

 PDF
PDF Citation
Citation Print
Print


Introduction
Peroxisome proliferator-activated receptors (PPARs) are ligand-activated transcription factors that belong to the nuclear receptor superfamily (1). They have been identified in a variety of different species including the xenopus, mouse, rat, and humans (2). After the isolation of PPARα (NR1C1), as the receptor mediating peroxisome proliferation in rodent hepatocytes in 1990 (3), two related isotypes, PPAR δ (NR1C2; also called PPARβ/δ) and PPARδ (NR1C3) located on chromosomes 15, 17 and 6 in the mouse and chromosomes 22, 6 and 3 in human, were identified (4, 5). Once activated by their respective ligands, the PPARs control the transcription rate of a large panel of genes implicated in various physiological functions, including adipogenesis, lipid and glucose homeostasis, inflammation, cell proliferation, differentiation, and carcinogenesis (2, 6–8). PPARs heterodimerize with the retinoid X receptors and modulate gene expression of target genes containing peroxisome proliferators-responsive elements in response to ligand activation (9, 10).
The three isoforms of PPARs display distinct physiological and pharmacological activity that is dependent on their target genes and their tissue distribution (11, 12). PPARα, activated by polyunsaturated fatty acids and leukotriene B4, is expressed in tissues with high fatty acid catabolism such as the liver, heart, brown adipose tissues, kidney, and intestine. PPARγ, mainly expressed in adipocytes, macrophages, placenta, and other tissues, is activated by specific fatty acid metabolites, such as 15-deoxyprostaglandin J2 (15d-PGJ2), and by thiazolidinediones. Both PPARα and PPARγ response genes are involved in lipid homeostasis. Therefore, it is not surprising that the main functions of PPARα and PPARγ are related to glucose and lipid homeostasis (13–15). On the other hand, the ubiquitous distribution of PPARδ, (although gut, kidney, and heart express higher levels than other tissues) makes it difficult to associate PPARδ with a specific biological function (14). Although PPARδ is the least studied PPAR, it has been reported that PPARδ is associated with a diverse range of functions. Indeed, PPARδ participates in many biological processes, including lipid and glucose metabolism (16–19), epidermal maturation and wound healing (20–22), muscle development and function (23–25), tumorigenesis (26–30), inflammation (31, 32), and cytoprotection (33). In addition, combining multiple research approaches, PPARδ has been implicated in embryo development including stem/progenitor cell proliferation (34, 35), embryo implantation (13, 36, 37), embryo organogenesis, and diabetic embryopathy (38, 39). Therefore, this review will focus on the role of PPARδ in the ES cells.
Go to : 
Physiological functions of PPARδ
PPARδ has been the most elusive among the three PPAR subtypes. Due to its broad tissue distribution, it is difficult to identify a specific function for this receptor. PPARδ has a broad expression pattern in adult animals, and is detected very early during embryogenesis (40). Several studies have shown that PPARδ is activated by a large variety of ligands and is implicated in the developmental and metabolic regulation of several tissues. PPARδ activators include fatty acids (41, 42), triglycerides (43), the cyclooxygenase (COX) product, prostacyclin (42), the COX/prostacyclin synthase derived endocannabinoid metabolites (44), and all transretinoid acid (45). A number of synthetic PPARδ ligands have been described including GW0742X, GW2433, GW9578, L-783, 483, GW501516, L-796,449, L-165,041, and compound F (46–48). In addition, GW501516 and GW0742 activate PPARδ at very low concentrations both in vivo and in vitro with a 1,000-fold selectivity over the other PPAR sub-types (49).
A mouse knockout model, although difficult to generate due to highly penetrant lethality (21, 50), indicated a role for PPARδ during embryo implantation, as well as in myelination of the brain, lipid metabolism and adiposity, and epidermal cell proliferation (37, 50, 51). In addition, PPARδ has been linked to cell differentiation, inflammation, cell motility, and cell growth (9, 52). Recent studies suggest that PPARδ plays a role in cell growth. For example, the PPARδ expression is increased in colorectal cancer cells compared with normal colon epithelial cells (30, 53). Treatment of GW501516, a PPARδ ligand, increased the number and size of intestinal polyps (29). In addition, PPARδ has been implicated in the growth of several other cell types including vascular smooth muscle cells (54), preadipocytes (17, 55) and epithelial cells (9). It has recently become clear that PPARδ has a function in epithelial tissues; however, this role continues to be debated in reports with inconsistent findings. Indeed, some reports suggest that ligand activation of PPARδ potentiates cell growth (56), whereas other reports suggest that ligand activation of PPARδ attenuates cell growth (57). Activation of PPARδ by its agonist increases COX2 expression in human hepatocellular carcinoma cells (58). In addition, PPARδ has been shown to mediate prostaglandin E2 (PGE2)-induced cell growth (59). More importantly, several reports have suggested that PPARδ-mediated cell growth is induced by COX/prostaglandin (PG) signal pathways. Consequently, the interplay between the PPAR δ and cytosolic phospholipase A2 (cPLA2)/COX2/PGE2 signaling pathways acts as a positive regulator in cell growth.
Apart from cell growth, the fact that PPARδ plays an important role in embryonic development is supported by the observation that most PPARδ-deficient mice die early during embryonic development due to a placental defect. A recent study showed that the genetic loss of PPARδ signaling does not influence ovulation, fertilization or preimplantation (60). Mouse embryos express PPARδ detectable at the two-cell stage (61) or eight-cell stage (36), and throughout the preimplantation period. Mouse blastocysts also express PPARδ in the inner cell mass and the trophectoderm. On the embryonic cell level, prostacyclin or the PPARδ agonist increased the embryonic cell mass, indicating that PPARδ is essential for embryo development, blastocyst hatching and implantation (36).
Go to :
PPARδ and embryonic stem (ES) cells
The elucidation of PPARδ modulation of ES cells will provide new insights into embryonic development. In addition, research on how normal embryonic development is regulated will provide new clues as to how to maintain stem cells in culture. The description of the interaction among signal molecules is a key to our fundamental understanding of stem cell proliferation and its translation into therapeutic strategies. However, information regarding the potential role of PPARδ in physiological and/or developmental processes is very limited, although PPARδ is widely expressed in embryonic tissue. There is evidence that PPARδ can modulate stem and progenitor cell expansion and a differentiated phenotype. In neural stem cells, PPARδ contributes to the maintenance of the un-differentiated and proliferative status, by regulating both the genes involved in cell cycle control, as observed in other cell types (54, 62, 63), and inhibiting the activity of the other PPARs, which may be involved in cellular differentiation (64–66). In addition, the PPARδ agonist GW 501516 has been shown to be a promoter involved in the development of adenosquamous carcinomas with high expression of stem cell markers CK19 and Notch1, as well as Proliferin, a growth factor that mediates many of the effects of the stem cell markers such as Musashi1, in mammary cells (67). PPARδ is expressed in the crypt cells of the small intestine and negatively regulates Hedgehog signaling to block differentiation (68), a process that would be expected to promote transformation. The association of Wnt activation with stem cell expansion, activation of β-catenin/T-cell factor (TCF) signaling by 3-phosphoinositide-dependent protein kinase 1 (PDK1) and the identification of PPARδ responsive genes suggest a common mechanism for the tumor promoting action of PPAR δ agonists that may involve stem and progenitor cell proliferation (35).
For successful implantation and pregnancy, recent evidence suggests that the implantation timing of PG signaling resulting from cPLA2, COX2 or lysophosphatidic acid receptor 3 plays an important role in the subsequent developmental processes (69–72). However, the underlying mechanism and the molecular link between the critical steps are still unclear. A previous report provided evidence that PPARδ serves as a molecular link that coordinates multiple signaling pathways in mouse ES cell proliferation allowing their self-renewal (Fig. 1) (34). In that study, high glucose (25 mM) increased PPARδ gene expression rather than PPARα or PPARγ in the ES cells. In addition, the PPARδ agonist, L-165,041 increased ES cell proliferation, but the PPARδ antagonist, GW9662 or the PPARδ specific small interfering RNAs inhibited the effects of high glucose. Moreover, high glucose increased COX2 and PGE2 synthesis activating PPARδ, which increases cell cycle regulatory protein expression such as cyclins and cyclin dependent kinases (CDKs). It has been consistently shown that COX2-derived PGE2 and PGI2 mediate their function via PPARδ receptors during the early steps of decidualization in mice (73). Although the exact mechanisms involved in COX2-derived PG activation of PPARδ has not been completely elucidated, previous reports have suggested that the activity of PGE2 is mediated by possible activation of the EP2 receptor, which increases the cAMP levels (74–76) and PPARδ receptors. Previous work has shown that embryos from streptozotocin (STZ)-induced diabetic rats have diminished PGE2 content, although they can produce PGE2 in large amounts (77). One can speculate that arachidonic acid (AA) might be depleted if PGE2 generation and release is increased in the diabetic embryo in order to maintain the intracellular PGE2 levels (39). In addition, it has been suggested that ligand activation of PPARδ induces the expression of COX2 (56), which could theoretically promote cell growth and inhibit apoptosis through mechanisms that involve the production of prostaglandins. These data raise the possibility that impaired activation of PPARδ may alter the lipid signaling required for normal self-renewal of ES cells, which raises that possibility that PPARδ might be a putative target for the maintenance of ES cell characteristics. The results of a previous study suggested that the loss of PPARδ leads to reduction of the phosphorylation status of Akt and STAT3 in the trophoblast (60). Akt is a down-stream pathway of PPARδ signaling that is active during cell proliferation and survival (60); it was observed that PPARδ null mice were not able to progress through the normal developmental steps. The PI3K/Akt signaling pathway also has been implicated in ES cell self-renewal in studies of ES cells without PTEN (78). Thus, this observation supports the participation of PPARδ in ES cell proliferation and maintenance of self-renewal. In addition, several groups have shown that STAT3 is an important signal transducer and activator in the maintenance of pluripotency and the propagation of mouse ES cells (79–81). Although STAT3 phosphorylation with LIF was not influenced by the PPARδ −/− trophoblast, the STAT3 was not phosphorylated by the PPARδ agonist. This suggests that PPARδ activity not only plays a role in normal development but also is involved in ES cell proliferation and self-renewal. It has been shown that STAT3 could direct the expression of key regulators of the mitotic cycle in ES cells and stimulates their entry into the S phase (79). Thus, it is possible that PPARδ plays a role in inducing cell cycle molecules that are involved in ES cell proliferation. Gene expression profiling experiments will help gain insights into the mechanisms involved in PPARδ activity during the process of self-renewal.
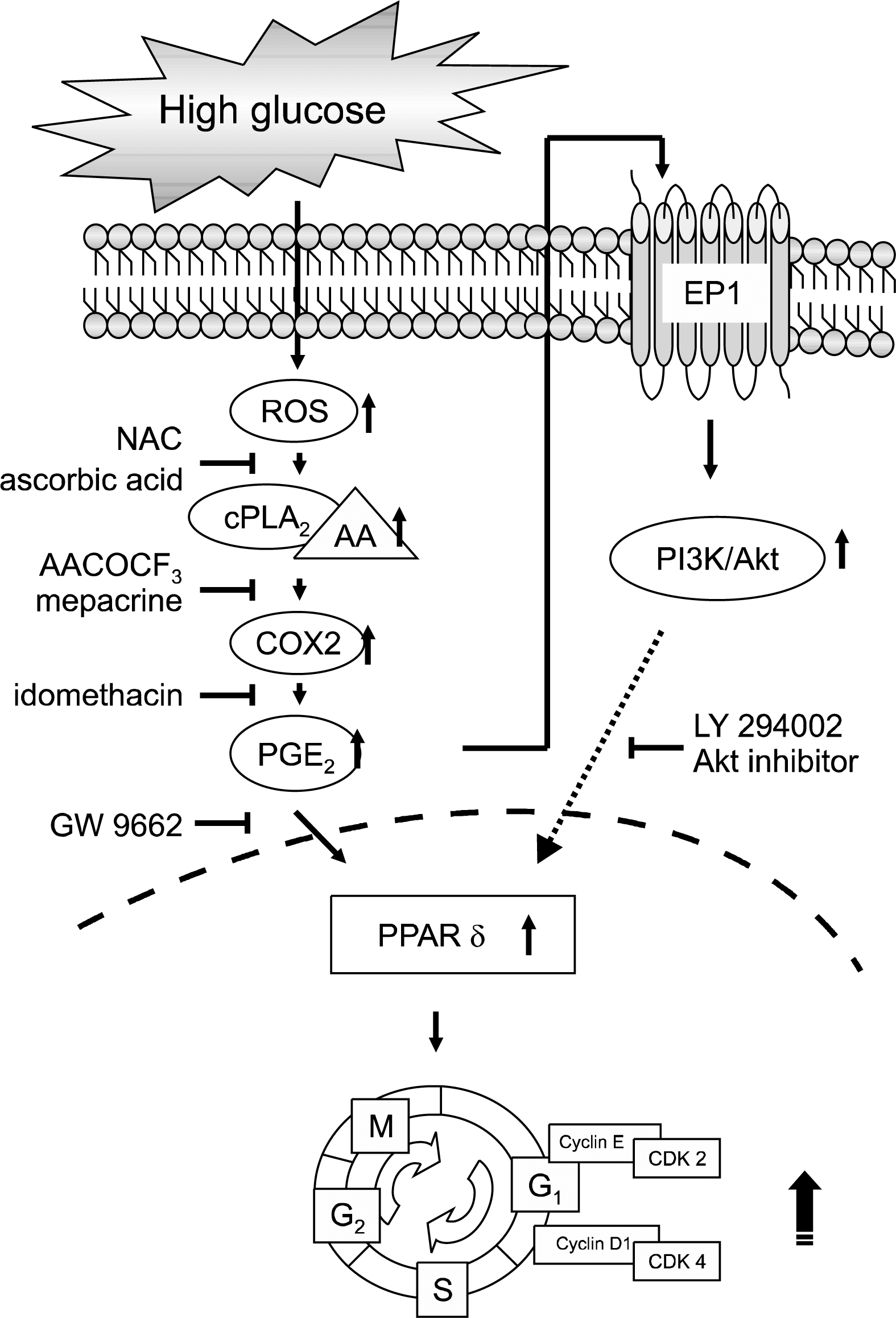 | Fig. 1.The hypothesized model for the signal pathways involved in high glucose-induced mouse ES cell proliferation. High glucose increased PGE2 synthesis, which is controlled by the coupled activation of cPLA2/AA and COX2 via ROS. After PGE2 is released into the extracellular space, it binds to the membrane coupled EP receptors. This effect is mediated, at least in part, by the activation of PI3K/Akt. Finally, these molecules may induce PPARδ, which increases cell cycle regulatory protein expression levels. Abbreviations: AA, arachidonic acid; CDK, cyclin-dependent kinase; COX2, cyclooxygenase-2; cPLA2, cytosolic phospholipase A2; NAC, N-acetylcysteine; PGE2, prostaglandin E2; PI3K, phosphoinositide 3-kinase; PPAR, peroxisome proliferator activated receptor; ROS, reactive oxygen species (34).
|
Go to :
Conclusions
Over the past few years, knowledge of the physiological activity of PPARδ has expanded. The study of PPARδ characteristics has added to improve understanding of cell physiology. PPARδ has been implicated in many cell processes, from the embryo to adult cells, and from cell proliferation to cell differentiation; it has been shown to be crucial for energy homeostasis. Until recently, because the function of PPARδ remained elusive, the therapeutic potential of PPARδ agonists for lipid and glucose metabolism, embryo development and wound healing has been tested in mice only. Our current understanding of PPAR δ has demonstrated that PPARδ also plays a critical role in ES cell proliferation. Future studies will likely clarify the physiological role of PPARδ in ES cell growth and differentiation.
Go to :
 XML Download
XML Download