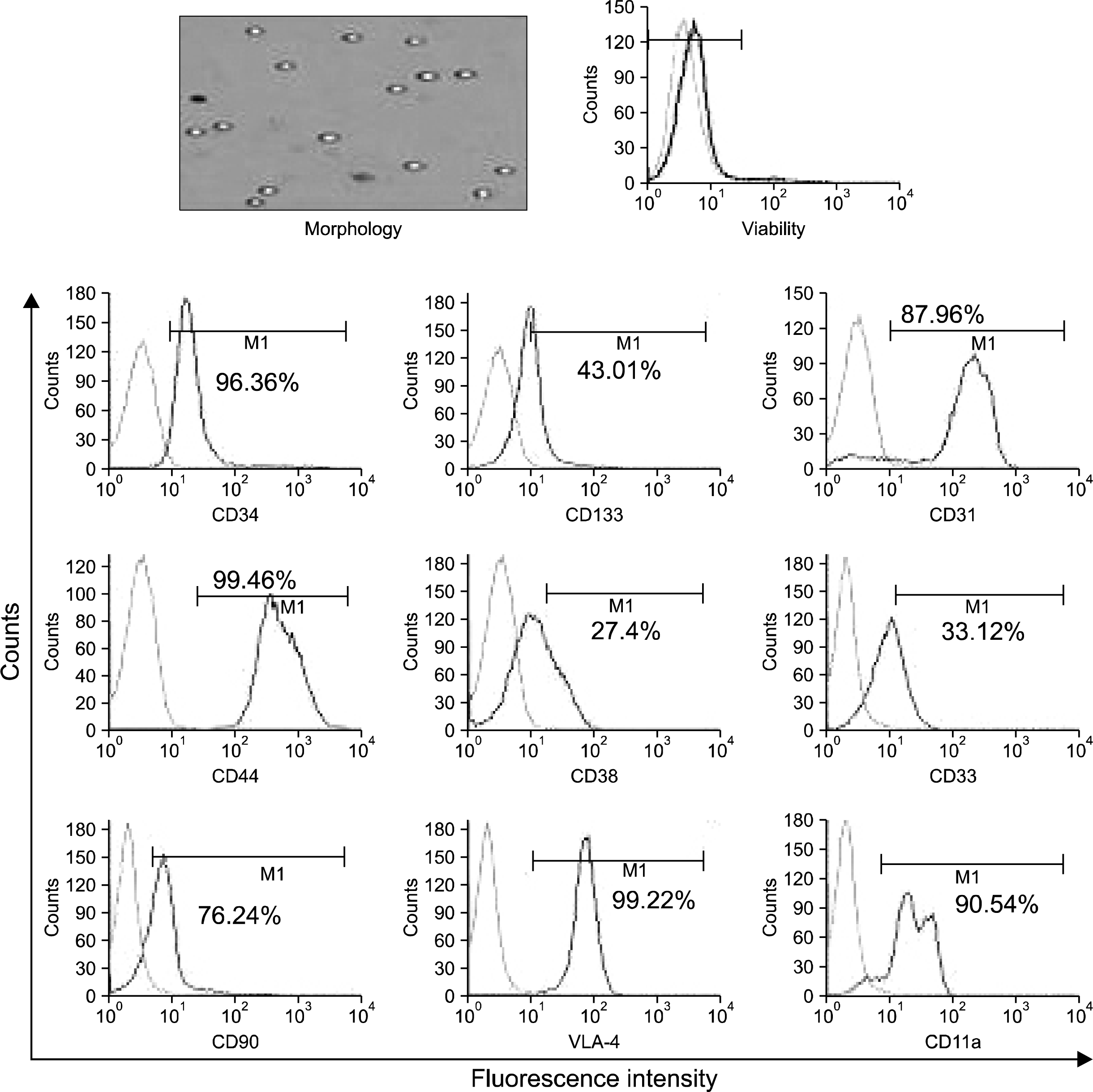 | Fig. 8.Stability studies post freeze - thaw of the expanded CD34+ cells. The CD34+ cells expanded with UCMSCs as the supporting cells were cryopreserved post expansion. These cells were thawed after a period of 6 months and checked for their stability. There was no change in their morphology as well as their phenotype. Post thaw viability was observed to be 97% (n=3).
|
Cytokine release assay
We could demonstrate cytokine release by UCMSCs and BMMSCs. Secreted cytokines, chemokines and other biological molecules secreted by UCMSCs and BMMSCs may also participate in the expansion and maintenance of CD34+ cells. In this study, we have examined the presence of SDF-1, Flt-3, IL-6, ENA-78, GRO, Thrombopoetin, G-CSF, GM-CSF, SCF, LIF and other factors in the supernatant of two MSC cultures (Fig. 9). Some of these factors are known to support growth and expansion of HSCs in culture. We observed that the intensity of certain cytokines released by UCMSCs were clearly higher than those released by BMMSCs, although we did not quantitate the concentration of the cytokines. UCMSCs released ENA-78, G-CSF, MCP-2, EGF, thrombopoietin and LIF but BMMSCs did not secrete these cytokines and growth factors as shown in Table 4. The expression of GM-CSF, SDF-1, Angiogenin, Flt-3 ligand and HGF secreted by BMMSCs was comparatively lower to that produced by UCMSCs. These results, though qualitative in nature, may suggest that UCMSCs obtained from the fetal cord tissue, release important cytokines and chemokines essential to support growth and expansion of other cell types.
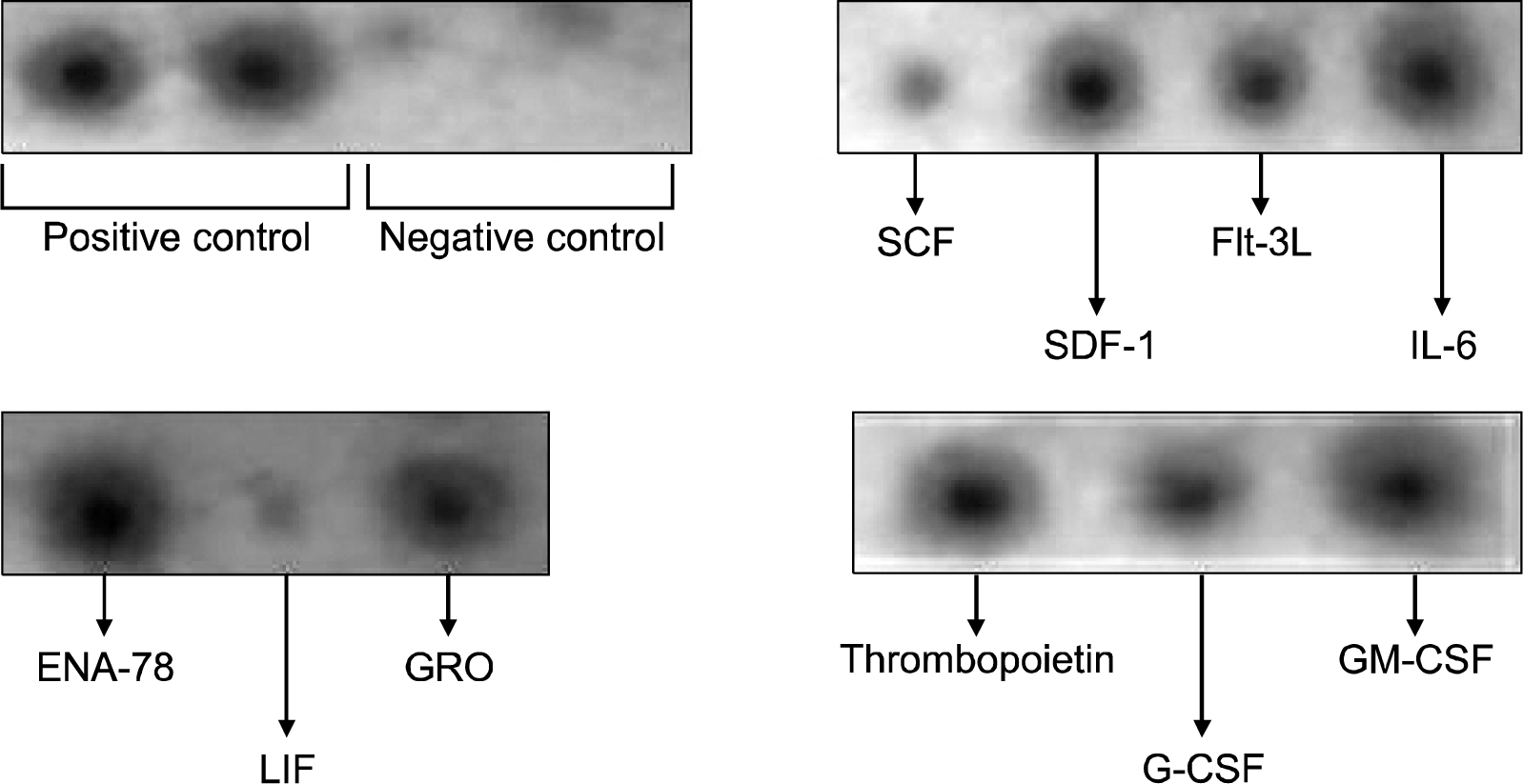 | Fig. 9.Cytokine bead array analysis for determination of cytokines released by UCMSCs. The supernatant of the UCMSCs collected after 48 hours of culture and concentrated 20 times was used. The image from the membrane was then developed using an X-ray. The UCMSCs released high concentrations of SDF-1, Flt-3, IL-6, ENA-78, GRO, Thrombopoetin, G-CSF, GM-CSF and low concentration of SCF and LIF which are responsible for supporting the maintenance and expansion of CD34+ cells in culture.
|
Table 4.
Cytokine bead array analysis of cytokines released by UCMSCs and BMMSCs. It has been observed that UCMSCs release few of the cytokines more as compared to BMMSCs. LIF, HGF, Flt3, G-CSF, GM-CSF, Angiogenin, TPO etc are all released at higher concentrations by UCMSCs as compared to BMMSCs
![]()
Go to :
Discussion
Umbilical cord blood is an excellent source of hematopoietic stem cells (22). The success in cord blood transplantation has been demonstrated in more than 10,000 cases worldwide so far (23). These cases have been predominantly for hematopoietic transplantation in children suffering from various types of genetic disorders or neoplasias and require HSC transplantation. The limited number of HSC content of cord blood units has been the primary reason for their restricted application only in children as compared to adults. Broxmeyer HE and Kurtzberg et al have shown the significance of total nucleated cell dose (TNC) and CD34+ cell counts being the most important parameters that determine the outcome of HSC transplantation (14, 24). In addition, Wagner JE and Gluckman in their compilation of the data on umbilical cord blood transplantations in the last 20 years have found a correlation between HLA match and cell dose, such that the deleterious impact of HLA mismatch could be partially compensated for by increasing the cell dose (25). In addition, Rubeinstein et al and Wagner et al in their observation on unrelated cord blood transplantation have demonstrated the importance of both TNC and CD34 content of the graft being responsible for faster engraftment (3, 26).
Our cord blood bank, at Reliance Life Sciences Pvt Ltd., has units representing various ethnic populations of the Indian subcontinent. Needless to say, this representation will bring in more match rates for patients from all different Indian ethnicities. Pitchappan et al and Querol et al. have independently described their experiences on incidence of matches keeping in mind the ethnic mix, highlighting its significance (27, 28). On the same lines, our match rate has been encouraging and our observations in the last 4 years on this subject are under review. There were several consensus meetings however, whether or not CD34+ cell counts alone will affect transplant outcome. However, Ian McNiece in 2004, in his communication regarding CD34 content strongly recommended and suggested the importance of this factor (16).
The delay in engraftment kinetics is likely to be primarily due to relatively low number of CD34+ stem and progenitor cells in cord blood units, because the total cell number (TNC) and total CD34+ cell number in a graft correlate with engraftment kinetics (3, 26). Recently, Scaradavou et al published results on transplantation using multiple cord blood units, compelling researchers and clinicians to consider this modality even for adults (29). In addition, this strategy was also tested in non-myeloablated patients with some successes as shown by Barker et al. (8, 30). However, there is definite alloimmunization and inter-donor sensitization hazards in using multiple cord blood units. This may be due to incompatibilities on account of mismatch in one or more loci. Thus adequate dose of CD34+ cells from a single UCB sample will be the most desirable option, which Ian McNiece’s group demonstrated through his work on ex vivo expansion of hematopoietic stem cells from cord blood (16, 31). But this method of expansion relied on addition of exogenous growth factors to prevent apoptosis and stimulate proliferation, potentially risking differentiation at the expense of self renewal, thereby decreasing primitive hematopoietic progenitor cells from the microenvironment (17).
The idea of using bone marrow MSCs as the feeder layer for CD34+ cell expansion has became a useful approach for successful engraftment (16). The protocols and processes seem easy enough to adopt, but there are few limitations. Autologous BMMSCs as feeder layer to expand CD34 was shown in clinical trials by SN Robinson et al. (17). The results clearly show better engraftment and lower GvHD rates. Bone marrow collection, however, is an invasive process and suitable donors may not be always available. Radiation and chemotherapy induced myelodepression further makes autologous bone marrow derived MSCs a difficult proposal. Furthermore, patients requiring allogeneic stem cell transplantation following radiation and chemotherapy treatment may not be able to donate bone marrow for MSC isolation and expansion; a pre-requisite in order to use these cells to expand CB derived CD34+ cells.
Our study was focused on examining alternate sources of MSCs like the umbilical cord matrix for their ease of availability and its growth kinetics (20, 32). Our group has shown derivation of UCMSCs, their plasticity and their differentiation potentials in vitro and in vivo (33). The umbilical cord blood derived hematopoietic stem cells reside in close association with umbilical cord throughout the gestation period (Table 5). The physical association between MSCs of umbilical cord and the HSC of UCB during the gestation period may enable UCMSCs to provide optimal support for growth of HSCs. Hence, in our study, we have aimed at understanding if MSCs derived from the umbilical cord can support the expansion of UCB derived CD34+ HSCs, in vitro too.
Table 5.
Comparative characterization of UCMSCs and BMMSCs. The UCMSCs and the BMMSCs were immuno-phenotyped using flow cytometry. It was observed that the umbilical cord derived MSCs were phenotypically similar to those derived from bone marrow
| BMMSCs | UCMSCs | |
|---|---|---|
| CD 73 | ++ | ++ |
| CD 105 | ++ | ++ |
| CD 45 | − | − |
| CD 90 | ++ | ++ |
| SSEA 4 | + | + |
| HLA-DR | +/− | − |
| HLA-ABC | + | + |
| CD 31 | − | − |
| CD 44 | ++ | ++ |
| CD 29 | ++ | ++ |
| CD 14 | − | − |
| vWF | − | − |
| CD 271 | + | − |
![]()
We have shown the cytokine release profile of UCMSCs in vitro. These cells released SDF-1, Flt3, Thrombopoetin, G-CSF, GM-CSF, IL-6, SCF and LIF (Table 4) which are known to play important role to support HSC growth. In addition, UCMSCs also expressed the cell adhesion molecules CD29 and CD44 that are required for homing of HSCs to the tissue site in vivo. The UCMSCs can thus perhaps create a hematopoietic stem cell niche, serve as structural elements and nurse HSCs. The UCMSCs, therefore, although similar to BMMSCs in many ways, seem to be much better candidates to support HSC engraftment after stem cell transplantation, for all the reasons described above.
UCMSCs are known to possess important immunomodulatory and immunosuppressive characteristics (under publication in J Immunology and Cell Biology) (34, 35). We have recently shown that these cells do not induce allogeneic T cell proliferation in an in vitro MLR assay. This inability to induce T cell activation is maintained by UCMSCs even in the presence of a potent inflammatory activator, interferon gamma (IFN-γ). They also do not express HLA class II molecule on its surface but on the contrary express the negative regulator of immune system, B7-H1. Further, the UCMSCs are also able to inhibit an ongoing allogeneic mixed lymphocyte reaction between T cells and dendritic cells. This confirms that the UCMSCs are immunosuppressive in nature. They also release molecules like indoleamine 2, 3 dioxygenase (IDO), HGF, IL-12, which can help create an immunosuppressive environment around themselves. Hence, the UCMSCs can be termed as immunopriviledged cells.
We have shown through this study, the capability to expand CD34+ cells using unrelated umbilical cord matrix derived mesenchymal stem cells resulting in 25∼28 fold expansion (Table 3). This is in agreement with Ian McNiece’s observation, although they used bone marrow derived MSCs (16). They obtained about 16∼25 fold expansion with good stability in culture. We could obtain a slightly higher i.e. 25∼28 fold expansion. However in all these cases, the starting counts varied from as low as 0.8×106 cells to 5×106 cells. So even as low as 1 million CD34+ cell counts gave a dependable expansion up to 28 fold at the end of 2 weeks. All characterization and clonogenic potentials were confirmed and are as desirable. Our experiments to study stability of expanded CD34+ cells stored in liquid nitrogen for varying periods further strengthen its candidature for clinical use (Fig. 6). As shown (Fig. 4) there were no karyotypic abnormalities observed post expansion in all cases. Our studies on more than 25 samples gives us the confidence to limit the expansion to 25∼28 fold at 2 weeks time point, as further culture resulted in differentiation and loss of stemness (data not shown).
The immunological properties established by UCMSCs make allogeneic MSCs as good as autologous MSCs from bone marrow, atleast for expansion purposes. Lack of T cell stimulation and poor dendritic cell activation, despite presence of immune regulator IFN-γ, which is usually present in the transplant setting, makes UCMSC a promising cell for clinical application. Thus readily available CD34+ cell in the appropriate dosage addresses a long time unmet medical need.
Moving forward, if there is a good match available from the umbilical cord blood banks, the clinician can still have the option to get them expanded in vitro using these immunopriviledged cells. It remains to be seen however, if co-transplantation of these two cell types will give acceptable results. Till research findings in this regards get published, in vitro expansion of CD34+ cells looks very promising.
Go to :
 XML Download
XML Download