

 PDF
PDF Citation
Citation Print
Print


Introduction
T cells are a type of white blood cell that plays a central role in cell-mediated immunity. The majority of T cells are produced after colonization by bone marrow (BM) progenitors in the thymus, where the thymic microenvironment directs differentiation, as well as positive and negative selection. During these differentiation steps, the earliest progenitors that are clearly T-lineage committed are CD34lowCD3−CD4+CD8− intrathymic T progenitor (ITTP) cells (1, 2). Notch signaling was regarded as essential during T-lineage development from multi-potent hematopoietic progenitors (3).
Compared to peripheral blood transplantation, human umbilical cord blood (UCB) transplantation is a more promising route in order to treat pediatric and adult patients because of its rapid availability, lower risk of infectious disease transmission, and lower risk of graft-versus-host disease (4). UCB transplantation has been well studied. CD34+ cells, regarded as the population containing hematopoietic progenitor cells, were the valuable fraction when used for transplantation. However, transplantation purified CD34+ cells have been reported to have reduced diversity of the peripheral T-cell repertoire (5).
Previously, in order to overcome delayed engraftment and impaired immune reconstitution, several researches have been tried, intensively. However, the major limitations were remained (6). Verhasselt et al. (7) has applied several conditions for expanding lymphocyte progenitors that could generate T cells and nature killer cells in fetal thymus organ culture. However, applying fetal thymus organ might bring extrinsic contamination and face the obstacle of resource limitation. In this study, a CD3−CD4+ CD8− intrathymic T-cell progenitor (ITTP) enriched population has been directly harvested from a CD34+ population, derived from UCB in a feeder layer-free system. This population has shown their progenitor character by the colonigenic potential. Thus, we developed a method for expanding large scale of T lineage cells from cord blood, efficiently, which might contribute to both basic research and clinical usage.
Go to : 
Materials and Methods
CD34 positive cells isolation from human umbilical cord blood by MACs
Human UCB was obtained from the umbilical vein immediately after vaginal delivery with the informed consent of the mother, approved by Borame Hospital Institutional Review Broad (IRB). Also, this experiment was approved by Seoul National University IRB.
UCB-mononuclear cells (MNCs) were prepared from heparinized UCB from a single parturient by Ficoll-Hypaque density centrifugation, according to standard procedures (8). CD34+ cells were isolated from the UCB-MNC fraction, using the MACS magnetobead separation system (Miltenyi Biotec). CD34+ enriched cell were sorted for deletion of CD4+ cells by FACs.
Ex vivo expansion and long term culturing
CD4−CD34+ cells (5×104/ml) were culture in 24-well plates with culturing medium that is composed of Iscove’s modified Dulbecco’s medium (IMDM), supplemented with a cytokine-limited cocktail, consisting of 20 g/ml Thrombopoietin (TPO), 50 ng/ml Stem Cell Factor (SCF), and 50 ng/ml Flk-2/Flt3 ligand (FL) (all recombinant human cytokines were purchased from PeproTech EC Ltd, London, UK), and 1%BSA (abbreviate as STF medium) in 5% CO2. Every 7 days of culture, a proper amount of cells were used for analyses. The remaining cells were sub-cultured in STF medium by half medium, changed weekly after the cells were centrifuged down at 1,500 rpm, 10 min.
Flow cytometry analysis of surface antigen expression
Right after MACs separation and every 7 days during culture term, cells were analyzed by flow cytometry on a FACSCalibur analyzer with CellQuest software (Becton-Dickinson). Cells were stained with fluorescein isothiocyanate (FITC)-conjugated anti-human CD34, CD45, CD38 CD4, CD51/61, CD2, or CD20, and phycoerythrin (PE)-conjugated anti-human CD10 or CD117, CD135, CD29, CD31, CD44, and CD90 (all antibodies were purchased from Becton-Dickinson). A replicate sample stained by FITC or PE conjugated anti-human IgG was used as an unstained control to ensure specificity.
Colony formation unit assay
Long term (8 weeks) cultured human UCB derived cells were sorted by FACS after stained by the CD4 antibody (Becton-Dickinson). Sorted CD4+ cells were then plated in duplicate plates in methylcellulose, supplemented with 100 ng/ml granulocyte-colony-stimulating factor (G-CSF), 1 ng/ml granulocyte macrophage-colony-stimulating factor (GM-CSF), 5 ng/ml recombinant human IL-3 (rhIL-3), and 20 ng/ml recombinants SCF. Colony-Forming Units (CFU-GM) with more than 100 cells were counted after 10 to 12 days.
Western blotting
Whole cell lysates extracted from freshly isolated hematopoietic progenitor cell and ITTPs cultured in STF medium for 30 days were subjected to western blotting.
Blots were incubated with primary antibodies: Mouse anti-human pPTEN (Cell Signaling Technology), rabbit anti human PTEN (Cell Signaling Technology), rabbit anti-human Notch-1 (Chemicon International, Inc., Temecula, CA), rabbit anti-human Akt (Cell Signaling Technology), rabbit anti-human pAkt (Cell Signaling Technology), rabbit anti-human JNK (Santa Cruz Biotechnology), rabbit anti-human c-Myc (Santa Cruz Biotechnology) and beta-actin (Sigma) as control. and the primary antibodies detected using horseradish peroxidase labeled rabbit anti-mouse immunoglobulin G or goat anti-rabbit immunoglobulin (both from Zymed) followed by enhanced chemiluminescence (Perkin Elmer Life Science).
Statistical analyses
The difference between groups was analyzed using student’s t test. P values <0.05 were considered as significant difference.
Go to :
Results
Expansion of CD3−CD4+CD8− ITTPs-enriched population
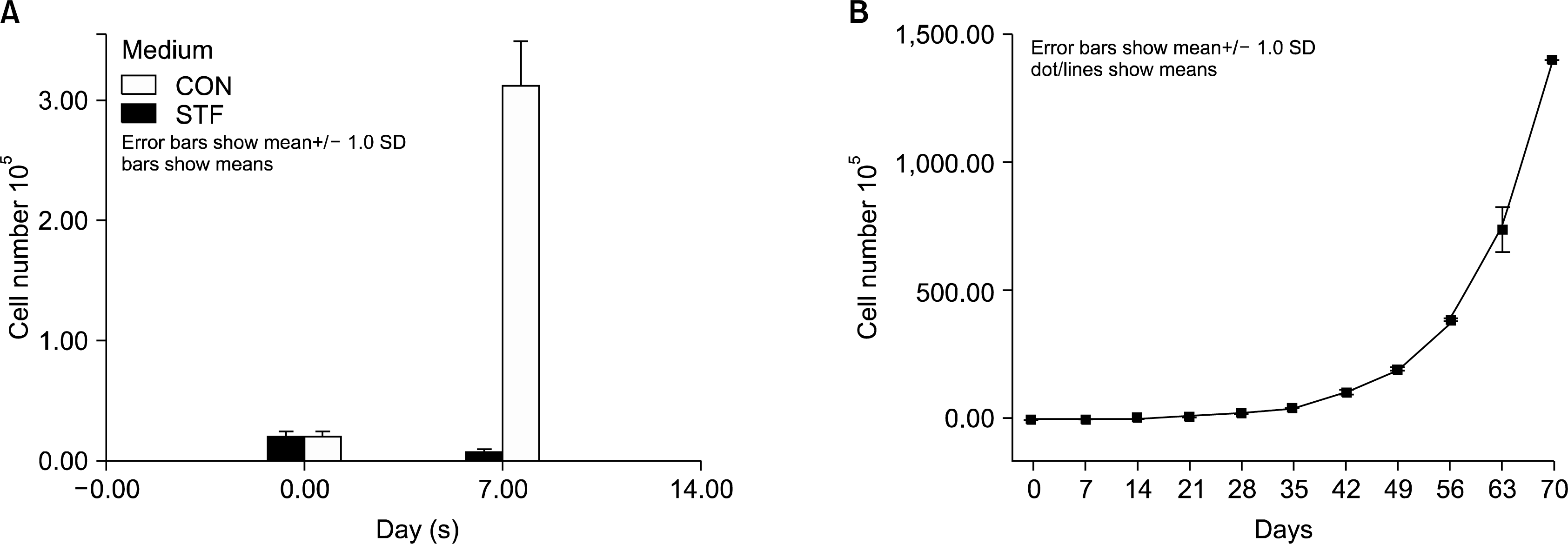
The highly purified CD34+ cells fraction was cultured in the STF medium in a 25 flask. In the first week, the cell number was dramatically increased nearly by 15 times (Fig. 1A). At the second week, cells were transferred to a 75 cm flask with medium, and another 5ml of new medium were added. From the third week on, half of the medium was changed weekly. Cells were expended continuously by adding new medium and the cells were maintained at a concentration less than 1×106/ml. Cells proliferated continuously at least for 10 weeks (Fig. 1B). At the end of the 10th week, cells were able to expand nearly 2.5×104 fold, compared to the seed concentration as 0.6×105/ml. Those results have shown, while the cells shift from CD34+ population to CD3−CD4+CD8− ITTPs-enriched population, they could expand dramatically to satisfy the requirement of transplantation for adult.
FACS analysis after conversion CD4+ cell population from CD34+ population
After CD4+ cells deletion by FACs, selected CD34+ cells had a purity of 90∼98% and the CD4+ population was totally deleted, as determined by fluorescence-activated cell sorting (FACS) (Fig. 2B, C). However, after culturing in STF medium for more than five weeks, CD4− CD34+ fraction decreased to 1%. However, the fraction expressing CD4 went up to 86.5% (Table 1).
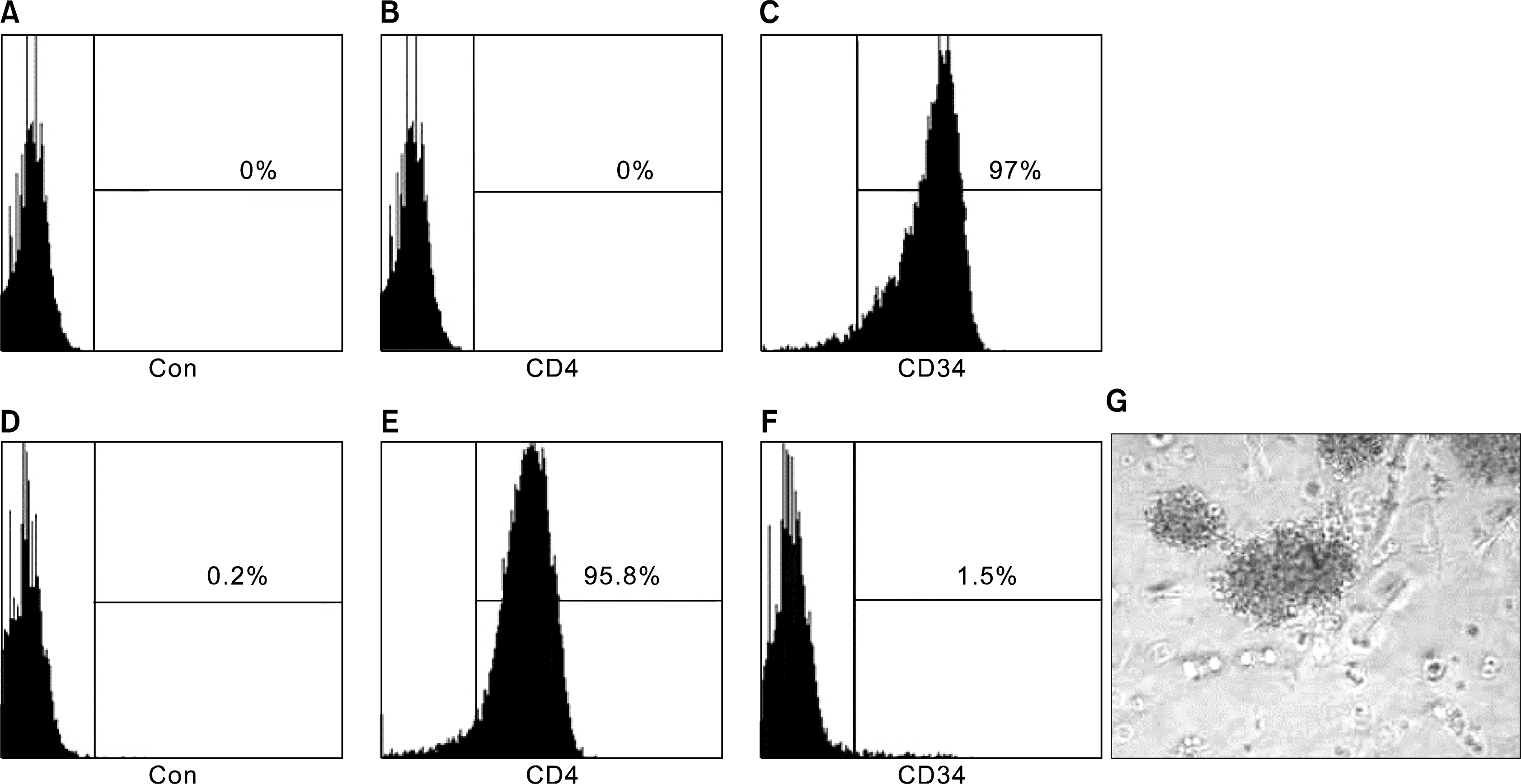 | Fig. 2.The change of CD4 and CD34 expression and the Colonigenic capability of long term cultured CD3−CD4+CD8− cells. Panel (A–C) were the result for the CD34 enriched cells right after of MACs, while (D–F) show the result of the same population cells after 5 weeks culturing. (A, D) was the fluorescence level in control, (B) and (E) shown the CD4 expression and (D) and (F) shown the CD34 expression. (G) The colony forming unit derived for long term cultured CD3−CD4+CD8− cells. Colonies were colorless and compact.
|
Table 1.
The expression of each selected CD antigens in the CD3− CD4+CD8− cells enriched population (n=3)
![]()
Cells, after long term culturing, were characterized by their surface CD series antigen markers. the original CD34 positive cells, undergoing long term culture in STF medium, had a large fraction which expressed CD29 (94%), CD4 (86.5%), CD44 (94.9%) and CD45 (95.6%). The fraction expressing CD34 was lost (Table 1) whereas ITTPs were dominant in the long term cultured cell population.
CFU assay of the long term cultured CD3−CD4+CD8− Intra-thymic T cell progenitor-enriched population
In order to confirm if there were some T cell progenitor cells in the CD3−CD4+CD8− cells population, after long term culturing in STF medium, we preformed CFU assay at the 7 and 8 week after sorting the CD3−CD8− population with the CD4 antibody. After ITTPs were cultured in methylcellulose-based CFU medium, about 40 colonies/2×104 cells developed. The developed colonies were colorless and compact (Fig 2G). Results gave the evidence that the CD3−CD4+CD8− cells-enriched population contained progenitor cells.
Signaling pathways involving in the expansion and differentiation
In order to examine the specific signaling pathways involving in proliferation of ITTPs, first of all, we examined the expression of pPTEN. The level of expression was slightly down regulated after the cells cultured in STF medium. An increase activation of Akt was not observed. However, PI3K, JNK and c-Myc were dramatically up regulated (Fig. 3A). In order to know whether the CD34+ hematopoietic progenitor cells convert to T-cell lineages, we examined the expression of Notch-1 in the freshly isolated CD34+ cells and the long term cultured CD4 positive cells. Because the essential role of Notch-1 signaling in intrathymic T lineage commitment and subsequent T cell development has been clearly documented (9). Notch-1 was highly expressed in the freshly isolated CD34+ cells and then down regulated in CD4+ cells after culturing and expanding in STF medium (Fig. 3A).
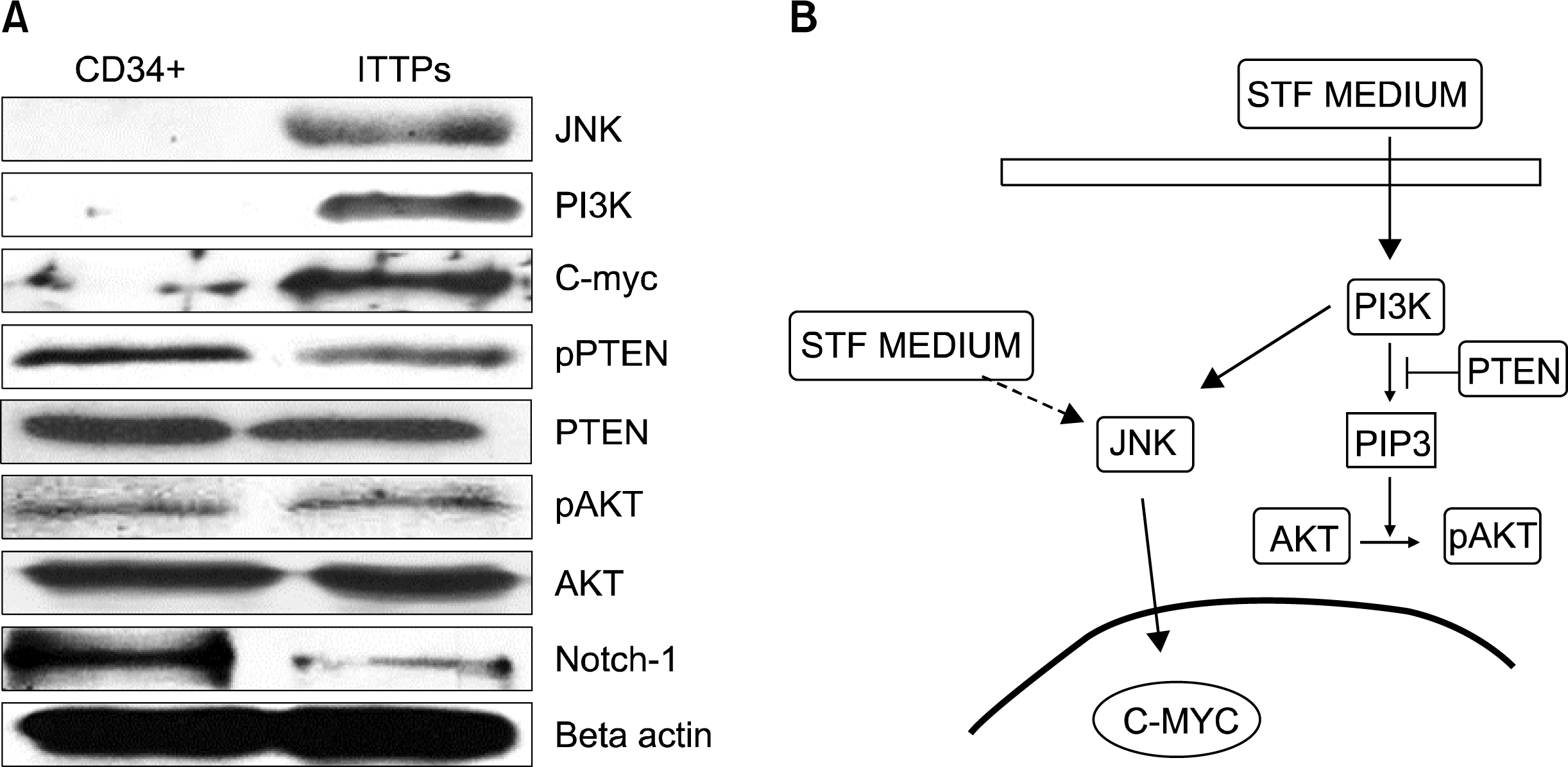 | Fig. 3.(A) The expression of related proteins in the freshly isolated CD34+ population and ITTPs cultured for 30 days. (B) Schematic diagram for a possible pathway which might regulate the proliferation of ITTPs. After culturing in STF medium, the expression of PI3K was upregulated. However, Akt could not be active due to PTEN suppress the production of PIP3. The upregulation of JNK, c-Myc indicate a PI3K/Akt-independent pathway (indicate by the bold arrow). Other factor in STF medium also might contribute to the activation of JNK (indicate by the broken line).
|
Go to :
Discussion
Transplantation with highly purified CD34+ cells results in reduced diversity of the peripheral T-cell repertoire during the early post-transplant period compared to patients receiving un-manipulated or MoAb-depleted transplants (5). In our study, the STF medium supported the development and long term expanding of T cell progenitors, which clinically could be of more fundamental help to relieve this dilemma.
We obtained the population which could generate sufficient amount of CD3−CD4+CD8− ITTPs simply by CD34+ cells selection, and what’s more, in our STF medium, those purified CD34+ cells could differentiate into T cells lineages, despite, physically, T cell development occurs in the thymus, where the Notch pathway could play the critical role (10). In the present study, we showed the high expression of Notch-1 in the CD34+ population in human cord blood. Obviously, in the STF medium, the balance leans to drive the hematopoietic stem cells to differentiate to T cell lineage. This might help to explain that in our study, the ITTPs cells population were expanded, while CD20+ cells, which is indicative of the B cells, are still rare (11). Unlike the thymus, which is incapable of supporting the long-term self-renewal of progenitors (12), the ITTPs could expand continually at least 10 weeks without the sign of exhausting in the present study.
The colony forming ability and long term culture-supporting ability indicate evidence of progenitor cells. Together with the evidence provide by Verhasselt Bruno et al. that in the similar condition, CD34+ cells could expand and provide lymphocyte progenitors for several weeks (7).
The up-regulation of PI3K, JNK and c-Myc indicate an Akt-independent pathway which might correspond for the proliferation of ITTPs. The expression of PI3K induced by the binding of SCF and KIT were reported (13). In stead of the Akt activation by PIP3, there is a PI3K/Akt-independent survival signal (14) that could response for the survival and proliferation of ITTPs. However, other growth factor like FL, TPO or the unknown factors contained in serum may also activated JNK pathway (15–19) (Fig. 3B).
T cell progenitor cells can generate both CD4+ and CD8+ T cells. Consideration of this, how to control the progenitor cells differentiation into desired direction needs to be studied further. In this study, the STF medium might direct the hematopoietic progenitor cells to differentiate into T cells progenitors. Though this needs to be fully investigated in further studies, our results indicated that the CD34+ enriched cells could switch to a CD3−CD4+CD8− ITTP-enriched population simply in STF medium but do not need culturing on fetal thymus or by adding other supplement. This will help the studies related to the development of T cell, as well as to contribute to improvement of the difficulties in clinical transplantation and therapy for various diseases, where the T cells play a central role.
Go to :
 XML Download
XML Download