

 PDF
PDF Citation
Citation Print
Print


Introduction
Cytokine-mediated mobilization of hematopoietic stem cells (HSCs) has become an established method in the field of autologous and allogenic stem cell transplantation. Furthermore, it presents a new concept in tissue repair and regenerative medicine. Currently, G-CSF remains the gold standard mobilizing agent for peripheral blood stem cell transplantation (PBSCT). However, there are many areas for improvement that can have significant clinical impact: specifially, a significant proprtion (10∼20%) of healthy allogeneic donors mobilize poorly in response to G-CSF and this proportion dramatically increases (up to 40%) in autologous donors; inability to predict optimal mobilization times; inability to expanse bone marrow stem cell pool and graft composition (1–3. Thus, intense efforts have been made to alter mobilization strategies, such as Pegfilgrastim, the bicyclam AMD3100, chemokines and their receptors. In the present study, we studied the efficiency of rhEPO plus rhG-CSF on mobilization of peripheral blood stem cells (PBSCs), and also focused on the content of CD34+, CD90+, CD90+ CD34−and Sca-1+ stem cells in the mobilized peripheral blood graft.
Go to : 
Materials and Methods
Animals
The expriment was performed on male mice (KM strain, n=60) weighing about 22±2 g. Animals were provided by Laboratory Animal Center of Yunyang Medical College, China. Animals were housed in polypropylene cages in 12 h dark/12 h light cycles with temperature of the laboratory animal house ranging from 20°C to 25°C, and humidity between 50% to 65%. Saw dust was used as the bedding, which was changed during cleaning of cages on alternate days. All the animals were produced from the Laboratory Animal Center of Yunyang Medical College, China. And all the animals were received care in accordance with Guide for the Care and Use of Laboratory Animals published by the US National Institutes of Health (NIH publication No. 85-23).
Adiministration of rhG-CSF or rhEPO
Mice were divided into 4 groups, each comprising 15 mice:1) adiministration of saline daily for 5 days. 2) adiministration of rhG-CSF (Lot:20090206, produced by Shandong Qilu Phaimaceutical Co., Ltd.) daily for 5 days (250 μg per kg body weight per day, intraperitoneally). 3) adiministraion of rhEPO (Lot: 20081109S, produced by Sansheng Shenyang Pharmaceutical Co., Ltd.) daily for 5 days (1,000 IU per kg body weight per day, intraperitoneally). 4) adiministraion of rhG-CSF in combination with rhEPO for 5 consecutice days (dosage as above). All mice were anaesthetized with 10% Chloral Hydrate and sacrificed at 6 hours, day 1, day 3, day 5 and day 7 after the last injection drugs.
Hematologic parameters
Mobilized-peripheral blood samples were analyzed using Automatic Hematology Analyzer (Cell Dyn 3700, Abbott, United States).
Reverse transcriptase-polymerase chain reaction (RT-PCR) analysis
Total RNA was extracted from WBC or bone marrow cells using the TrizolTM kit (GIBCO BRL, Invitrogen, Paisley, UK) according to the manufacturer's instructions. For PCR, the primers were designed to exclude detection of genomic DNA. As positive controls, total RNA was extracted from mice bone marrow. Primers were used as indicated in Table 3. The PCR was performed using a Hot Star Taq Master Mix Kit (Qiagen, Hilden, Germany) according to the manufacturer's instructions and was carried out in a programmable Applied Biosystems Veriti 96 Well Thermal Cycler (Applied Biosystems, America) with the following conditions: 94°C for 5 min and then 30 cycles, each comprising 94°C for 30s, annealing for 1 min with: GAPDH 62°C, CD34 62°C, CD90 62°C, Sca-1 55°C and then 72°C for 1 min. After the PCR was completed, the tubes were kept for 5 min at 72°C. Negative controls were performed without template. The difference of the expression level of a specific RNA was determined by normalization to the expression level of GAPDH and quantification was made using a standard curve assay (Table 1).
Table 3.
MNC counts at different time after mobilization (109/L) (Mean±S.E.)
| Group | Sixth hour | Firth day | Third day | Fifth day | Seventh day |
|---|---|---|---|---|---|
| control | 0.67±0.46 | 1.04±0.50 | 1.13±0.47 | 1.07±0.19 | 0.99±0.32† |
| G-CSF | 1.24±0.71 | 2.82±1.23 | 2.28±0.76 | 4.18±1.49* | 6.88±2.89 |
| EPO | 2.59±1.48* | 4.84±1.82* | 3.28±1.15* | 3.56±1.62* | 5.53±1.33* |
| EPO+G-CSF | 1.93±0.72 | 2.59±1.25 | 3.63±1.29 | 2.88±1.38 | 5.22±1.94* |
![]()
Table 1.
Primer pairs of GAPDH, CD34, CD90 and Sca-1
![]()
Flow cytometry analysis
At 6 hour, day 1, day 3, day 5 and day 7 after the last adiministration, 1mL peripheral blood was harvested from each mouse by aspiration from heart. Bone marrow cells were obtained by flushing tibias and femurs with phosphate-buffered saline. To determine CD90+/CD34+ and CD90+/CD34− subpopulations, cells were incubated for 40 minutes in the dark at 4°C with the following fluorescein isothiocyanate-(FITC) and phycoerythrin-(PE) conjugated monoclonal antibodies: CD90-FITC and CD34-PE (all from eBioscience, San Diego, USA). Isotype-identical antibodies: IgG2α-FITC and IgG2α-PE (eBioscience) served as controls. Cells were analyzed by two-color flow cytometry using a Coulter Epics XL- MCLTM flow cytometer (Beckman Coulter). Each analysis included 10000 events.
Statistical analysis
Data are shown as mean±standard error of mean. Multiple group comparison was performed by one-way analysis of variance followed by the Bonferroni procedure for comparison of means, and performed by K Independent Samples Test for comparison of rate by way of SPSS13.0 statistical package program for Windows. Comparisons between two groups were performed using the unpaired t-test and Chi-Square test. Values of P less than 0.05 were considered statistically significance.
Go to :
Results
Hematologic parameters
At first, hematologic parameters were analyzed to compare the effects of rhEPO, rhG-CSF+rhEPO with standard rhG-CSF treatment as a positive control according to established mobilizing strategies for PBSCT. The number of leukocytes and mononuclear cells was summarized in Table 2 and Table 3. These data showed leukocytes significantly increased values at the 1st day of rhEPO stimulation (1.67- fold; p<0.001) and at the 3rd day of rhG-CSF+rhEPO stimulation (1.52-fold; p<0.01) compared to the positive control group. Increased levels were also detected for mononuclear cells at different time of rhEPO and rhG-CSF+rhEPO stimulation, but there was no significant compared to the positive control group.
Table 2.
WBC counts at different time after mobilization (109/L) (Mean±S.E.)
| Group | Sixth hour | Firth day | Third day | Fifth day | Seventh day |
|---|---|---|---|---|---|
| control | 1.98±0.91 | 2.00±0.87 | 1.82±0.51 | 1.76±0.26 | 1.88±0.56† |
| G-CSF | 2.50±1.09 | 3.99±1.32 | 3.50±0.99* | 6.67±3.82* | 9.17±3.22 |
| EPO | 3.33±1.44 | 6.67±1.75*‡ | 5.00±1.44* | 5.17±1.98* | 7.50±1.76* |
| G-CSF+EPO | 2.83±1.11 | 3.50±1.44 | 5.33±1.25*‡ | 4.17±1.44 | 7.17±1.52* |
![]()
FACS analyses of CD90+ cells populations in peripheral blood
In order to know whether defined populations of bone marrow stem cells circulate in peripheral blood with the stimulation of rhG-CSF, rhEPO or rhG-CSF+rhEPO, fluorescein-activated cell sortiong (FACS) analyses were performed. We found a significant increase of the defined subtypes of CD90+ cells (CD90+/CD34+ and CD90+/CD34−). Especially, CD90+/CD34− cells were stronger increased than CD90+/CD34+ cells(the percentage of CD90+/CD34+ cells was very low and the data was not shown). After rhEPO application, a higher increase of CD90+/CD34− cells at different time excluded at hour 6 than adiminstration of rhG-CSF and rhG-CSF+rhEPO. At day 5 and day 7, CD90+/CD34− cells were mobilized with the stimulation of rhG-CSF+rhEPO more than rhG-CSF (p=0.000, p=0.025, However, this phenomenon wasn't seen at day 1 and day 3 (Fig. 1).
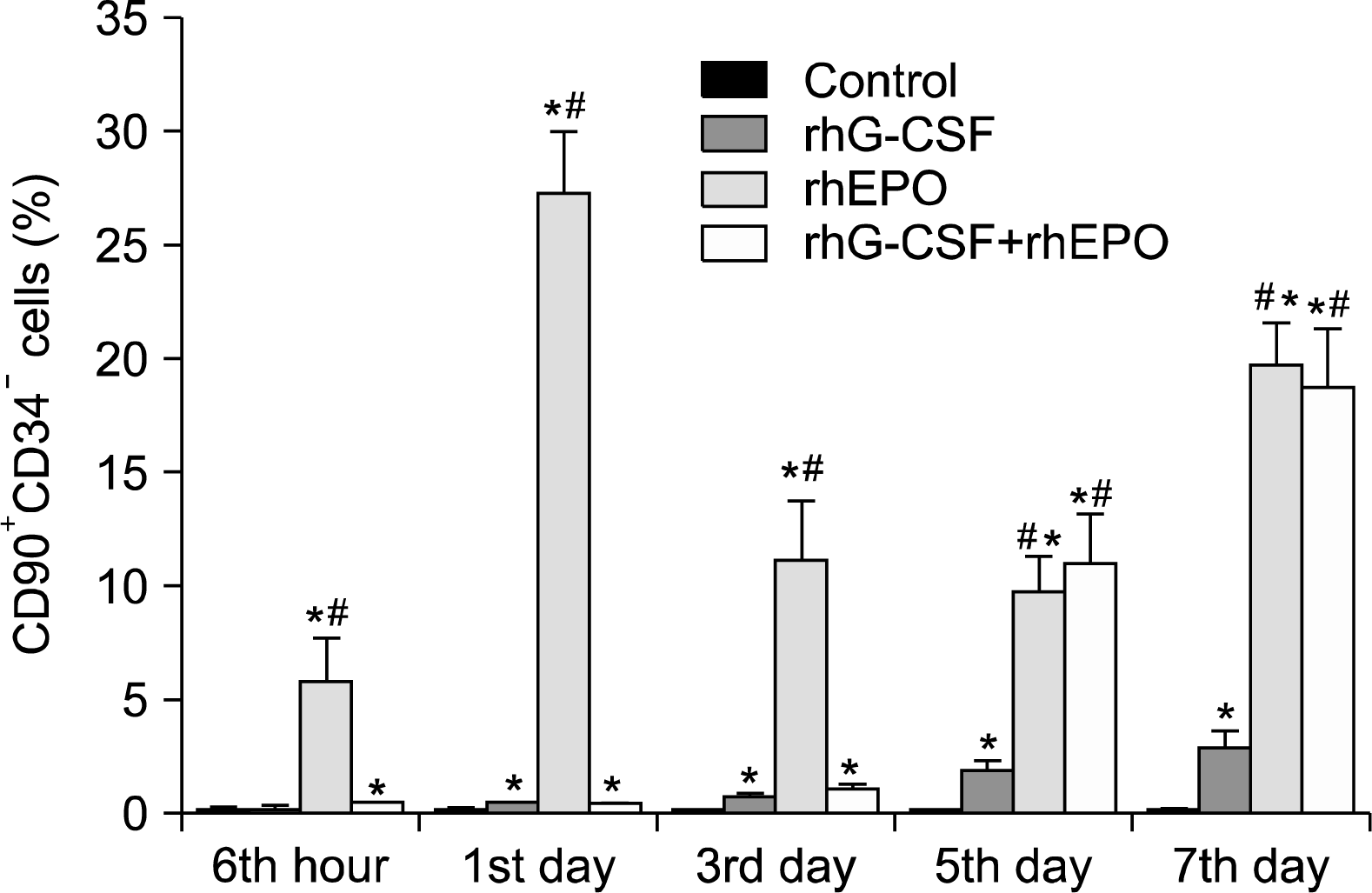
Graft composition during mobilization
To analyze the graft composition in mobilized peripheral blood, the expression of stem cells surface makers: CD34, CD90 and Sca-1 were investigated by RT-PCR. As show in Fig. 2, mRNA expression of CD34, CD90 and Sca-1 at hour 6, day 1 of rhEPO stimulation was significantly higher than adiministration of rhG-CSF and rhG-CSF+rhEPO. At day 3, mRNA expression of CD34 was no significant difference among the three groups; mRNA expression of CD90 of rhEPO or rhG-CSF+rhEPO stimulation was more higher than adiministration of rhGCSF, but there was no significant difference between two groups; Sca-1 mRNA expression in rhEPO group was higher than rhG-CSF or rhG-CSF+rhEPO group (p= 0.003). At day 5 and day 7, the changes of three types mRNA levels showed consistency, rhEPO or rhG-CSF+rhEPO group was higher than rhG-CSF group, but there was no difference between rhEPO and rhG-CSF+rhEPO group (p=0.16).
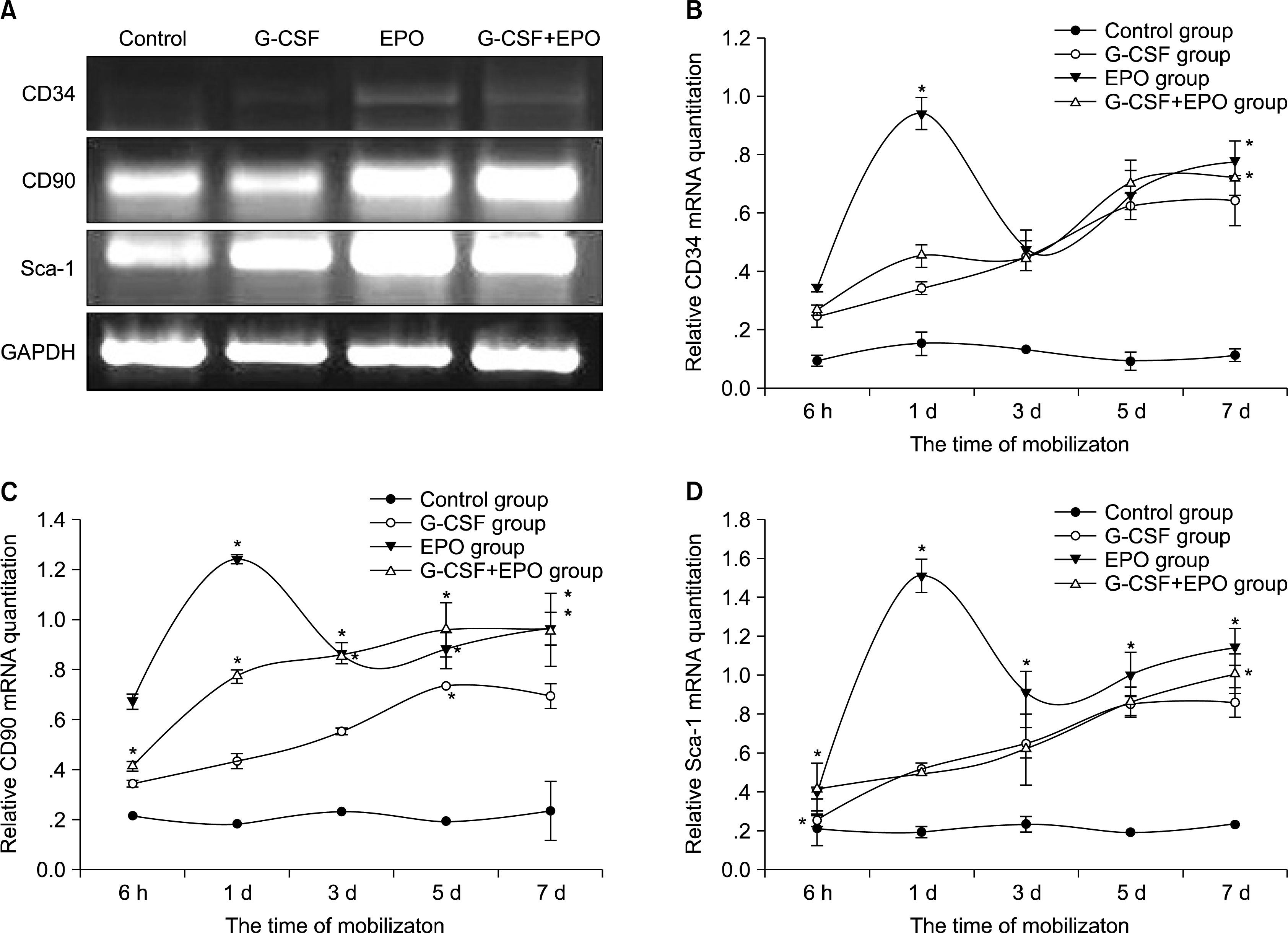 | Fig. 2.RT-PCR analyses of mobilized-peripheral blood for surface makers gene expression. (A) Examples for RT-PCR from four different mobilized groups and on day 7 after mobilization. (B) Qualitative PCR analysis the expression of CD34-mRNA (Mean±S.E.). (C) Qualitative PCR analysis the expression of C90-mRNA (Mean±S.E.). (D) Qualitative PCR analysis the expression of Sca-1-mRNA (Mean±S.E.). *p<0.05, Compared with the G-CSF group.
|
Go to :
Discussion
Recent data from the International Bone Marrow Transplant Registry (IBMTR) show PBSCs are used in more than 90% of autologous stem-cell transplantation and 69% of allogeneic transplantation, and G-CSF is the predominant mobilizer used clinically (1–4). Although the success of G-CSF, there are many areas for improvement that can have significant clinical impact: Meisel et al. (5) showed the survival of G-CSF-based mobilization in pediatric siblings donating for children is shorter than bone marrow transplantation, it mainly beacuse rare HSCs in mobilized peripheral blood; many other studies (6–8) reported PBSCs grafts contained 3∼4 times the dose of CD34+ cells/kg, 10∼50 times total number of T cells, which seriously impact the incidence of graft-versus-host disease, infectious complications and even survival post transplant. These studies raise the issue of what is the optimal graft content.
Adult bone marrow is the primary source of stem cells that regenerate hematopoiesis, and also harbors other stem and progenitor cells including endothelial progenitor cells (EPCs) and mesenchymal stem cells (MSCs). MSCs and EPCs can be used as accessory cells of hematopoietic stem and progenitors cells (HSPCs) in transplantation to promote hematopoietic reconstitution process, reduce complications of post-transplantation and speed up the repair of damaged organs such as heart, liver, kidney and vascular (9, 10). As we konw multiple agents induce PBSCs mobilization, each with different kinetics and efficiencies. Whether combination agents can be developed that differentially mobilize adult stem cells in order to obtain the optimal graft. In our study, we wanted to know whether rhEPO+rhG-CSF can efficiently mobilize bone marrow-derived progenitor cells into peripheral blood and which types of cells are mainly released from bone marrow.
Our data showed that the peripheral blood was collected on hours 6, days 1, 3, 5 and 7, the number of WBC and MNC in peripheral blood gradually increased in cytokines treated mice, and the peak have occurred during the 7 th day after the last adiministration. The high amount of leukocytes indicated a mobilizing effect of cytokines. However, the addition of rhEPO to the treatment protocol did not further increase the mobilizing effect of rhG-CSF as reported by Grassinger et al. (11). The counts of WBC and MNC in KM mice is consistently lower than the study reported (12). This could be explained by the characteristics of agents: electrical conductivity, pH, osmotic pressure, ionic strength, hemolytic agents and concentration used in automatic hematology analyzer are the same as human blood cells, while leukocytes of mice have unique characteristics which are different with human in size, internal structure and so on.
Kang et al. (13) showed G-CSF+EPO enhance the mobilization of EPCs, benefit to angiogenesis and improve the myocardial fuction, this protocol would be a promising option to overcome the significant but limited efficacy of cytokine-based cell therapy for patients with myocardial infarction. This suggest that the graft content has changed with the mobilizing of EPO+G-CSF. According to HSCs in mouse bone marrow have the phenotype of Sca-1+CD34+/−CD45+Lin− and c-kit+CD90low Lin-Sca-1high, and Sca-1, c-kit, CD34 and CD90 are commonly used (14). We found that mRNA expression of CD34, CD90 and Sca-1 increased higher with the stimulation of rhEPO than rhG-CSF alone. This could be explained by more HSCs mobilized by rhEPO.
Although the dosage of CD34+ cells is used in humans to indicate the mobilization of HSPCs, this population is heterogeneous, which containing primitive as well as more mature progenitors. In contrast to humans, CD34+ is a less useful marker in the mouse, and there is good evidence that murine HSCs can also be CD34 negative. For example, Lin−c-Kit+CD34+ is the major phenotype of HSCs in fetal and newborn mice, whereas CD34− HSPCs emerge in 7-week-old mice and gradually increase thereafter, and after 10 weeks there is mainly CD34− HSPCs, only 20% of HSPCs are CD34+ cells (15). Brunner, et al. (16) reported about 70% of “HSC” purified as of Lin−Sca−1+c-Kit+ express CD45 + CD34−, only 17% express CD45+ CD34 + . All these datas show CD34− is the major of HSPCs in mice BM. Our study found the expression of CD34 is lower than CD90 and Sca-1, which may be explained CD34−HSCs are more than CD34+cells. Using FACS analyses that CD90+ CD34− cells increased more significantly after the administration of rhEPO or rhEPO +rhG-CSF than rhG-CSF alone. According to the phenotypes of mice HSCs, our datas suggest more HSCs mobilized by rhEPO.
Currently, many techniques can be used to identify HSPCs, such as the capable of forming colonies, flow cytometric assays and in vivo transplantation. However, the phenotypes and biological characteristics of MSCs remain controversy. The International Society for Cellular Therapy (ISCT) in 2006 have provided a set of standards identifying MSCs: MSCs must express CD105, CD73 and CD90; must lack the expression of hematopoietic antigens like CD45, CD34 and markers for monocytes, macrophages and B cells (17). However, MSCs has a strong heterogeneity. For example, Yamamoto et al. (18) showed the phenotypes of MSCs from adipose tissue in mice as follows: (94.6±2.6)% CD90+, (89.4±4.5)% Sca-1+ and (4.6±2.4)% CD34+. Obviously, some phenotypes of MSCs and HSCs are consistent. In addition, whether MSCs can be mobilized or not has also aroused controversy. While Kassis et al. (19) successfully isolated MSCs by fibrin microspheres in G-CSF mobilized peripheral blood. Therefore, in our experiment, FACS analyses that CD90+CD34− cells increased more significantly after the administration of rhEPO or rhEPO+rhG-CSF than rhG-CSF alone, may also mobilize a certain number of MSCs.
The importance of this study is that exploring rhEPO or combined with rhG-CSF had a strong capacity of mobilization of CD90+CD34−and Sca−1+stem cells, which may enhance the content of HSCs or MSCs or both in graft. As a result of this experiment only identified cells phenotypes and lack of detecting other biological characteristics such as the potential of self-renewal and self-differentiation, it is difficult to accurately definite the CD90+ CD34−and Sca-1+ cells in the mobilized-peripheral blood, which is the next subject we study in the future.
Go to :
 XML Download
XML Download