

 PDF
PDF Citation
Citation Print
Print


Introduction
Bone marrow (BM), as an ample adult stem cell source, contains at least three distinct stem cell species, namely, mesenchymal stem cells (MSCs), hematopoietic stem cells (HSCs) and multipotent adult progenitor cells (MAPCs) (1). Among them, MSCs have been most extensively investigated with respect to tissue engineering and cell therapy. Since they can be conveniently isolated from various human tissues, expanded in vivo to a great extent, and specifically induced to differentiate into cells of multiple line-ages, they are regarded as one of ideal sources for stem cell-based regenerative medicine (2).
Within BM, MSCs continuously proliferate and generate differentiated cells of a mesodermal lineage, such as osteoblasts, chondrocytes, adipocytes and myoblasts, thus serving as a bona-fide cell reservoir for connective tissues (3). Besides this tissue regenerative function, MSCs play a central role in a microenvironment or niche that controls the localization, self-renewal and differentiation of HSCs (4–7) and modulates the cellular functions of a variety of immune cells including, B and T lymphocytes, natural killer cells, monocytes and dendritic cells (8–13).
Presumably, the HSC niche is operated by a complex interplay of short- and long-range signaling that may entail a wide spectrum of molecular mediators, including soluble cytokines and growth factors, as well as diverse molecules on the plasma membrane and the extracellular matrix (ECM) (5–7). In the past year considerable efforts have been directed to identifying the key molecular players of the niche, leading to the findings that the physical contact of the HSCs with neighboring osteoblasts regulates the developmental stage and size of their niche (14, 15), and that such processes are mediated, alone or in combination, by a number of diverse signaling pathways, such as bone morphogenic protein (BMP) (15), Notch (16), Tie-2/Angiopoietin-1 (17) and Wnt pathways (18, 19). On the other hand, the result from a recent gene expression profiling study has indicated that the molecular context of the niche may be much more complex than originally anticipated (20).
In particular, MSCs embedded in the niche have been known to play crucial roles in the regulation and fine-tuning of the HSC development, primarily through a collective action of as-yet-unidentified soluble mediators (4–6). Of prime importance in understanding the niche at the molecular level is, therefore, to characterize the trophic nature of MSCs in a comprehensive manner. For this, a number of cytokine gene expression profiling studies of BM-derived MSCs have been performed (21–25), but the compilation of the data has generated neither a consistent nor comprehensive expression profile. In this study, we attempted to determine a comprehensive cytokine secretion profile of MSCs with the use of a wide-spectrum human cytokine antibody array. This profile was then compared to DNA microarray-based gene expression data recently obtained using the same cell populations (26).
Go to : 
Materials and Methods
Cell culture of human MSCs
Human BM-derived MSC samples were purchased (Cambrex BioScience, Baltimore, MD), all of which exhibited an immunophenotype of CD105+ CD166+ CD29+ CD44+ CD14− CD34− CD45−, and in vitro mesengenic differentiation potential. And full-term umbilical cord blood (UCB) sample was collected with mother’s consent and the protocol approved by internal review board of our institutions. Mononuclear cell (MNC) fraction was separated from UCB using Ficoll-Paque PLUS (Amersham Biosciences, Uppsala, Sweden) and MSCs were derived by continuous subculture. All these cells were further cultured as monolayers in culture media consisting of Low Glucose Dulbecco’s Modified Eagle Medium (LG-DMEM, Life Technologies, Gaithersburg, MD), 20% fetal bovine serum (FBS, RH Biosciences, Lenexa, KS), 2 mM L-glutamine, 1 mM sodium pyruvate, and 1% antibiotics/an-timycotics (Life Technologies, Gaithersburg, MD) comprising of 100 U/ml penicillin, 100μg/ml streptomycin, and 25μg/ml amphotericin B. The cells were detached with with 0.1% trypsin-EDTA when they reached their 60∼70% confluence and re-plated at a density of 2×103/cm2 in Falcon culture flasks. For this study, the cells and culture media were collected at the early (3rd) passage, and total RNA was separately prepared from each cell sample using RNeasy Mini Isolation Kit (Qiagen, Valencia, CA).
Cytokine secretion profiling by cytokine antibody array
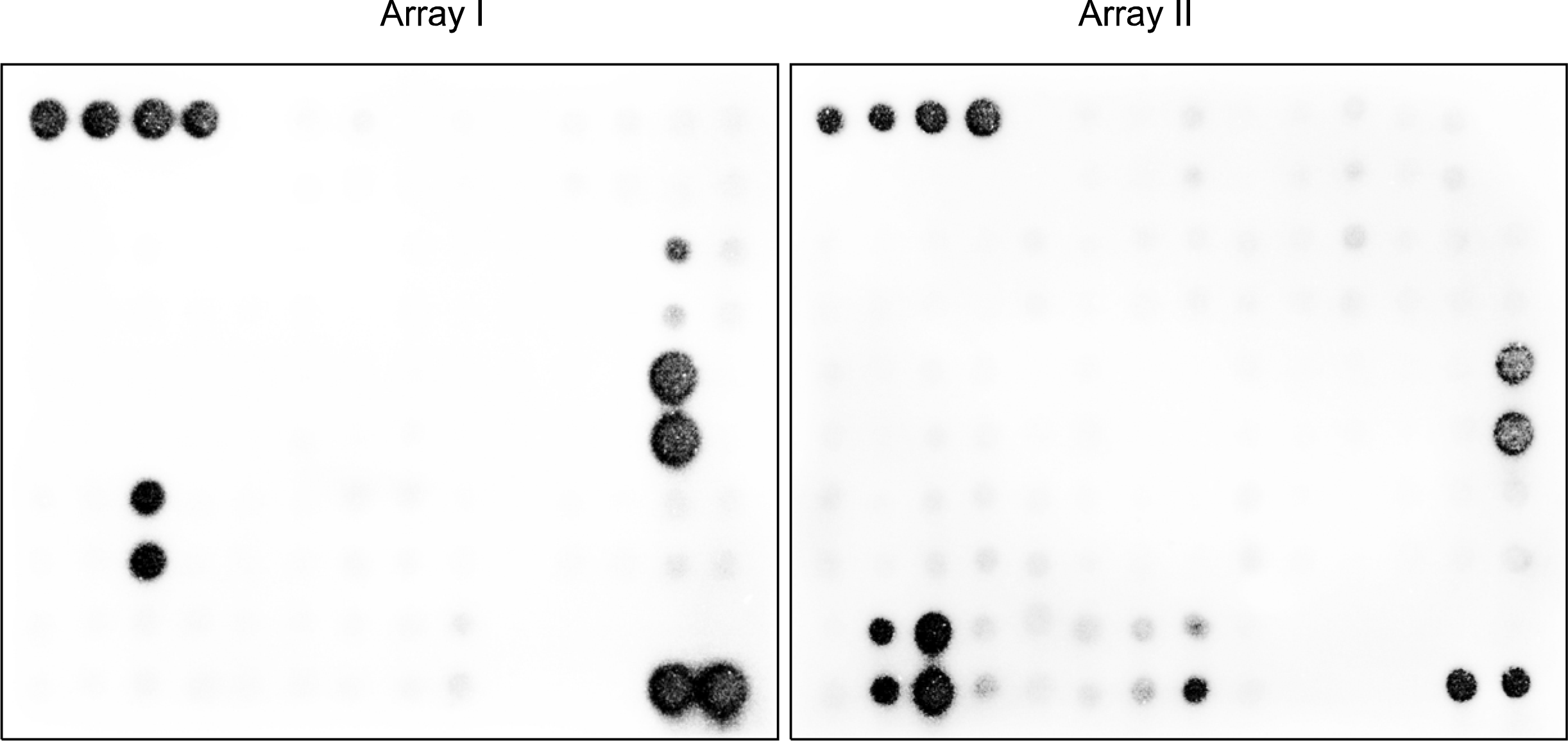
Human Cytokine Arrays VI and VII (Ray Biotech Inc, Norcross, GA) consisting of total 120 different cytokine and chemokine antibodies spotted in duplicate onto two membranes were used (Fig. 1A). Array membranes, each in separate wells of provided six-well plate, were incubated for 10 or 90 min in 2 ml of blocking buffer consisting of 10% bovine serum albumin in Tris buffered saline, further incubated for 10 min or 90 min in a shaker at room temperature with 2 ml of 4 or 20-fold diluted incubated or fresh culture medium. After the membranes were thoroughly washed, 2μl of biotin-conjugated antibodies at 1: 250-fold dilution was added to each membrane, and the mixture was incubated on a shaker for 10 or 90 min at room temperature. Following the wash, the membranes were incubated with a 1:40,000 dilution of streptavidin-conjugated peroxidase for 45 min at room temperature. Proteins were detected by enhanced chemiluminescence and signals were captured on X-ray films. The exposure time was varied from 1 min to 1 hr to optimize the signal-to-background ratio of the image. Arrays images were photographed and processed with Chemi Doc XRS (Bio-Rad Laboratories, Hercules, CA).
 | Fig. 1.Control cytokine profiling of fresh culture medium. (A) Layout of antibody arrays and abbreviated names for 120 different cytokine probes. The full names of cytokines are indicated in Table 1. POS; positive control, BNK; blank, NEG; negative control. Membrane hybridization signals of (B) the antibody array incubated for 90 min with the 4-fold diluted culture medium and (C) the array incubated for 10 min with the 20-fold diluted medium. The latter hybridization condition was used for further cytokine secretion profiling studies of human MSCs. Table 1.A list of 120 cytokine probes implemented in the antibody array. Their respective full and systematic names are provided with the antibody array-generated spot intensities as well as DNA microarray-based gene expression intensity (26)
|
Semi-Quantification of cytokine antibody array data
Chemiluminescent cytokine array data were semi-quantified with Quantity One software (Bio-Rad Laboratories). The intensity of each cytokine spot was measured on the basis of gray-scale levels. For every spot, the net density gray level was determined by subtracting the average gray level of four negative controls from the measured density gray levels. The density gray level of six positive controls was used to normalize the results from different membranes. The density for each cytokine was then averaged over duplicated spot signals.
RT-PCR analysis
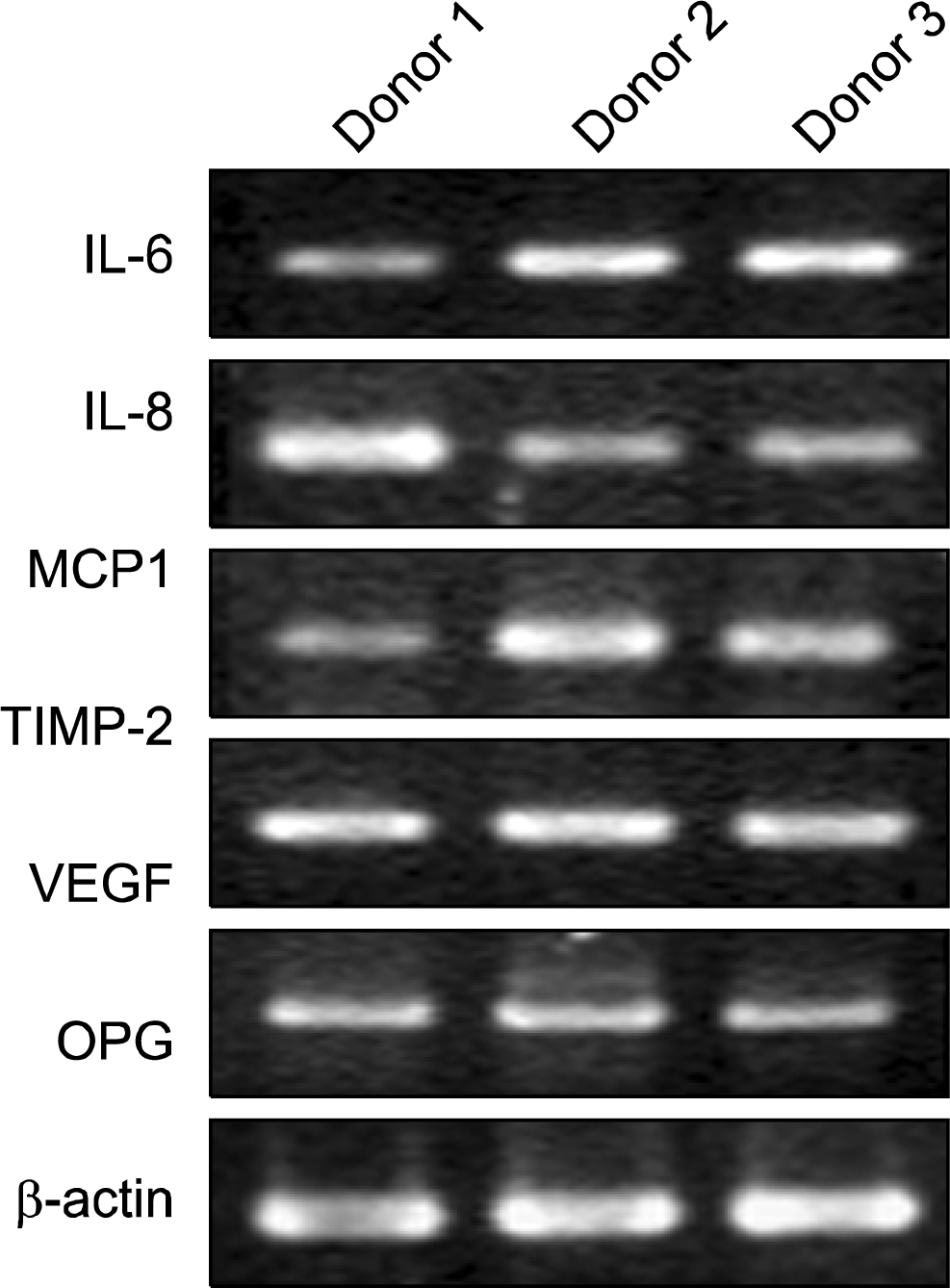
To confirm the cytokine secretion profile determined by antibody arrays, the gene expression levels for abundantly secreted cytokines were measured by RT-PCR, using total RNAs derived from three different MSC samples. Messenger RNA of β-actin served as an internal control. The sense and antisense primers used for each gene were as follows: IL-6, sense 5′-TAC CCC CAG GAG AAG ATT CC-3′ and antisense 5′-CAG GGG TGG TTA TTG CAT CT-3′; IL-8, sense 5′-TCT GCA GCT CTG TGT GAA GG-3′ and antisense 5′-TGA ATT CTC AGC CCT CTT CAA-3′; MCP-1, sense 5′-TCT GTG CCT GCT GCT CAT AG-3′ and antisense 5′-GAG TTT GGG TTT GCT TGT CC-3′; TIMP-2, sense 5′-GGA AGT GGA CTC TGG AAA CG-3′ and antisense 5′-GTC GAG AAA CTC CTG CTT GG-3′; VEGF, sense 5′-GGC AGA AGG AGG AGG GAC AGA ATC-3′ and antisense 5′-CAT TTA CAC GTC TGC GGA TCT TGT-3′; OPG, sense 5′-TCC TGG CAC CTA CCT AAA ACA GCA-3′ and antisense 5′-CTA CAC TCT CGG CAT TCA CTT TGG-3′;β-actin, sense 5′-AGA AAA TCT GGC ACC ACA CC-3′ and antisense 5′-CCA TCT CTT GCT CGA AGT CC. The thermocycler conditions were initial denaturation at 95°C for 5 minutes; 35 cycles of 95°C for 30 seconds, 57°C for 30 seconds, 72°C for 30 second; and finally 72°C for 7 minutes. The amplified PCR products were resolved in 1.5% agarose gel, stained with eithidium bromide, visualized and photographed with Chemi Doc XRS (Bio-Rad Laboratories).
Go to :
Results
FBS contains numerous cytokines that are reactive to human antibodies
The cytokine antibody array incubated for 90 min with the 4-fold diluted culture medium as suggested in the manufacturer’s protocol yielded a number of spots whose intensities were significantly stronger than the back-ground level (Fig. 1B), indicating that FBS contains numerous soluble factors that could be cross-reactive to human antibodies. However, when the incubation time was reduced to 10 min with 20-fold diluted medium, the hybridization signals for most of these serum proteins were minimized to background (Fig. 1C). This hybridization condition was used for further cytokine secretion profiling studies of BM-derived MSCs.
The cytokine secretion profile of BM-derived MSCs is donor-independent
Next in order to investigate whether the cytokine secretion profile of BM-derived MSCs was dependent on individuality factors such as gender, race and age, we prepared MSC populations derived from three different donors of diverse genetic backgrounds. Since the transcriptomes of these populations were found to be highly similar to each other, with the correlation coefficients ranging from 0.943 to 0.972, it was expected that their variation in cytokine expression and secretion patterns if any would be minimal. As it turned out, the three MSC populations exhibited a common hybridization pattern on cytokine antibody arrays (Fig. 2), suggesting that the cytokine secretion profile of the cells might be donor-independent under the normal condition. This consistent cytokine secretion profile in MSCs was believed to be essential for performing their intrinsic functionalities.

Only few cytokines were secreted at a sufficiently high level from MSCs
As being common to three cell populations, the arrays featured a single predominant hybridization signal for IL-6, and moderate elevated signals for five cytokines, IL-8, TIMP-2, MCP-1, VEGF and OPG (Fig. 2). To facilitate further analyses, all spots in the arrays were semi-quantified and their net intensity values were obtained by subtracting the background intensity. The intensity for a specific cytokine was then computed by averaging over duplicated spots and the three membranes (Table 1). Among 120 cytokines implemented in the array, only six cytokines have the spot intensity above 50 while the intensity for a majority of other cytokines remained at the background level. This observation indicates that only a small subset of cytokines may be abundantly secreted from human BM-derived MSCs. It was worthwhile to note that a number of cytokines, previously known as MSC-related cytokines, such as MCSF, GM-CSF, G-CSF, LIF, IL-11, IL-7, TPO, SCF, HGF, SDF-1 and TGF-β were detected with no measurable intensity.
RT-PCR but not DNA microarray analysis confirms gene expressions for secreted cytokines
To validate the cytokine secretion profile at the transcriptional level, we analyzed gene expressions of highly secreted six cytokines by RT-PCR. The result showed that the cells from all three donors expressed the messenger RNAs of these cytokines at a similar and relatively high level (Fig. 3), indicating that these molecules might be consistently up-regulated at both the gene expression and protein synthesis/secretion levels. However, when all cytokines implemented in antibody arrays were compared to recent DNA microarray-based gene expression data (26), a poor agreement was yielded. Among above-mentioned six cytokines, only IL-6, TIMP2 and VEGF were found to have sufficiently high gene expression intensities, but the genes for the other three cytokines appeared to have minimal or negligible gene expression intensities. Moreover, a number of molecules that were higher in gene expression intensity, such as IGFBP4, IGFBP6, MIF and TIMP1, turned out to be secreted at the minimal level. This result indicated a poor agreement between the antibody array-generated secretion profile of the cytokines and DNA microarray-based gene expression data. This gene-to-protein disparity, however, appeared not to be unusual as reported elsewhere (27, 28). between the present secretion profile of cytokines and its gene expression profile.
Comparison of cytokine secretion profile between BM and UCB-derived MSCs
Finally, in order to investigate whether the current cytokine secretion profile could be generalized to all MSC populations from diverse cell sources, we derived a MSC population from a UCB sample and determined its cytokine secretion profile by the identical approach. The result indicated that the profile of UCB-derived MSCs shared a high similarity with that of BM-derived cells. However, a noticeable difference was found in the secretion level of IL-8 (Fig. 4), that appeared to be more up-regulated in the UCB-derived cells than in the adult counterpart. In addition, the UCB-derived MSCs, but not BM-derived cells, were found to secrete cytokines of the IGBP class cytokines slightly in a donor-dependent manner.
Go to :
Discussion
Cytokines or growth factors are decision-makers of cell fates toward survival versus death and interaction versus protection, etc. Stem cells are no exception to this rule; it is now well established that the core stem cell properties, such as self-renewal, lineage-specific differentiation and tissue engraftment, are largely influenced by the local microenvironment (or niche) and soluble molecular signals. A multitude of cytokines and growth factors may form a complex cytokine network, which confers not only considerable stability and flexibility to the cells, but also rapid amplification of response against a particular stimulus. Thus, the disequilibrium of this network, usually caused by either progressive diseases or bacterial/viral infections, may distort the intrinsic biological functions of stem cells. The cytokine network is best studied in adult BM, in which a plethora of molecular mediators regulates the development of HSCs and also possibly other stem or progenitor cells. It has been long known that a variety of soluble molecules secreted from a neighboring MSC population largely contribute to this network, but their molecular constituents and respective roles have not been well delineated. Therefore, a characterization of the trophic nature of MSCs at the molecular level is regarded as an inevitable step for understanding of the complex cytokine network in BM.
In this study, we have attempted to determine the cytokine secretion profile of human BM-derived MSCs with using an antibody array that could analyze simultaneously protein expressions of up to 120 human cytokines. The overall spectrum of this array is broad enough to encompass most of MSC-secreted cytokines, such as M-CSF, GM-CSF, G-CSF, IL-6, IL-11, IL-7, TPO, SCF, FLT3L, HGF, IL-8, SDF-1, MCP-1, TGF-β, IGF-1, PDGF, and bFGF (5, 6). However, when the array membranes were shortly incubated with secretion media of the cells, the pronounced hybridization signals were only observed for six cytokines; IL-6, IL-8, MCP-1, TIMP-2, VEGF and OPG and the rest majority of cytokines were hybridized at zero or minimal level.
It was found that this cytokine secretion profile was highly conserved among BM-derived MSCs from donors of diverse race, age and sex. The conserved secretion pattern, together with the results from our genome-wide gene expression (26) and proteomic profiling studies (Jeong et al., unpublished results), suggested that the molecular resource of the cells would not be greatly influenced by the individuality factors of the donors. Furthermore, the profile was found to be similar, although not identical, to that of UCB-derived MSCs, reinforcing, together with high commonalities in cellular characteristics, differentiation potential and genetic resources, the idea that the two cell populations are the same cellular identity.
In both cases with BM and UCB-derived MSCs, a spot for IL-6 on the array membrane produced the strongest hybridization signal, indicating that IL-6 might be a cytokine secreted predominantly from the MSC population. This pleiotrophic factor were already known to be expressed in a constitutive and excessive manner for diverse biological functions, such as proliferation of HSCs and stromal cells, growth and differentiation of B and T lymphocytes, regulation of inflammatory responses, and many other functions related to bone and BM (29). IL-8 was also known as an MSC-secreted cytokine with mitogenic and angiogenic potential (30), but its higher secretion level in UCB-derived MSCs compared to BM-derived cells was not previously reported. UCB-derived cells requires a higher amount of IL-8, probably because the target cells in the neonatal tissue are less mature with respect to the integrity of the cell signaling system. Aside from the two interleukins, the preferential secretion of the rest four cytokines over others in MSCs has not been well documented. It was only recent to show that MCP-1, TIMP-2, and VEGF, but not OPG, would be secreted from UCB-derived cells in appreciable quantities (31), although the functional roles of these cytokines with respect to MSC’s intrinsic functionalities are not as yet known.
In summary, the trophic nature of human BM-derived MSCs was assessed by an antibody array. This wide-spectrum cytokine secretion profile would not only provide a valuable insight on MSC’s roles in hematopoietic support and immune modulation, but also facilitate future studies on molecular mechanisms underlying their core biological properties.
Go to :
 XML Download
XML Download