

 PDF
PDF Citation
Citation Print
Print


Adipogenesis has been defined as the overall outcome from the prenatal and postnatal processes of proliferation, differentiation, and maturation of stem-like (mesodermal) cells to form terminally committed cells of the fat cell lineage, which are capable of conducting lipogenesis and the subsequent storage of visible triacylglycerides (1–5). This understanding of adipogenesis is primarily based upon the use of rodent cell lines or stromal vascular cells and is somewhat flawed, unidirectional and inflexible (1, 4–8). For example, there is little room in the definition for any other cell physiology. In other words, once the cell begins to accumulate lipid and express markers of “differentiation”, the cell is terminally differentiated (reviewed in (4)). Any adipose depot hyperplasia from that point onward is via either spindle-shaped preadipocytes or other stem cells (without lipid) that reside in the depot (1, 2). We suggest that mechanisms to regulate adipose deposition patterns (within all animals) should also include one additional mechanism, the regulation of the mature adipocyte-derived progeny (purified) adipofibroblasts (1, 3–5, 8–22) from different adipose depots.
A historical perspective of the evolution of research in this area, as well as thoughts regarding physiological options for the cell phenotype converting to a proliferative competent cell, have been presented previously (3–5, 7, 8). While once purely speculation, a groundswell of support now exists to suggest that mature adipocytes possess the ability to resume the proliferative program and, even with lipid, can begin to divide (6, 13, 23, 24).
What happens to the progeny cells that result from such dedifferentiation? Are these cells committed to the adipose lineage, or were they ever committed to a single phenotype? Non-clonal cultures of mature adipocytes (11, 16, 25) have suggested that proliferative-competent progeny cells may produce other forms of stem cells (26), while data derived from clonal (purified) cultures (3) of proliferative-capable mature adipocytes have suggested that similar possibilities may exist (6, 24).
We have provided information about the prevalence of mature cells in adipose tissue that possess ability to undergo dedifferentiation, which is a significant number, and we have defined specific methods to isolate and propagate clonal cultures of these cells (3). Our studies have shown that purified cultures (3) of mature adipocytes behave quite differently than batch cultures of mature adipocytes. First, the time required for mature adipocyte dedifferentiation is substantially longer than that observed in non-clonal cultures, even if exposed to similar in vitro conditions (3, 24). Second, proliferation dynamics (24) and handling of the lipid load appears to be different depending on the animal species from which the mature adipocytes were obtained. For example, clonal cultures of beef-derived mature adipocytes resume proliferation in both a symmetric (equal amounts of parental cell lipid retained by both progeny cells) or an asymmetric (unequal amounts of lipid retained by both progeny cells) manner (3–5). In none of the cultures examined, did the beef-derived adipocyte expel lipid from the cell. Moreover, in the purified bovine-derived system, the progeny adipocytes appear to be proliferative-competent (4, 24) but undergo redifferentiation in a different manner (24) and also express some differences in intrinsic programming than other adipogenesis cell systems (27, 28).
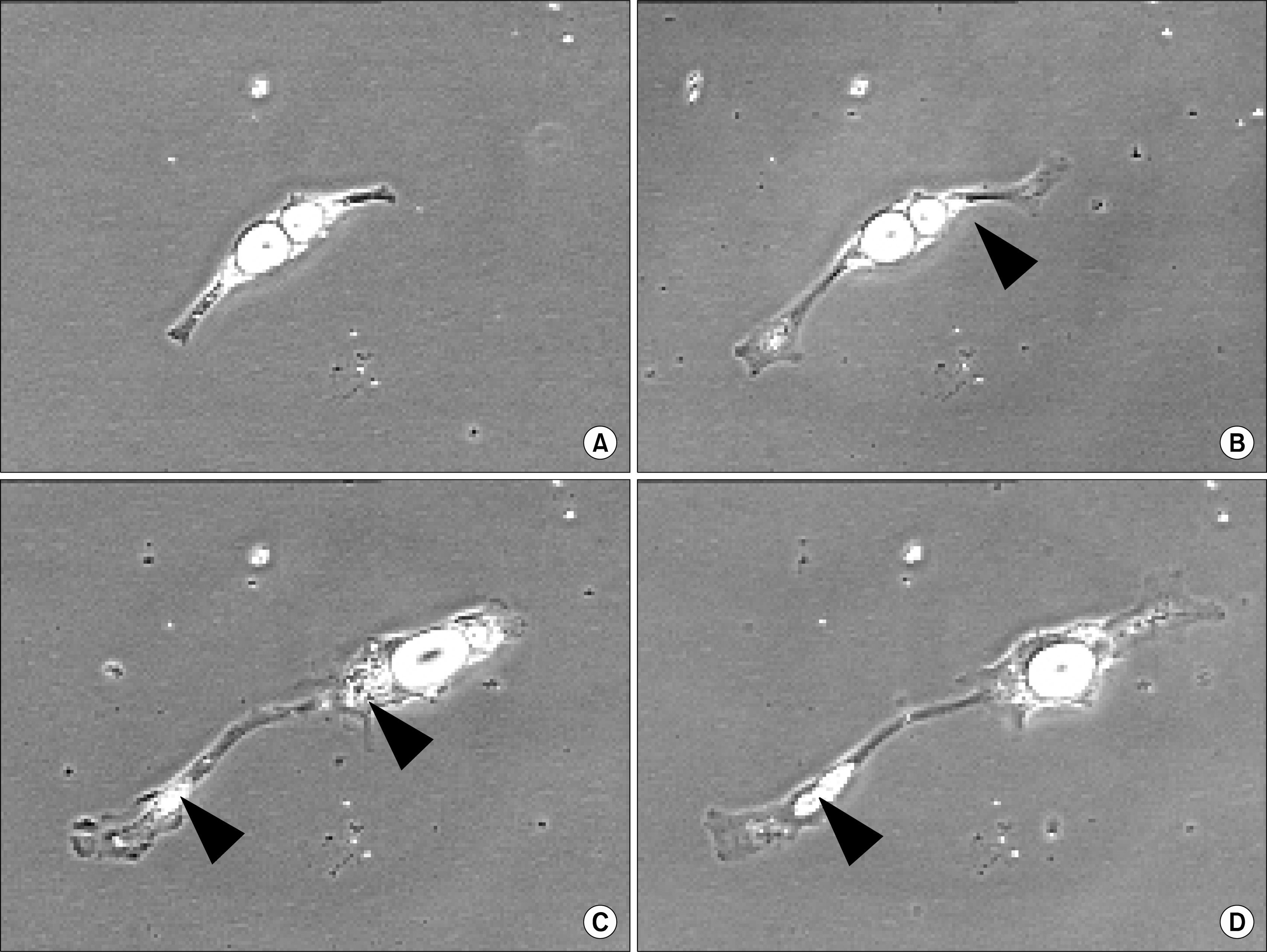
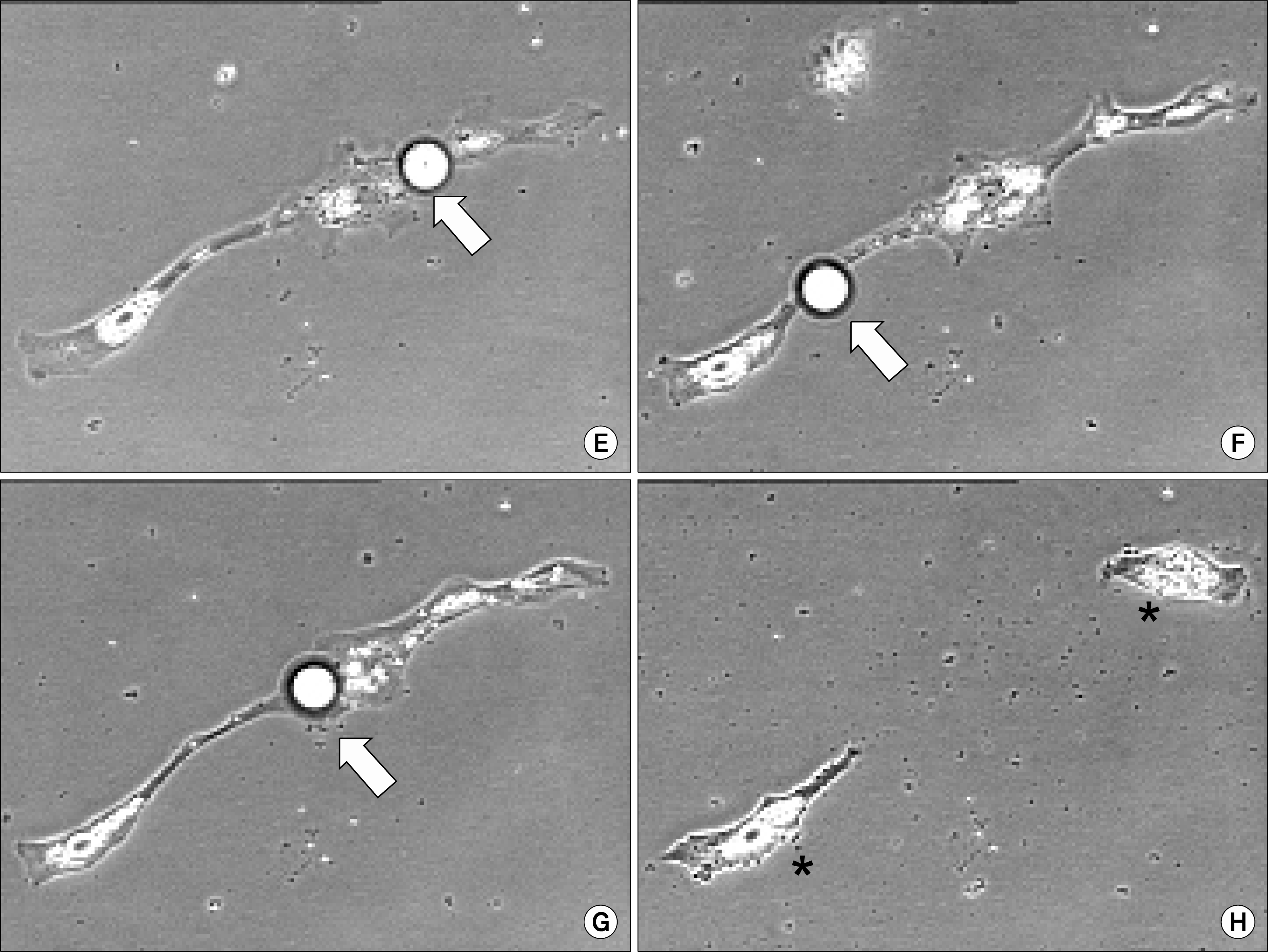
When pigs were used to obtain mature adipocytes for evaluation of this dedifferentiation phenomenon, something quite different was observed (Fig. 1). First, the initial ceiling cultures of pig-derived adipocytes provided qualitatively more mature adipocytes than in the beef system. Second, isolated mature pig-derived adipocytes seemed to include cells with all levels of lipid filling. Third, prior to division, clonal cultures (3) of pig-derived adipocytes were shown to eliminate substantial lipid from the cell, resulting in only an asymmetric cell division. Similar observations were not observed in the beef-derived mature adipocyte cultures (3–5) and suggest that the lipid handling machinery is different and the proliferative machinery of pig-derived adipocytes require less cytoplasmic lipid loads in order to complete the process of cell division.
In part, these types of data offer new insight into species differences in the process of reverse adipogenesis (or de-differentiation), as well as processing of lipid prior to resumption of cell division. This research with clonal populations of proliferative-competent pig-derived adipocytes may prove to be an emerging area of growth physiology that may modify present thinking about adipose tissue renewal capabilities in animals (within and between species). Moreover, it presents a model system for the study of lipid metabolism and mobilization. As adipose tissue provides a reservoir for obtaining putative stem cells (24), knowledge of cellular lipid processing dynamics may aid in our generation of viable stem cells.
 XML Download
XML Download