

 PDF
PDF Citation
Citation Print
Print


Introduction
The hair follicle contains dermal (dermal papilla and connective tissue sheath) and epidermal (matrix and inner and outer root sheaths) components. Its structure consists of a hair shaft surrounded by several concentric layers of epithelial cells that form the inner and outer root sheath, a mesenchymal sheath surrounding this epithelial core, and a sebaceous gland, which is an outgrowth at the side of the hair germ. Hair follicles are self-renewing structures that cycle between growth (anagen), regression (catagen), and resting (telogen) phases and reconstitute themselves throughout life because they contain stem cells (1). Chase postulated that upper outer root sheath contains a population of multipotential stem cells which is capable of forming the follicle, the epidermis and the sebaceous gland. More recent work based on cell culture studies (2) and a wide range of in vivo studies in the mouse (3, 4) have suggested a somewhat more complex organization and distribution of stem cells in skin, with stem-like cells implicated at specific locations in the interfollicular epidermis, in the upper regions of the outer root sheath of the hair follicle (the so-called bulge region), and in the germinal matrix of growing hair follicles. The interrelationship between these three separate stem cell compartments remains obscure, although it can be hypothesized that the bulge region stem cells represent the most potent reserve population of ultimate stem cells (5).
During the past decade, several molecules have been suggested as putative markers of hair follicle stem cells. The candidate molecules are α6-integrin (6), β1 integrin (7), cytokeratin 15 (8), cytokeratin 19 (9), p63 (10), S100A (11), Caveolin-1 (12), CD200 (13), PHLDA1 (pleckstrin homology-likedomain, family A, member-1) (13), WIF1 (WNT inhibitory factor 1) (13), DKK3 (Dickkopf homolog 3) (13), FSH (follistatin) (13), FZD1 (frizzled homolog 1) (13), nestin (14), CD71 (15) and CD34 (16, 17). However, a major obstacle in this field have been lacking of reliable molecular markers for hair follicle stem cell so far.
CD34 is a 105∼120 kDa glycoprotein expressed on early hematopoietic progenitor cells and is believed to behave as a stage-specific antigen as its expression is progressively lost during cell maturation. (18). Moreover, CD34 expression is also found in several nonhematopoietic tissues, including skeletal muscle (19), gastrointestinal tract (20), and endothelial cells (21). It was recently proposed that CD34 is specifically expressed on stem cells isolated from the bulge region of hair follicles in animal or mice. Trempus et al (16) identified CD34 staining on keratinocytes in the bulge region of mouse hair follicles, CD34 expression co-localized with label-retaining cells, cells were quiescent when isolated from the bulge region, and cells showed a high proliferative potential in vitro. This implies that the expression of CD34 in the bulge region suggests potential relations between haematopoietic stem cells and epidermal stem cells. However, epidermal cells that are CD34−negative also show stem cell properties (22, 23) and epidermal stem cells sorted by Hoechst 33342 dye exclusion method from newborn mice did not express CD34 either (24). Moreover, Ohyama et al reported CD34, which delineates hair follicle stem cells in the mouse, is not expressed by human hair follicle stem cells, while CD200 is expressed by stem cells in both species. Much more work is needed to clarify these conflicting results about CD34.
In this report, given these roles of CD34 in hair follicle, to determine whether human hair follicle CD34+ cells can regenerate new hair follicle in vivo, we isolated CD34+ cells from cultured human hair follicle cells and injected the CD34+ cells to immunodeficient nude mouse. And then, we analyzed skin biopsies from the nude mice.
Go to : 
Materials and Methods
Human occipital skin biopsies
Normal human scalp specimens were taken from volunteers without systemic disease, who were undergoing plastic surgery and all subjects gave written informed consent. Biopsies were uniformly taken from the occipital scalp region of men who did not suffer from male-pattern alopecia or from any other diseases affecting the growth of hair and scalp. All tissue donors (ten patients) were male with average of 33 years. (range 26∼41 years). The tissue sample was transported to the laboratory in the media consisting of Dulbecco’s Modified Eagle Medium (DMEM) (Gibco Life Technologies, Gaithersburg, MD, USA) containing antimycotic agents to yield final concentrations of 15 units /ml penicillin, 15 mg/ml streptomycin, 30 ug/ml neomycin (Sigma, St Louis, MO, USA) and 100 ug/ml Normosin™ (Invivogen, San diego, CA, USA).
Cell culture
The sample tissue was rinsed in Hanks Bss to remove blood and debris and was placed in a 100-mm plastic tissue culture dish (Nunc, Rochester, NY, USA). The entire procedure was performed under a sterile laminar flow hood. Fat and connective tissue was cleared from the sample tissue. The rest of the tissue was minced with sterile scalpels into 1∼2 mm fragments and digested with Accumax™ (2%; Chemicon international Inc., CA, USA) and Dispase (0.4 mg/ml; Sigma.) in DMEM medium (Gibco Life Technologies) which consisted with 10% fetal bovine serum (Gibco Life Technologies) plus antibiotic and antimycotic agents (as above) for 30min at 37°C on a shaking incubator. And then the tissue washed with sterile PBS was digested with collagenase type IA (2 mg/ml; Sigma) in DMEM medium (Gibco Life Technologies) which consisted with 10% fetal bovine serum (Gibco Life Technologies) plus antibiotic and antimycotic agents (as above) for 30 min at 37°C on a shaking incubator. The tissue digest was vigorously pipetted. After gentle centrifugation (2,500 g, 5 min), the supernatant was removed, and the pellet was resuspended in Medium 199/Ham’s F12 (M199/F12; 1:1) medium which suplemented with 10 ng/ml human EGF, 20 ng/ml FGF, 5 mg/ml insulin, 0.5 mg/ml hydrocortisone, 2 mg/ml human transferin, 125 ug/ml bovine serum albumin, 10% fetal bovine serum (FBS) and antibiotic and antimycotic agents (as above) and distributed on a 25 cm2 flask (Nunc). After 3 days, Culture medium of the hair follicle cells attached on the flask was changed to serum free medium (without FBS) and suspended cells on the flask were transfer to 75 cm2 flask (Nunc) with complete culture medium (as above). As a same manner, the complete culture medium was changed to serum free medium (without FBS) after 3days and placed for 14 days at 37°C in a humidified atmosphere containing 5% CO2. Each culture medium was carefully removed every 3 days and replaced by fresh culture medium. After 7 days in culture, all culture medium was changed to Defined keratinocytes serum free medium (Gibco Life Technologies). Atfter 2 weeks in primary culture, cells were collected by incubation with 0.5 mg/mL trypsin, 0.2 mg/mL EDTA (ethylenediamine tetraacetic acid) solution (Gibco Life Technologies), for 5 min at 37°C. After this incubation period, clusters of cells were released from the hair follicles. These were harvested by centrifugation at 300 g at 4°C for 10 min.
Isolation of CD34+ and CD34− cells
To remove cell clump before isolation, The cells were vigorously pipetted and added to a stacked sterile mesh sieve assembly with 50 Pre-Separation Filters (30 um size; Miltenyi Biotech, Bergish-Gladbach, Germany). After the human hair follicle cells collected were added to the top of the sieve assembly, cell aggragates or large particles from the samples were retained in mesh sieve while single cells passed through to the receptacle below. The single human hair follicle cells were resuspended cold phosphate buffered saline containing 0.5% bovine serum albumin (BSA) and 5 mm EDTA (MACS buffer), pH 7.4. and uniform, magnetic beads (Miltenyi Biotech), which were coated with a monoclonal antibody specific for human CD34, were used to separate CD34+ cells. The human hair follicle cells were resuspended to a concentration of 1×108 cells in 300 ul PBS, 5 mM EDTA. These cells were incubated with 300 ul MACS buffer, 100 ul beads and blocking reagent (human IgG to prevent non-specific binding) per 108 cells for 30 minutes at 4°C. After washing with MACS buffer, cells were loaded onto a MiniMACS column (Miltenyi Biotec Ltd, Bisley, Germany) held within a high-density magnetic field to retain CD34+ cells. Unbound cells (CD34− cells) were eluted with MACS buffer and retained CD34+ cells were eluted by washing with buffer after the column had been removed from the magnet. Cells in the MACS positive fraction were applied to a second column and the purification step repeated. Human hair follicle cells and cells from the MACS positive and negative fractions were assessed by cell counting and flow cytometry. Selected CD34+ cells were detached from the magnetic beads using the plunger. CD34+ and CD34− cells were resuspended in Iscove’s modified Dulbecco’s medium (IMDM) (Gibco Life Technologies).
Flow cytometry studies
To confirm the CD34+ and CD34− populations by FACS analysis, Each 2×104 cells of the CD34+ and CD34− cells isolated as described above seeded in each well of 96 well plate. Cells were incubated for 40 min at room temperature with 1:10 final fluorescein isothiocyanate (FITC)-, phycoerythrin (PE)-conjugated mAb in PBS, 2% FCS. After washing, Cells were fixed with 1% paraformaldehyde in PBS before analysis on a flow-cytometry (FACSCalibur, BD Immunocytometry Systems, CA, USA). The mAbs were: CD34-FITC, CD45-FITC, CD24-PE, CD44-PE, CD133-PE, CD29-PE (BD Phar-Mingen, CA, USA). Controls did not include any mAb.
Hair-growth studies
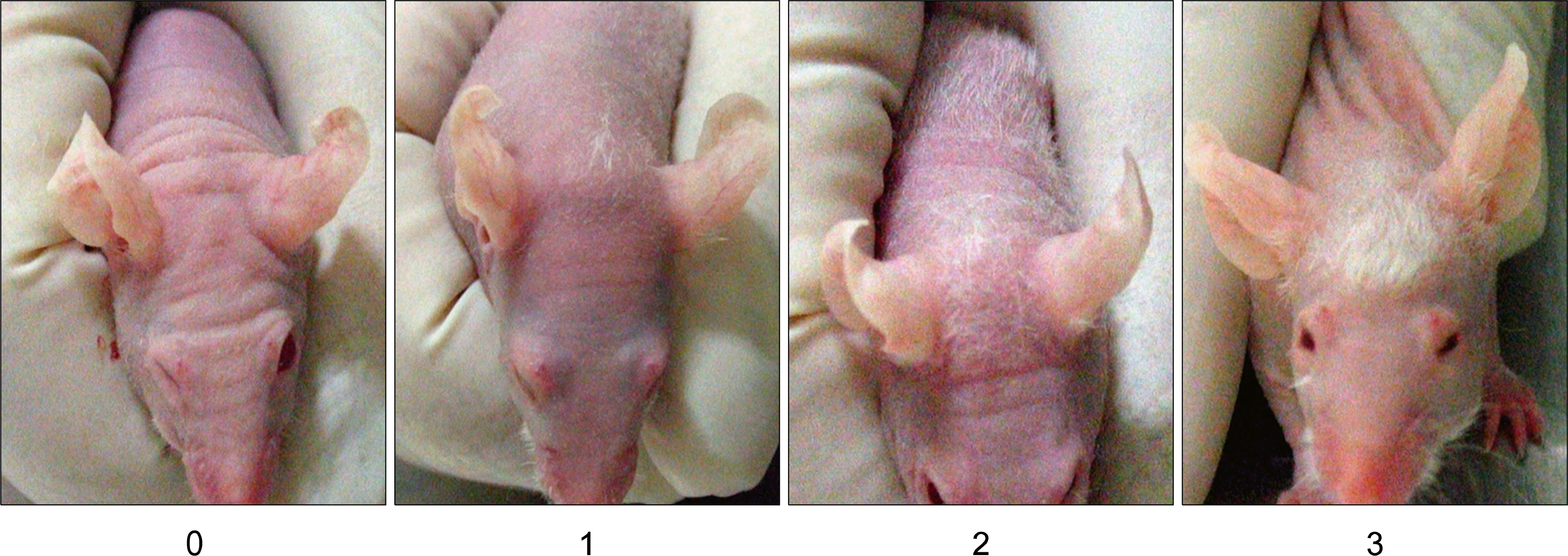
A total of 20 female BALB/c AnNCrjbji-nuBR mice (mean body weight 25.2±2.0 g) were obtained at 6 wks of age from Charles River (Boston, MA, USA) and used in accordance with the laboratory animal guidelines of Seoul National University. Mice were kept in protective cages (five mice in each) with unrestricted access to water and diet at standard laboratory conditions and were kept under 12h light/dark cycles. The animals were divided into four experimental groups as follows: group I (n=5), no injection; group II (n=5), Saline injection alone, as a control; group III (n=5), intradermal transplantation of isolated CD34− hair follicle cells; group IV (n=5), intradermal transplantation of isolated CD34+ hair follicle cells. Before transplantation, the hair of each group mice was removed by the topical application of calcium thioglycolate. The next day, approximately 1×105 isolated CD34+ and CD34− hair follicle cells suspended in 50 uL of physiologic saline were injected into the parietal area of each group mice head using a 30-gauge Venocath (Becton Dickinson Medical systems, Sandy, UT) and 50 uL of physiologic saline was injected into same area of group I, II mice. In the in vivo studies, hair-regrowth activities when injecting cells was measured by observing hair of each mouse every day. All experiments lasted for 30 days. Fig. 3 shows the scale for evaluating hair growth in this mouse.
Western blot analysis
After 30 days from injection of the cells, biopsies were obtained from the parietal area of the mouse head. Samples of parietal skin from all group homogenized in lysis buffer (150 mM NaCl, 50 mM Tris-HCl, pH 7.4, 2 mM ethylenediamine tetraacetic acid (EDTA), 1% NP-40) containing protease inhibitors (Complete, Roche, Mannheim, Germany) on ice. Homogenate was centrifuged at 12,000 g for 1 h at 4°C. The supernatant was collected and subjected to determination of protein concentration using commercial protein assay kit (Bio-Rad, Richmond, CA, USA). Equal amounts of protein samples (25μg/lane) were subjected to 12% SDS-PAGE and transferred to nitrocellulose membranes. The membranes were subsequently incubated with the corresponding primary antibodies, as indicated: mouse anti-Bcl-2 monoclonal antibody (Zymed Laboratories Inc., CA, USA); rabbit anti-MAPK polyclonal antibodies (Zymed Laboratories Inc); rabbit anti-p38 monoclonal antibodies, anti-Bax polyclonal antibody (Santa Cruz Biotechnology, CA, USA); rabbit anti-active MAPK, anti-active p38 MAPK polyclonal antibodies (Promega Corporation, WI, USA); rabbit anti-Akt polyclonal antibodies (Cell Signaling Technology Inc. Beverly, MA, USA); (Sigma). Antibody recognition was detected with the respective secondary antibody, ether anti-mouse IgG, or anti-rabbit IgG antibodies linked to horse-radish peroxidase (Zymed Laboratories Inc.). Antibody-bound proteins were detected by the ECL western blotting analysis system (Amersham Pharmacia Biotech U.K. Limited).
Tissue processing
After 30 days from injection of the cells, biopsies were obtained from the parietal area of the mouse head after anesthesia with dimethyl ether. Samples of parietal skin from control group and experiment group were fixed in 10% (wt/vol) neutral buffered formalin for 24∼48 hours, dehydrated in an alcohol-xylene series, and embedded in paraffin wax. From each block, sections with 3μm thickness were prepared and stained with haematoxylin and eosin (HE) for histological examination and it was serially sectioned at 3μm, floated on a water bath containing diethylpyrocarbonate-treated water, and mounted on positively charged slides (Superfrost/Plus slide, Erie Scientific Co., Portsmouth, NH).
Primer & PCR amplifications
The specific primers for human Alu sequences were Alu-sense, 5’ ACG CCT GTA ATC CCA GCA CTT 3’, and Alu-antisense, 5’ TCG CCC AGG CTG GAG TGC A 3’, which produced a band of 224 bp. The primers were positioned in the most conserved area of the Alu sequence (25). The PCR amplifications were performed by using the following settings: 94°C for 30 s, 58°C for 30 s and 72°C for 45 s, with a total 30 cycles. The PCR products were analysed in 1% agarose.
Preparation of labeled probe
Polymerase chain reaction (PCR) products were purified using a 30-kd cutoff membrane ultrafiltration filter. The nucleotide sequences of the purified PCR products were determined by use of Big Dye chemistry with the ABI Prism Sequencer (Applied Biosystems, Foster City, CA, USA). Sequencing was performed of the purified PCR products before PCR products were labeled by random priming with digoxigenin-dUTP (Roche, Grenzacherstrasse, Switzerland), according to the manufacturer’s instructions.
In situ hybridization
Sections were deparaffinized in xylene and rehydrated in phosphate-buffered saline (PBS) (pH 7.4, 0.01 M) for 5 minutes. Deproteinization was carried out in 0.2 N HCl for 20 minutes at room temperature. Tissues were then digested at 37 C for 20 minutes in 100μg/ml proteinase K (GIBCO-BRL) in PBS. After digestion, tissues were fixed in 4% paraformaldehyde in PBS for 10 minutes. After rinsing twice with PBS, the slides were acetylated in 300 ml of 0.1 mM triethanolamine-HCl buffer (pH 8.0) to which 0.75 ml of acetic anhydride (0.25%) had been added. After 5 minutes, an additional 0.75 ml of acetic anhydride was added, and 5 minutes later the slides were rinsed in 2× saline sodium citrate (SSC) (1× SSC contains 50 mM NaCl and 15 mM sodium citrate, pH 7.0). The slides were allowed to equilibrate for 60 minutes in a standard hybridization buffer that consisted of 5× SSC with 50% deionized formamide, 10× 2% buffered blocking solution (Roche), 0.1% N-lauroylsarcosine, and 0.02% sodium dodecyl sulfate. Hybridization was done overnight at 45°C. The digoxigenin-labeled probe (0.1 ng/μl) was diluted in 300μl of the standard hybridization buffer, heated for 10 minutes at 95°C on a heating block, and quenched on ice before being applied to the tissue sections. Approximately, 50 ng of the digoxigenin-labeled probe was added to the standard hybridization buffer (50 μl), heated for 10 minutes at 95°C on a heating block, and quenched on ice before being layered over the section again. Fluid was held in place by a coverslip, and the edges were sealed with rubber cement. After overnight hybridization, sections were thoroughly washed twice in 4× SSC for 5 minutes at room temperature, twice in 2× SSC for 10 minutes at 37 C, twice in 0.2× SSC for 5 minutes at room temperature, and once in maleic acid buffer (100 mM maleic acid and 150 mM NaCl, pH 7.5) for 5 minutes at room temperature. For detection of hybridization, sections were incubated with anti-digoxigenin conjugated with alkaline phosphatase (Roche) diluted 1:250 in 0.1 M Tris-HCl (pH 7.4) and 0.15 M NaCl with 1% blocking reagent (Roche). After three washes in buffer, substrate consisting of nitroblue tetrazolium and 5-bromocresyl-3-indolylphosphate was layered over the sections. Color was allowed to develop for 5∼8 hours in the dark, and the reaction was stopped by dipping slides briefly in Triethylenediaminetetraacetic acid buffer (10 mM Tris-HCl and 1 mM ethylenediaminetetraacetic acid, pH 8.0). Sections were allowed to dry completely, dipped into the absolute xylene, and coverslipped with Canada balsam mounting medium (Hayashi Pure Chemical Industries Ltd., Osaka, Japan).
Statistical methods
All experiments were conducted at least in triplicate (n=3), and the results are expressed as the mean±SD. Statistical analysis was conducted via analysis of variance (ANOVA), followed by Duncan’s multiple range test.
Go to :
Results
Characterization of CD34+ cells isolated from human hair follicle cells
To confirm the CD34+ and CD34− population isolated by MACS, we performed FACS analysis of each population. CD34+ population was stained with antibodies to CD34 (FITC conjugate) and then subjected to FACS analysis gating out debris and dead cells, a distinct CD34+ population was found, which comprised 98% of isolated cells. Most of hair follicle CD34+ cells were CD133+ (approximately 99%), CD24+ (approximately 98%), CD29+ (approximately 99%), CD44+ (approximately 98%), but, a small population (approximately 1%) of human hair follicle CD34+ cells was also found to CD45+ (Fig. 1A). As might be expected, distinct CD34− population was found, which comprised very small percentage (approximately 1%) when were stained with antibodies to CD34 (FITC conjugate). Most of hair follicle CD34− cells were also negative for CD133 (approximately 17%). However, the intensity of expression of CD24 (approximately 96%), CD29 (approximately 99%), CD44 (approximately 92%) of CD34 negative population was also very high (approximately 96%) and the intensity of expression of CD45 was very low (approximately 2%), same as CD34+ population (Fig. 1B).
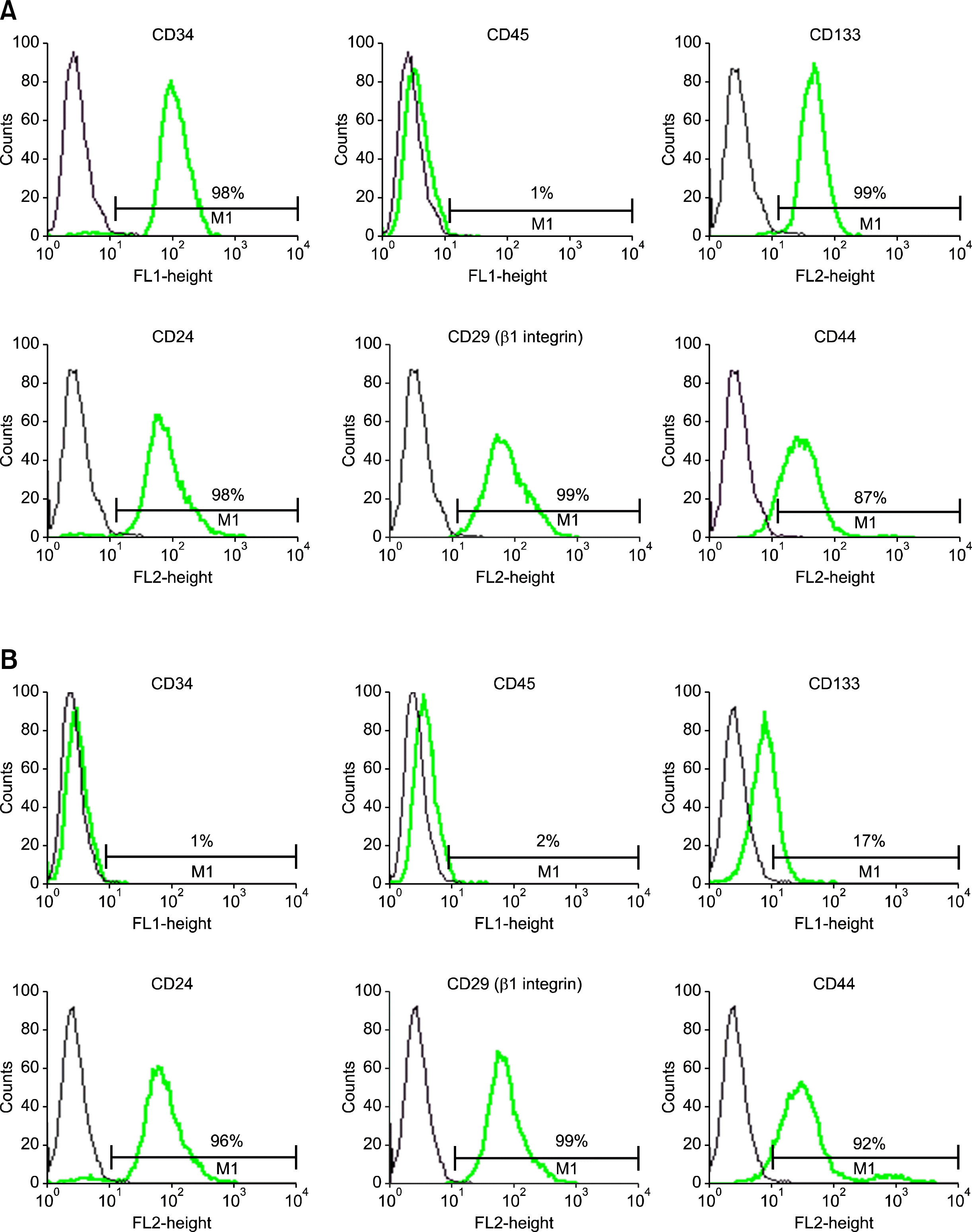 | Fig. 1.Flow cytometric histogram from CD34+ and CD34– cells isolated from cultured human hair follicle cells. Cells were stained with fluorescein isothiocyanate-conjugated antibodies against CD34, CD45 and phycoerythrin-conjugated antibodies against CD133, CD24, CD29, CD44 or immunoglobulin isotype control antibodies. Cells were analyzed using fluorescence-activated cell sorter Calibur. FSC/SSC gating to exclude debris. (A) The percentages of isolated CD34+ cells expressing FITC or PE are given. Staining on FACS analysis with CD34 was greater positive than 98%, and CD133, CD44, CD24, CD29 was also positive, whereas CD45was negative. (B) The percentages of isolated CD34− cells expressing FITC or PE are given. Staining on FACS analysis with CD34 was negative and CD45, CD133 was also negative, whereas CD24, CD29, CD44 was strongly positive, same as CD34+ population.
|
Human CD34+ hair follicle cells regenerate new hair in immunodeficient nude mice
New hairs were observed to head and dorsal area of mouse skin which had transplanted hair follicle CD34+ cells (Fig. 2B). But, CD34+ transplanted mice have more hairs and whiskers compared with CD34− transplanted mice (Data not shown). All mice of cell transplanted group (Experiment group) successfully produced new hair at 3 weeks after injection of the cells but physiologic saline-injected mice (Control group) fail to produce hair in the skin (Fig. 2A).
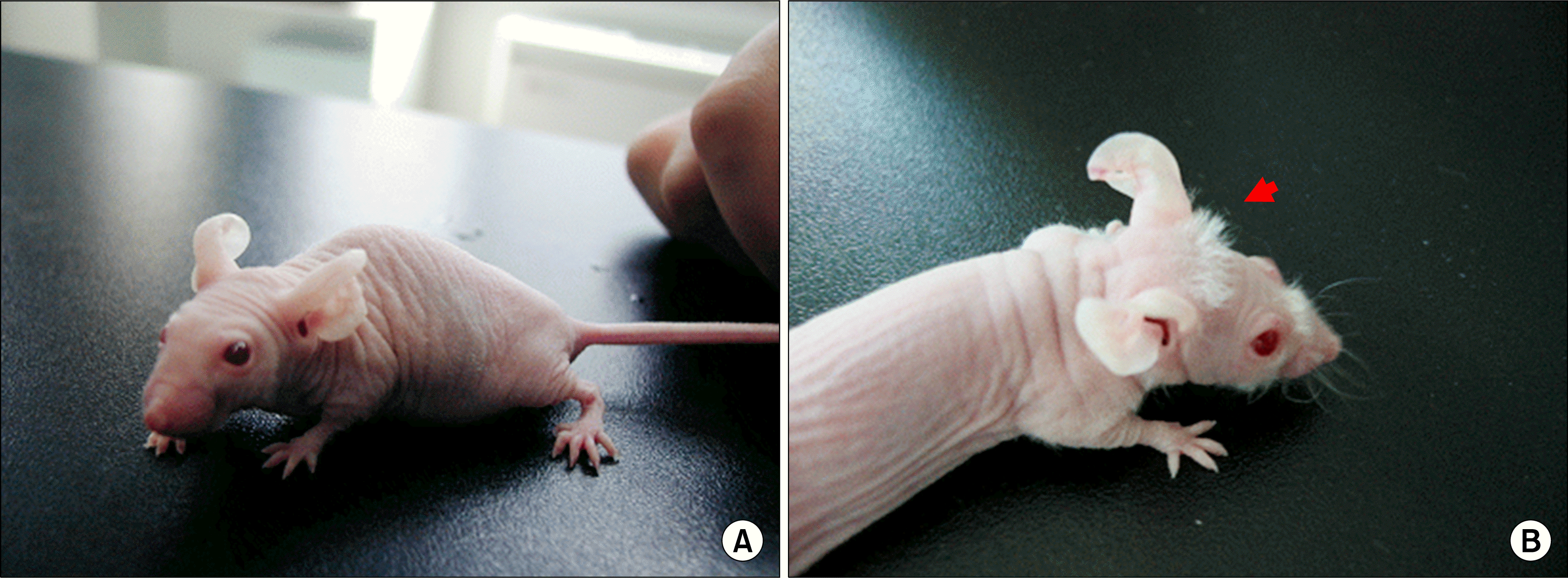 | Fig. 2.Hair growth from parietal region nude mouse head transplanted with human CD34+ hair follicle cells. As a control, physiologic saline-injected nude mouse fail to produce hair in the skin (A). Transplantation of nude mouse with human CD34+ hair follicle cells significantly resulted in new hair regeneration 3 weeks after injection of the cells (B, arrow).
|
Hair-growth studies
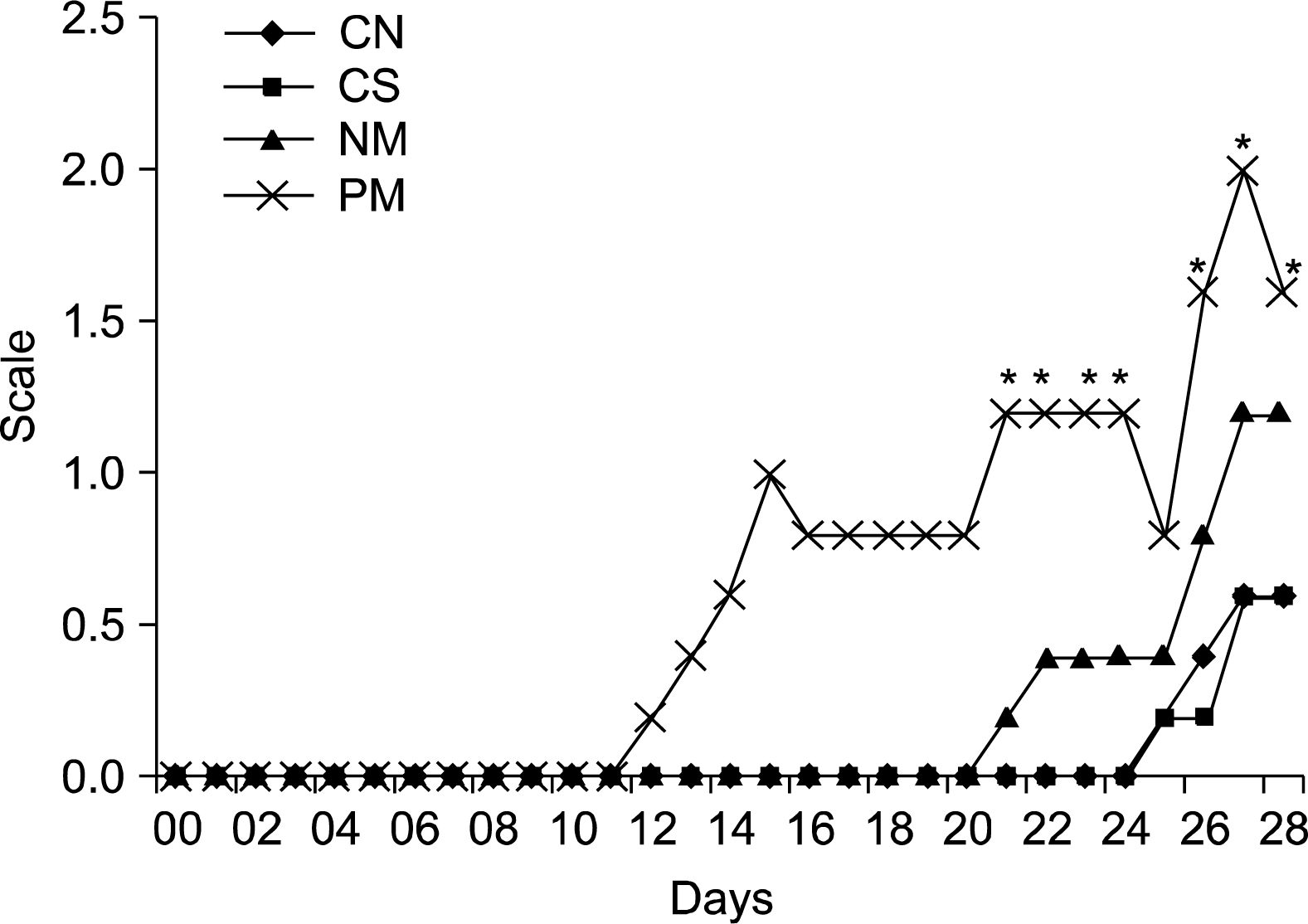
In comparison with the transplantation of vehicle only as the vehicle control and the no transplantation of as the negative control, CD34 positive and negative cells isolated from human hair follicle were transplanted to evaluate their efficacy on the promotion of hair growth using immunodeficient nude mice. Fig. 4 shows these results of quantified hair growth after transplantation using the scale illustrated in Fig. 3 and demonstrates hair growth for the negative control and vehicle control (Saline), respectively. In negative control mice, hair growth showed only a faint appearance 25 days after transplantation. After the transplantation of saline, it took about 25 days for the hair of mice to grow, indicating no effect of vehicle in the proliferative rate of hair growth. However, after transplantation of CD34+ and Cd34− hair follicle cells, it was evident that both were able to facilitate hair growth compare to the control.CD34+ transplanted mice have more hairs, whiskers and earlier compared with CD34− transplanted mice.
Histological examination of scalp sections
To examine Histological sections of skin in control group and experiment group, biopsies were obtained from the head parietal skin of each group 30 days after injection of human CD34+ hair follicle cells and haematoxylin and eosin (HE) staining was carried out. Very Few hair follicle was observed in the head parietal skin of a control mouse (Fig. 5A, B) but new hair follicles were observed in the head parietal skin of human CD34+ hair follicle cell-injected mouse (Fig. 5C, D).
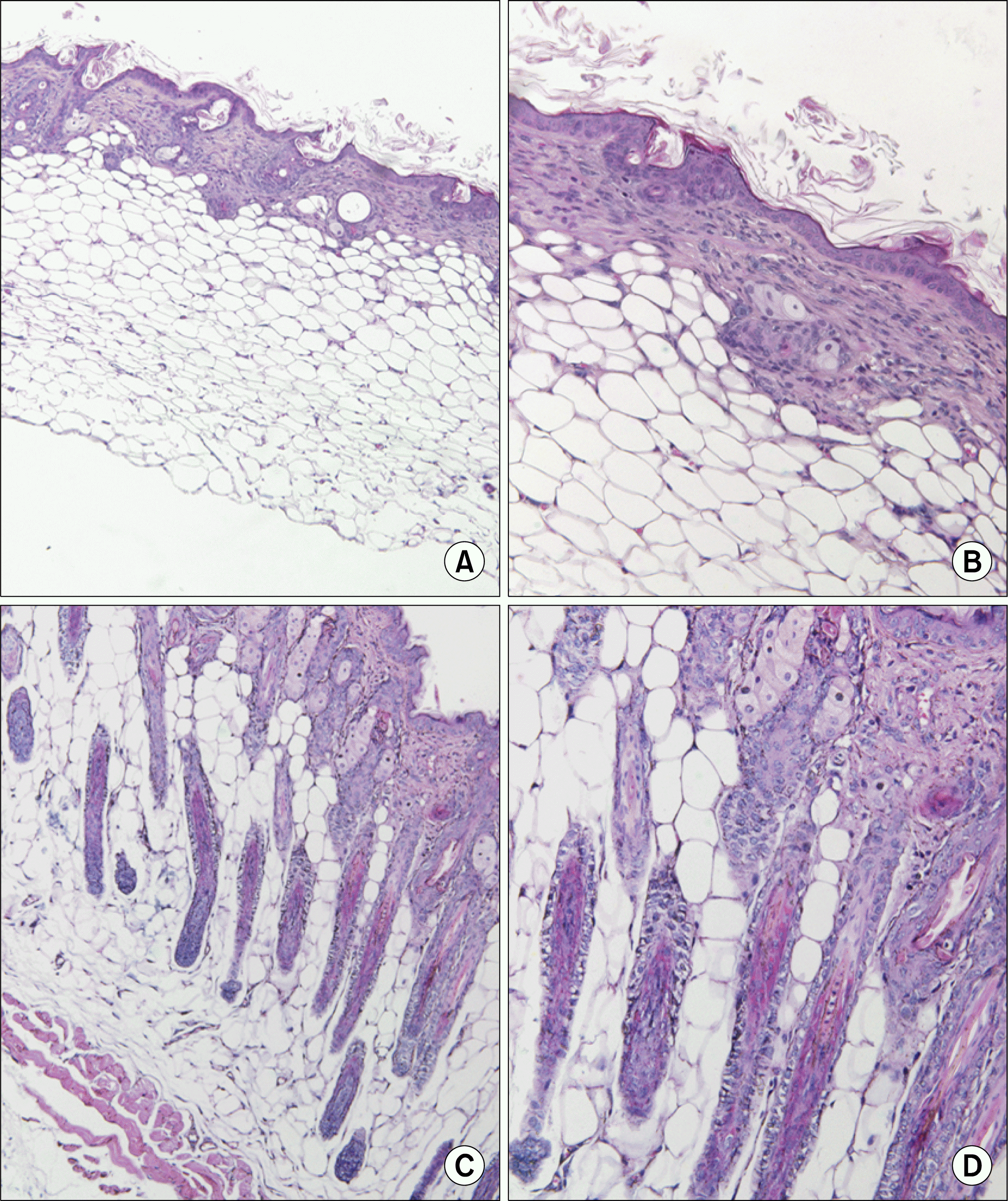 | Fig. 5.Histological appearance of human hair follicle CD34+ cell induced follicles within mouse head parietal skin. No hair follicle was observed in the parietal skin of a control mouse head (A, B) but new hair follicles were observed in the parietal skin of injected mouse (C, D) by using H&E staing.(Original magnification A, C; ×100, B, D; ×200).
|
Transplanted human CD34+ hair follicle cells were localized in outer root sheath (ORS) of new hair matrix in mouse
In situ hybridization of skin samples from each group with human cell specific probe, revealed the presence of transplanted human hair follicle CD34+ cells in the ORS keratinocytes of the nude mouse skin. The human specific DNA was identified around ORS of new hair matrix in CD34+ hair follicle-injected mouse (Fig 6B, C; arrows). This result indicated that human hair follicle cells transplanted were migrated and localized in ORS of new hair follicle: Control mouse was not detected (Fig 6A).
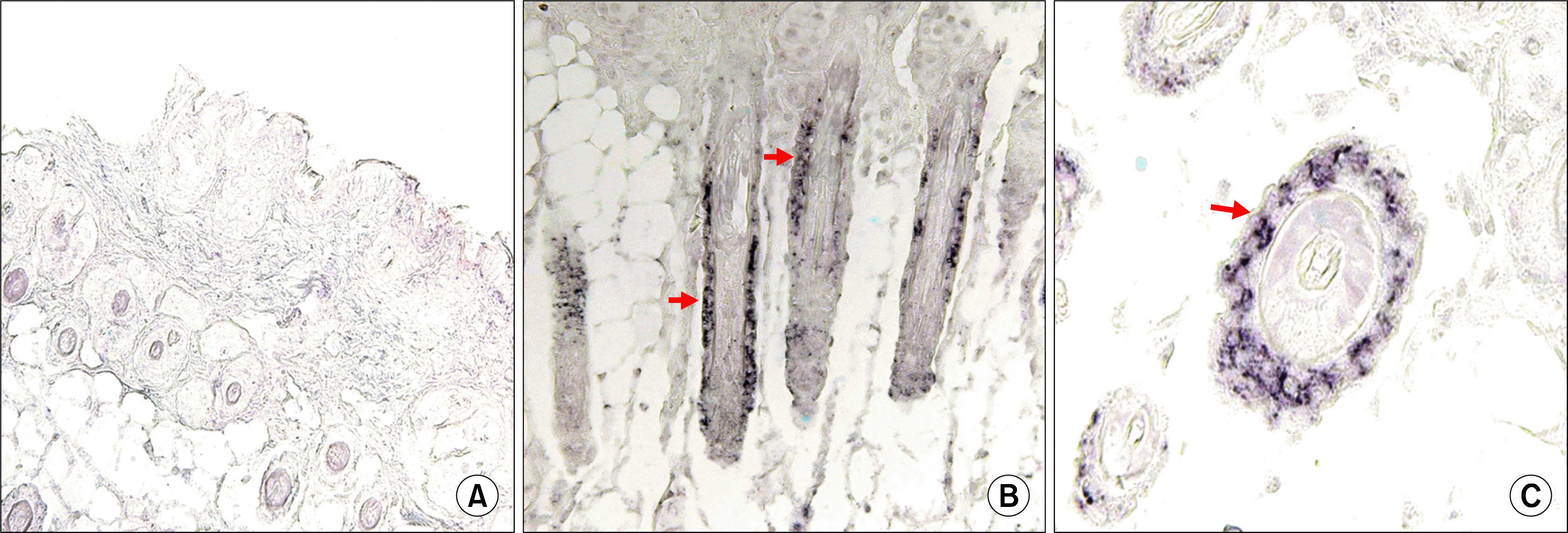 | Fig. 6.Human CD34+ hair follicle cells transplanted to nude mouse located in the ORS region of mouse hair follicle. Human specific DNA was not detected in control mouse (A), but identified in ORS of CD34+ human hair follicle cell-injected mouse hair follicle (B,C arrows) by in situ hybridization. Vertical section of mouse hair follicle (A, B); transverse section of CD34 human hair follicle cell-injected mouse hair follicle (C). (original magnification A, B; ×200, C; ×800).
|
Transplanted human CD34+ hair follicle cells induce phosphorylation of ERK up-regulation of Bcl-2 and down-regulation of Bax lgulat in nude mouse epidermal cells
Western blotting methods were used to investigate the expression pattern of several cell signaling proteins in control mouse and transplanted mouse. The results indicate that control mouse and transplanted mouse have the same expression level of total p38, pp38, ERK and AKT. But pERK expression levels were significant increased in transplantated mouse. The apoptosis-suppressing Bcl-2 was significant increased and the apoptosis-promoting Bax was significant decreased in CD34+ cell-transplantated mouse compared with control mouse (Fig. 7).
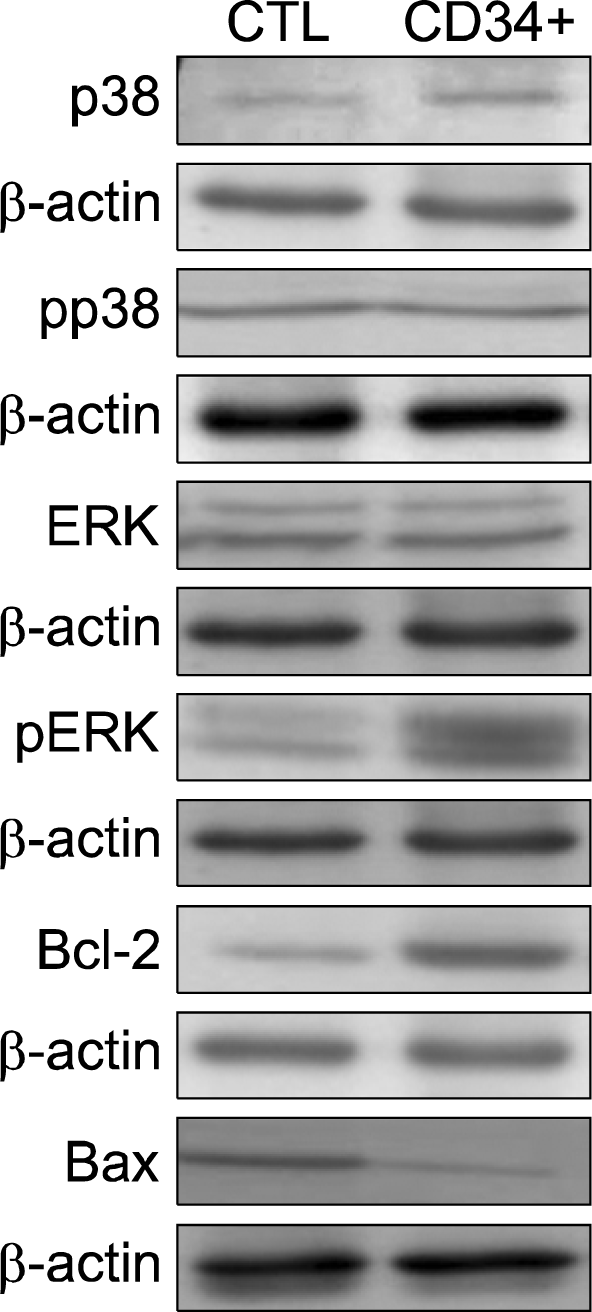 | Fig. 7.Change of protein expression for transplantation effect of human CD34+ hair follicle cells. Control mouse and transplantated mouse have the same expression level of total p38, pp38, ERK and AKT. But pERK expression levels were significant increased in transplantated mouse. Moreover, anti-apoptosis proteins, Bcl-2 was significant increased in transplantated mouse whereas proapoptosis proteun, Bax was significant decreased in transplantated mouse compared with control mouse.
|
Go to :
Discussion
In this study, we examined human CD34+ hair follicle cell and determined that the population specifically localized outer root sheath of recipient mouse hair follicle when transplanted to mouse. Besides, the human hair follicle CD34+ cells could regenerate more new hair than CD34− cells in the head and dorsal area of mouse.
CD34 known as hematopoietic stem and progenitor cell determinant localizes to the bulge region in mouse skin, and in combination with α6 integrin identifies a minor population of slowly cycling keratinocytes (26). In contrast, it reported that CD34, which is an excellent bulge cell marker in the mouse (16), is not on human bulge cells. Rather, CD34 is expressed in slightly more differentiated outer root sheath cells below the bulge in the human follicle (13). Even if CD34 is not specific marker for human bulge cells any more, the role of CD34 is very important for hair matrix or hair follicle stem cells as our results show that CD34+ cells were able to regenerate more hair follicle than CD34− cells. Therefore, these results suggest that not all human hair follicle stem cell compartments may exist in the bulge region. This is also supported by observation that cells with high clonal capacity are localized to a broad morphologically indistinct region of the proximal ORS (27). This suggests that in human hair follicles, stem cells do indeed occupy a larger niche than that occupied by murine stem cells, in terminal scalp hair follicles C8/144B expression is also extensive in the proximal ORS (28) and cells with CD29 (β1 integrin) appeared to be randomly distributed throughout the melanoma-associated chondrotin sulfate proteoglycan positive region of the ORS and were not restricted to any anatomical region (29). Moreover, This supports the data of Commo (9) who have also reported CD29 (β1 integrin) cells to be more widely expressed in the ORS.
We isolated CD34+ cells from hair-follicle cells by using magnetic bead and then CD34+ and CD34− cells isolated were confirmed by using fluorescent activated cell sorter (FACS). The purity of the isolated human hair follicle CD34+ cells was greater than 90%. We also analyzed immunological characterization of human hair follicle CD34+ cells. Our result show that most of CD34+ cells in human hair follicle expressed CD133. According to recent report, CD133 is expressed on immature hematopoietic stem and progenitor cells, and is not found on mature blood cells. (30). In addition to expression on hematopoietic stem/progenitor cells, recent reports suggest that CD133 also serves as a marker for stem/progenitor cells of the nonhematopoietic system, such as NSC (neural stem cells) (31), embryonic stem cell lines (32) and adult stem cells with pluripotent differentiation capacity (33). We also showed CD29 (β1 integrin) is expressed in most of CD34+ and CD34− cells. β1 integrin is expressed in all basal keratinocytes and it has been shown that as keratinocytes leave the basal layer, they down-regulate the expression of β1 integrins (34). Human keratinocytes can be analysed on the basis of whether they are rapidly (within 20 min) or slowly adherent to the β1 integrin ligand, type IV collagen. The rapidly adhering cells are found to have a high proliferative potential, whereas cells that adhere slowly divide only a few times before all of their progeny undergo terminal differentiation. Thus, rapidly adherent cells resemble stem cells, while the slowly adherent cells behave like transit-amplifying cells. This observation holds true whether cells are tested directly after isolating them from epidermis, or if they have been in culture (35). There is evidence to suggest that β1 integrin is required to maintain keratinocytes in an un-differentiated state. When keratinocytes are placed in suspension, they initiate terminal differentiation. This can be inhibited by ligating β1 integrin (36). An analysis of mutations in the β1 integrin subunit for their ability to regulate suspension-induced differentiation indicated that the signal transduced by β1 integrins is an instruction, do not differentiate (37). Interestingly, Levy et al. also demonstrated that the regulation of terminal differentiation is independent of β1 integrin-mediated adhesion (37).
The hair follicle represents a self-renewing system, which has been demonstrated to contain an intrinsic stem cell pool allowing the regeneration of the hair follicle (38). Recently, molecular markers have been described, which allow the identification and purification of hair follicle stem cells (16). The possibility to isolate and expand stem cells of different compartments of the hair follicle allows in vivo studies of hair follicle induction in animal models. Even if it was demonstrated by Trempus (16) that cell surface marker CD34 can be used for physical enrichment of a population of follicular cells with stem or progenitor cell characteristics, capacity of CD34+ cells for hair matrix is not fully understood in vivo. These studies demonstrated hair growth by freshly isolated hair follicle cells as well as by in vitro expanded hair follicle cells (39). However, the hair follicle cells used these studies were not limited stem cell population but whole population of hair follicle. We isolated CD34+ cells from human scalps and inject to the intradermis of nude mice. Based on our observation that new hairs were observed to mice head which had transplanted hair follicle CD34+ cells at 12 days, CD34− cells at 21 days after injection of the cells whereas physiologic saline-injected mice (Control group) fail to produce hair in the skin, we conclude that both human hair follicle CD34+ and CD34− cells can regenerate in vivo, however, CD34+ cells were able to regenerate much more hair follicle and earlier than CD34− cells. In histological examination, the results that new hair follicles were emerged in mouse skin transplanted with human hair follicle CD34+ cells also support mouse morphologic results and also, in this study, we demonstrated that human hair follicle CD34+ cells injected to nude mouse were located in outer root sheath with bulge region in the nude mouse hair follicle. Taken together, although it is unclear whetherthe regeneration due to paracrine actions to promote mouse hair follicle stem cells or direct regeneration response of human hair follicle CD34+ cells. These findings demonstrate that the transplanted cells play an important role in regeneration of hair follicle in nude mouse.
We also studied changes of protein level of mouse skin after transplantation with human hair follicle CD34+ cell and showed that active ERK level was significantly increased after transplantation. It was suggested that the hair follicle may be regenerated through active ERK in mouse skin. Also, the increased Bcl-2 level and decreased Bax level in mouse skin after transplantation endorses the assumption that the cells located in the bulge of human hair follicle have Bcl-2-positive and Bax negative phenotype (40).
Our results that CD34+ cells isolated from human hair follicle can produce new hair follicle in nude mice will also make a contribution towards cell-based approaches for treating alopecia are likely to use autologous tissue for bioengineering hair follicles to avoid immune rejection of donor cells.
Ohyama et al. recently demonstrated that CD34 expression is low or absent in human bulge ORS keratinocytes, suggesting that this molecule is not a marker of the human bulge area, but, CD34 expression is high in human sub-bulge region. Divergent CD34 expression between humans and mice suggests that substantial molecular and biological differences exist in bulge keratinocytes among mammalian species. In our results, CD34+ cells have more property regenerated new hair follicle in nude mice than CD34− cells. Therefore, we suggest that the human sub-bulge area with CD34+ cells may be the repository of multipotent stem cells that support hair follicle cycling and repopulate hair matrix than bulge region.
Taken together, our findings demonstrate that the human hair follicle cells with cell surface marker CD34 were located in the outer root sheath of nude mouse after transplantation and the cells can regenerate new hair follicle in immunodeficient nude mouse. CD34− cells also can form follicles in the mouse and the expression level of β1 integrin is high, same as CD34+ cells however CD34+ cells were able to regenerate much more hair follicle than CD34− cells. Moreover, the results of this study add new insight into the investigation of CD34 stem cell-related molecule in hair follicles and suggest that not all human hair follicle stem cells reside in bulge region, but in a lager niche. Although the way how the cells go to outer root sheath of nude mouse skin is unknown, further investigation of the functional properties of this CD34+ cell may well provide insight into stem cell biology.
Go to :
 XML Download
XML Download