

 PDF
PDF Citation
Citation Print
Print


Introduction
The special characteristics of stem cells (i.e., self-renewal capacity and pluripotency) that allow them to differentiate into any of three germ layer lineages (1, 2) can be regulated by a unique microenvironment called the stem cell niche, which consists of physicochemical and physiological niches, and the extracellular matrix (ECM) (3, 4). To date, the defined manipulation of physicochemical and physiological niches has been limited in its ability to precisely and specifically guide the fate of stem cells (5, 6). Therefore, the importance of niches based on ECM has been emphasized as an alternative to overcoming the difficulty of regulating the fate of stem cells.
The ECM is a non-cellular three-dimensional macromolecular network composed of a variety of fibrous ECM proteins, carbohydrates, and several glycoproteins (7, 8), which not only provide a physical scaffold but also signals to induce cellular responses including proliferation, migration, attachment, spreading, differentiation, survival, homeostasis, and morphogenesis (9). The integrins, a group of cell adhesion molecules that exist as heterodimers with various combinations of α and β subunits (10, 11), play pivotal roles as mediators in transporting ECM-derived signals into the cytoplasm by directly recognizing ECM components and inducing cytological changes (12). Accordingly, the combined stimulation of ECM protein-derived signals through integrin heterodimers can make it possible to accurately regulate the fate of stem cells, which requires detailed information on integrin heterodimers that are functionally expressed on the surface of stem cells.
Among the diverse stem cell types, spermatogonial stem cells (SSCs) are the only male germline stem cells that differentiate into sperm through spermatogenesis in the seminiferous tubule of the testis (13, 14). Their self-renewal is infinitely maintained on the seminiferous tubule basement membrane (STBM) with defined ECM components in vivo (15). Nevertheless, technical difficulties in constructing the artificial STBM has led to the use of a cellular niche based on feeder cells during the in vitro culture of SSCs (16-18). However, a small population of SSCs located in the seminiferous tubule of the testis derived from diverse species did not sufficiently expand under these conditions (19). Simultaneously, non-cellular niches using purified ECM proteins, not based on information regarding integrin heterodimers expressed in the undifferentiated SSCs also did not show efficient long-term in vitro maintenance of SSC self-renewal.
Outbred stocks of mice are widely used in toxicology, pharmacology, and fundamental biomedical research (20), although most such studies can be done much more effectively with inbred strains. Particularly, their usage in genetic studies has been very successful as a base population for selection to produce new or improved humanized mouse models (21), and for genetic fine mapping of quantitative trait loci (QTL) (22). In addition, their better breeding performance compared to inbred strains makes it easy to develop and maintain genetically modified mice (23). However, despite their usefulness, to date, there have been no reports of in vitro culture systems for SSCs derived from outbred stocks of mice.
Accordingly, as a step towards developing a precisely defined non-cellular niche engineering integrin signaling to promote self-renewal of SSCs derived from outbred mice, we examined the types of integrin heterodimers on the surface of outbred ICR mouse SSCs in the undifferentiated state. The types of integrin subunits expressed in undifferentiated SSCs were identified at the transcriptional and translational levels, and the combinations of integrin α and β subunits were determined by functional assays.
Materials and Methods
Animals
Three-week-old male ICR mice were purchased from DBL (Eumseong, Korea) and used as SSC donors. All of the animal housing, handling, and experimental procedures were approved by the Institutional Animal Care and Use Committee (IACUC) of Kangwon National University (IACUC approval No. KW-130307-1) and conducted in accordance with the Animal Care and Use Guidelines of Kangwon National University.
Isolation of SSC population from the testes
The tunica albuginea and epididymis were removed from the mouse testes, and the seminiferous tubules were digested by incubation for 20 min in Dulbecco’s Modified Eagle’s Medium (DMEM; Welgene Inc., Daegu, Korea) supplemented with 0.5 mg/ml type IV collagenase (Worthington Biochemical, Lakewood, CA) at 37℃. Subsequently, the fragmented seminiferous tubules were washed with DMEM containing 10% (v/v) heat-inactivated fetal bovine serum (FBS; Welgene), and dissociated with 0.25% trypsin-EDTA (Welgene) for 5 min at 37℃. The dispersed testicular cells were filtered through a 70 μm nylon strainer (SPL, Pocheon, Korea) to eliminate peritubular myoid, sertoli, and leydig cells, and the filtered testicular cells were purified by magnetic-activated cell sorting (MACS; Invitrogen, Carlsbad, CA) using anti-Thy1 antibody (Novus Biologicals, Littleton, CO) according to the manufacturer’s instructions. Subse-quently, the finally sorted cells (herein referred to as SSC population) were assigned to the following experiments.
Real-time PCR
Total mRNA was extracted from the cells using a Dynabeads mRNA DirectTM Kit (Ambion, Austin, TX), and cDNA was synthesized using ReverTra Ace qPCR RT Master Mix with gDNA remover kit (Toyobo, Osaka, Japan) in accordance with the respective manufacturer’s instructions. Subsequently, specific gene expression was quantified with THUNDERBIRDTM SYBRⓇ qPCR Mix (Toyobo) using a 7500 Real-time PCR system (Applied Biosystems, Foster City, CA). PCR specificity was determined by analyzing the melting curve data. The mRNA levels are presented as 2−ΔCt where Ct=threshold cycle for target amplification and ΔCt=Cttarget gene (specific genes for each sample) – Ctinternal reference (β-actin for each sample). Primer sequences were designed with Primer3 software (Whitehead Institute/MIT Center for Genome Re-search) with mouse cDNA sequence data obtained from GenBank. General information and sequences of primers are described in Supplementary Table S1.
Immunocytochemistry
Cells fixed with 4% (v/v) paraformaldehyde (Junsei, Tokyo, Japan) for 10 min were washed twice with Dulbecco’s phosphate-buffered saline (DPBS; Welgene). The fixed cells were stained with fluorescence-conjugated integrin α1, α5, α6, α9, αV, αE, β1, and β5 primary antibodies diluted in DPBS for 16 h at 4℃. After rinsing with DPBS, the stained cells were double stained by fluorescence-unconjugated glial cell line-derived neurotrophic factor (GDNF) family receptor alpha 1 (GFRα1; a SSC-specific marker) primary antibody diluted in DPBS, or florescence-unconjugated alpha smooth muscle actin (αSMA; a peritubular myoid cell-specific marker), GATA binding protein 4 (GATA4; a sertoli cell-specific marker), or luteinizing hormone receptor (LHR; a leydig cell-specific marker) primary antibody diluted in Hank’s balanced salt solution (HBSS, Invitrogen) containing 0.1% (w/v) saponin (Sigma-Aldrich, St. Louis, MO) for 16 h at 4℃. Subsequently, the detection of GFRα1 primary antibody was conducted by incubating Alexa Fluor 488- or 546-conjugated secondary antibody diluted in DPBS for 2 h at 4℃, and the detection of αSMA, GATA4, and LHR primary antibodies was conducted by incubating Alexa Fluor 488-, 546- or 568-conjugated secondary antibody diluted in HBSS containing 0.1% (w/v) saponin for 1 h at 4℃. Supplementary Table S2 describes the detailed information and dilution rate of the used antibodies. Then, rinsing twice with DPBS was performed and the double stained cells were counterstained with mounting medium for fluorescence with 4’,6-diamidino-2-phenylindole (DAPI) (Vector Laboratories, Inc., Burlingame, CA). Moreover, the triple-stained cells were monitored under a confocal laser scanning microscope (LSM880; ZEISS, Jena, Germany).
Fluorescence immunoassay
Three thousand three hundred of cells fixed in 4% (v/v) paraformaldehyde (Junsei) for 10 min were rinsed twice with DPBS. Subsequently, the fixed cells were stained for 2 h at room temperature with FITC-conjugated anti-mouse integrin α5, α9, αV, αE, and β1 and Alexa Fluor 488-conjugated anti-mouse integrin α6 antibodies diluted in DPBS. The detailed information and antibody dilutions used are listed in Supplementary Table S2. The stained cells were washed with DPBS, and fluorescence intensity was measured using SoftMaxⓇ Pro 6.2.2. (Molecular Devices Corp., Sunnyvale, CA) after adding 100 μl DPBS to the stained cells.
Attachment assay
First, 96-well tissue culture plates were coated with the following concentrations of purified ECM proteins overnight at 4℃: 0, 40, and 80 μg/ml fibronectin (Millipore, Burlington, MA); 0, 200, and 400 μg/ml laminin (Sigma-Aldrich); 0, 20, and 40 μg/ml tenascin C (RayBiotech, Peachtree Corners, GA) and 0, 5, and 10 μg/ml vitronectin (R&D Systems, Minneapolis, MN). Then, each well blocked with 1% (w/v) bovine serum albumin (BSA; Sigma) at 4℃ for 1 h was washed three times with DPBS. Aliquots of 1×104 cells were resuspended in SSC culture medium consisting of Stempro-34 medium (Invitrogen) supplemented with insulin–transferrin–selenium (ITS; Invitrogen), 60 μM putrescine dihydrochloride (Sigma), 6 mg/ml D-(+)-glucose (Wako Pure Chemical Industries, Ltd., Osaka, Japan), 0.11 mg/ml sodium pyruvate (Sigma), 1 μl/ml DL-lactic acid (Sigma), 5 mg/ml BSA, 2 mM L-glutamine (Invitrogen), 0.05 mM β-mercaptoethanol (Invitrogen), 1% (v/v) FBS, 1% (v/v) NEAA (Invitrogen), MEM vitamin solution (Sigma), 1% (v/v) antibiotic–antimycotic solution (Welgene), 0.1 mM ascorbic acid (Sigma), 103 units/ml mouse LIF (Chemicon International Inc., Temecula, CA), 10 ng/ml GDNF (R&D systems), 30 ng/ml β-estradiol (Sigma), 60 ng/ml progesterone (Sigma), 20 ng/ml human epidermal growth factor (EGF; Peprotech, Inc., Rocky Hill, NJ), and 10 ng/ml human basic fibroblast growth factor (bFGF; Peprotech, Inc.), and seeded in each well. After incubation at 37℃ for 2 h, non-adherent cells were removed by washing each well sufficiently, and adherent cells were fixed in 4% (v/v) paraformaldehyde at room temperature for 10 min. Subsequently, the fixed adherent cells were stained with 0.1% (w/v) crystal violet (Sigma) in 20% (v/v) methanol (Sigma-Aldrich) for 5 min and washed twice with distilled water. Levels of adherent cells were quantified at 570 nm using a microplate reader (Epoch Microplate Spectrophotometer; BioTek Instruments Inc., Winooski, VT) after adding 50 μl of 0.2% (v/v) Triton X-100 (Biopure, Cambridge, MA) diluted with distilled water.
Antibody inhibition assay
The wells of 96-well tissue culture plates were coated with 40 μg/ml fibronectin, 200 μg/ml laminin, 20 μg/ml tenascin C or 5 μg/ml vitronectin overnight at 4℃, and the coated wells were blocked with 1% (w/v) BSA for 1 h at 4℃. Subsequently, to inhibit integrin function, 1×104 cells were resuspended in SSC culture medium containing anti-integrin α5 (5H10–27 [MFR5]), anti-integrin α6 (NKI-GoH3), anti-integrin α9 (Y9A2) or anti-integrin αV (RMV-7) blocking antibody, and incubated at 37℃ for 2 h. The detailed information regarding the antibodies used is presented in Supplementary Table S2. Then, the functionally blocked cells were plated in each well and incubated at 37℃ for 8 h. To remove non-adherent cells, the wells were washed extensively with DPBS. The adherent cells were fixed in 4% (v/v) paraformaldehyde for 10 min at room temperature, and staining of adherent cells was performed with 0.1% (w/v) crystal violet in 20% (v/v) methanol for 5 min. Finally, the wells were washed twice with distilled water and supplemented with 50 μl of 0.2% (v/v) Triton X-100 diluted with distilled water. The amount of dye was measured at 570 nm using a microplate reader.
Statistical analysis
The SAS program was used for statistical analysis of the numerical data shown in each experiment. Comparisons among treatment groups were performed by the least-squares or DUNCAN method, and the significance of the main effects was determined by analysis of variance (ANOVA) in the SAS package. In all of the analyses, p< 0.05 was taken to indicate statistical significance.
Results
Identification of the types of cells isolated from outbred ICR mouse testes through MACS technique
A MACS technique based on antibody detecting Thy1 expressed on the surface of undifferentiated mouse SSCs did not show high yield in the isolation of undifferen-tiated SSCs from outbred ICR mouse testes. As shown in Supplementary Fig. S1, SSCs stained positively with GFRα1, peritubular myoid cells stained positively with αSMA, sertoli cells stained positively with GATA4, and leydig cells stained positively with LHR were detected in the retrieved SSC population post-MACS. Moreover, among four cell types included in the SSC population, SSCs (24.57±3.12%) and peritubular myoid cells (28.68±3.20%) showed significantly higher rate of abundance than sertoli (16.39±1.06%) and leydig (11.32±4.80%) cells. Therefore, in order to gain insight into the types of integrin subunit expressed on the membrane of outbred ICR mouse SSCs in the undifferentiated state, we investigated the transcriptional levels of integrin subunit genes in the SSC population and the translation of integrin subunit genes showing significantly strong transcription were identified per each type of cells through immunocytochemistry.
Transcriptional levels of integrin subunit genes in the SSC population derived from outbred ICR mouse testes
In transcriptional analyses of genes encoding a total of 25 integrin subunits, significantly higher levels of expression were observed for integrin α1, α5, α6, α9, αV, and αE (Fig. 1A), and integrin β1 and β5 (Fig. 1B) subunit genes. The minimum levels of expression were detected for integrin α2, α3, α4, α7, α8, α10, αL, αM, αD, and αX (Fig. 1A), and integrin β2, β3, β4, and β7 (Fig. 1B) subunit genes. No expression of integrin α11 (Fig. 1A) or integrin β6 and β8 (Fig. 1B) subunit genes was detected in these cells at the transcriptional level.
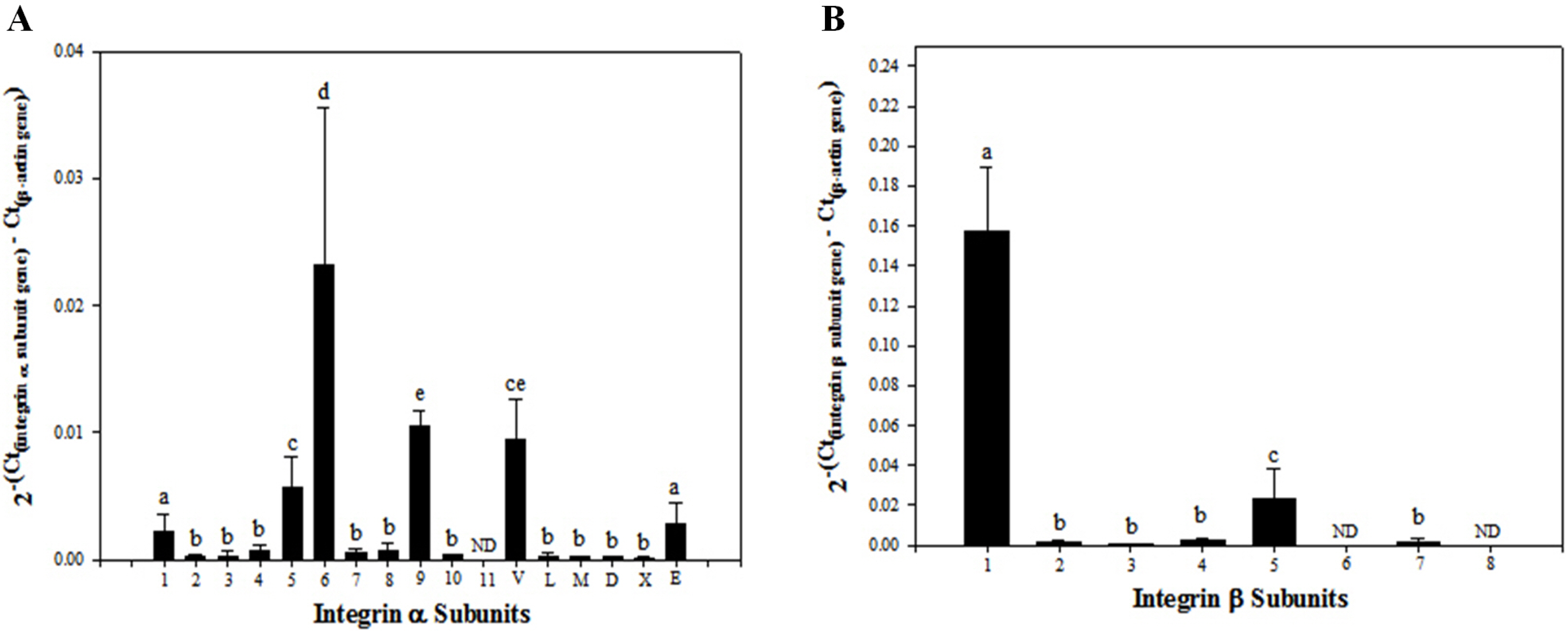
Fig. 1
Transcriptional levels of integrin α and β subunit gene expression in SSC population including undifferentiated SSCs and peritubular myoid, sertoli and leydig cells derived from outbred ICR mouse testes. Testicular cells were retrieved enzymatically from testis derived from ICR mice, and the preparation of SSC population was conducted by sorting these testicular cells using a MACS technique based on anti-Thy1 antibody. Subsequently, mRNA levels of integrin α and β subunits in the isolated SSCs were examined quantitatively by real-time PCR. A total of 6 of 17 integrin α subunit genes (α1, α5, α6, α9, αV, and αE) (A) and 2 of 8 integrin β subunit genes (β1 and β5) showed significantly increased expression (B). All data shown are means±standard deviation of three independent experiments. a∼ep<0.05. ND: not detected.
![]()
Determination of integrin subunits expressed on the membrane of undifferentiated outbred ICR mouse SSCs
Translational regulation of integrin α1, α5, α6, α9, αV, and αE, and integrin β1 and β5 subunit genes showing increased transcription was examined in each type of cells included in SSC population. As shown in Fig. 2, SSCs stained positively with GFRα1 showed co-localization of integrin α5, α6, α9, αV, αE and β1 subunit proteins on the cell surface, whereas co-localization of integrin α1 and β5 subunit proteins were not detected in GFRα1-positive SSCs. Furthermore, the surface of peritubular myoid (Supplementary Fig. S2), sertoli (Supplementary Fig. S3), or leydig (Supplementary Fig. S4) cells stained positively with αSMA, GATA4, or LHR did not show any co-localization of integrin α1, α5, α6, α9, αV, αE, β1, and β5 subunit proteins. Justly, co-localization of integrin α1 subunit protein was detected on the surface of LHR-positive leydig cells. These results demonstrate that dimerization of integrin α and β subunits as active forms of integrins can’t be absolutely formed on the surface of peritubular myoid, sertoli, and leydig cells, indicating the absence of their influence on the following functional assay using the retrieved SSC population post-MACS.
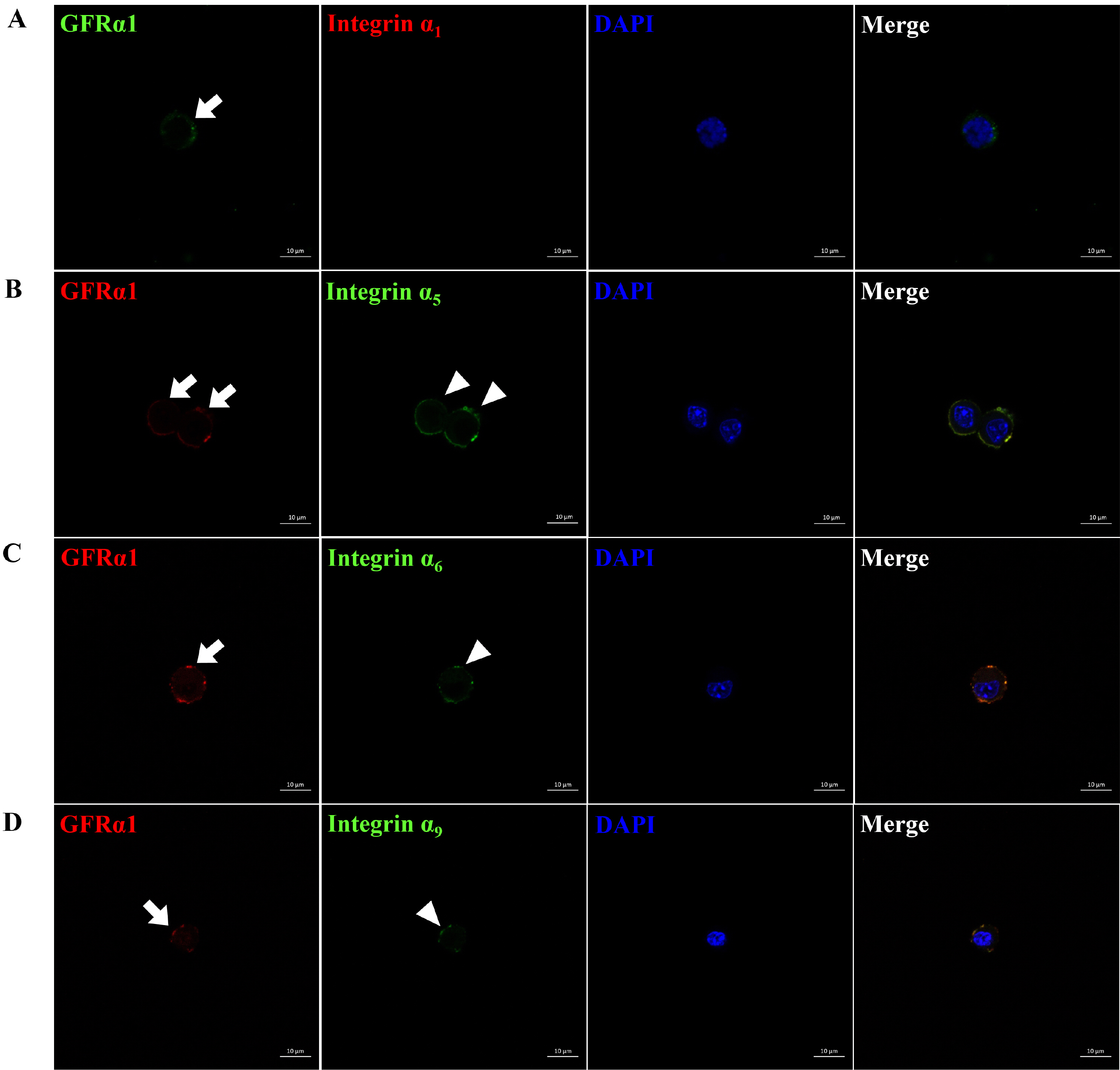
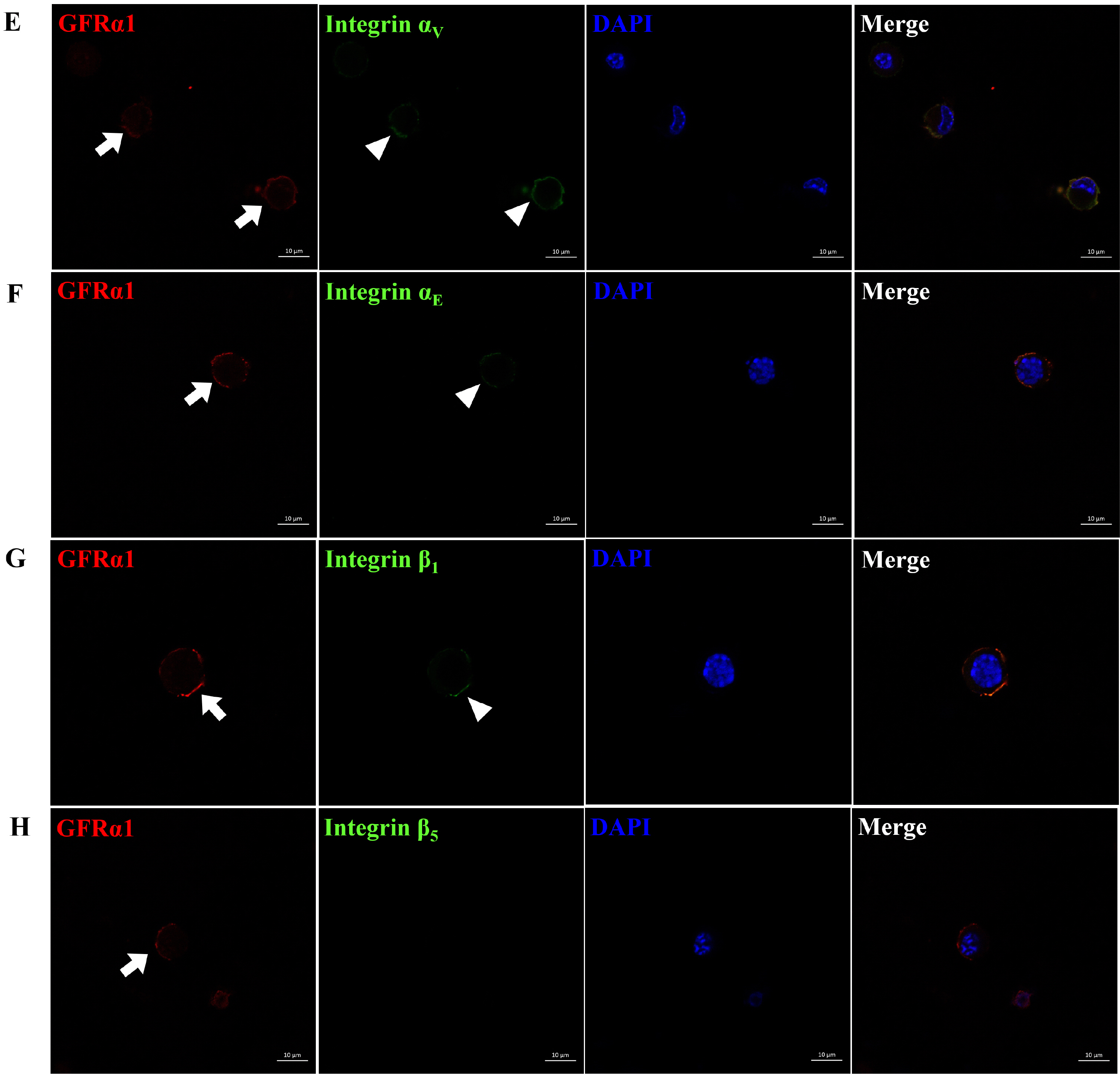
Fig. 2
Identification of integrin α and β subunit proteins expressed on the surface of undifferentiated SSCs derived from outbred ICR mouse testes. To obtain testicular cells, testis derived from ICR mice was enzymatically treated and the preparation of SSC population which the proportion of SSCs in testicular cells was increased was conducted using a MACS technique based on anti-Thy1 antibody. Subsequently, the protein expression of integrin α and β subunits in the SSCs included in the sorted SSC population were identified by immunocytochemistry. As the results, integrin α5, α6, α9, αV, αE and β1 subunit proteins (arrow head; B∼G) were localized on the surface of SSCs expressing GFRα1 (arrow; a SSC-specific marker), whereas any localization of integrin α1, and β5 subunit proteins (A, H) was not detected on the surface of SSCs expressing GFRα1. All figures are representative immunocytochemistry images of integrin subunit proteins expressed on the surface of SSCs. Nuclear counterstaining was conducted using DAPI. n=3. Scale bars represent 10 μm.
![]()
![]()
Subsequently, levels of integrin α5, α6, α9, αV, αE, and β1 subunit genes expressed translationally on the surface of undifferentiated SSCs s were quantified (Fig. 3). Among the six integrin subunit genes, integrin αV subunit gene had significantly the strongest translation expression, and integrin α5, α6, and β1 subunit genes had significantly intermediate translational expression, whereas integrin α9 and αE subunit genes showed significantly very weak expression at the translational level. These results indicate that integrin α5, α6, α9, αV, αE and β1 subunits are presented on the surface of outbred ICR mouse SSCs in undifferentiated state.
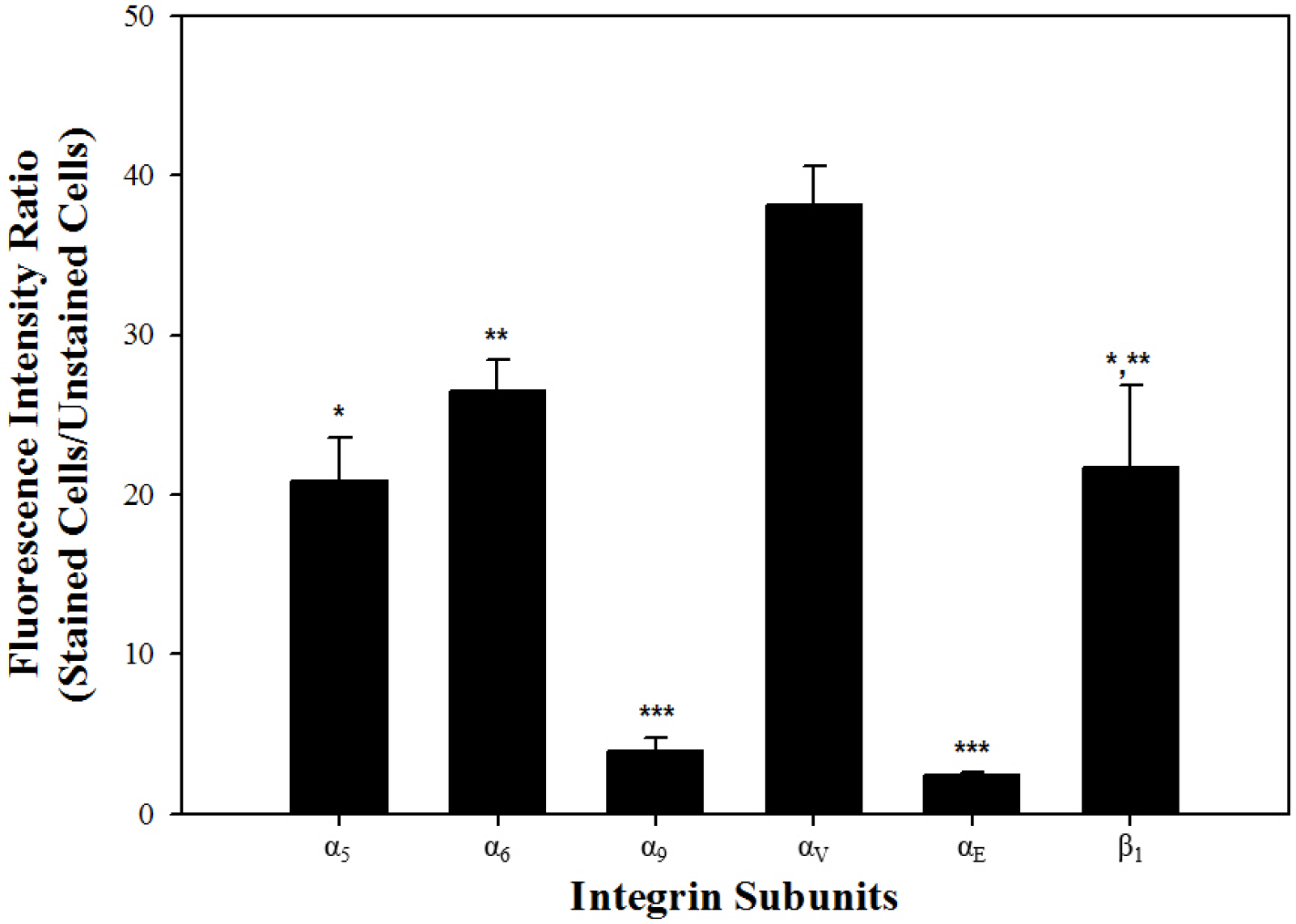
Fig. 3
Translational levels of integrin α and β subunit gene expression in undifferentiated SSCs derived from outbred ICR mouse testes. In order to prepare SSC population which the proportion of SSCs in testicular cells was increased, testicular cells retrieved enzymatically from testis derived from ICR mice were sorted using a MACS technique based on anti-Thy1 antibody. Subsequently, the protein expression levels of integrin α and β subunits in the SSCs included in the sorted SSC population was analyzed by fluorescent immunoassays, and at the translational level, each integrin subunit was represented as the ratio of fluorescence intensity of stained cells to that of unstained cells. Among the six integrin subunits expressed on the surface of undifferentiated SSCs, significantly the strongest translational expression of integrin αV subunit gene was detected, whereas integrin α9 and αE subunit genes showed significantly the weakest expression at the translational level. Moreover, integrin α5, α6 and β1 subunit genes showed significantly intermediate translational expression. All of the data shown are means±standard deviation of three independent experi-ments. *-***p<0.05.
![]()
Identification of integrin heterodimers functionally expressed on the cell membrane of undifferentiated outbred ICR mouse SSCs
Based on the results regarding integrin α and β subunits expressed on the cell membrane of undifferentiated outbred ICR mouse SSCs, we speculated that these cells may possess integrin α5β1, α6β1, α9β1, and αVβ1 as active forms of integrin heterodimers as previously described (24). The presence of these integrin heterodimers was examined by estimating levels of adherent SSCs cultured on purified ECM proteins that interact specifically with each integrin heterodimer and adherent levels post-culture of SSCs treated with antibodies specifically blocking each integrin function on each purified ECM protein. Fig. 4 shows significantly improved adhesion for SSCs incubated on fibronectin-, laminin-, tenascin C- and vitro-nectin-coated culture plates, compared to those cultured on purified ECM protein-free culture plates. These observations suggested the presence of the fibronectin-interacting integrin α5β1, laminin-interacting integrin α6β1, tenascin C-interacting integrin α9β1, and vitronectin-interacting integrin αVβ1 on the cell membrane of SSCs in the undifferentiated state. These specific integrin function-blocked SSCs were incubated on 40 μg/ml fibronectin, 200 μg/ml laminin, 20 μg/ml tenascin C, or 5 μg/ml vitronectin as the minimum concentration among those seen in the ECM showing significantly improved adhesion of SSCs (Fig. 5). Significantly weakened adhesion was detected in SSCs with blockade of integrin α5β1 (Fig. 5A), α6β1 (Fig. 5B), α9β1 (Fig. 5C), or αVβ1 (Fig. 5D), compared to those without blocking of these integrin heterodimers. These results confirmed that the undifferentiated outbred ICR mouse SSCs simultaneously show functional expression of integrin α5β1, α6β1, α9β1, and αVβ1 on the cell membrane.
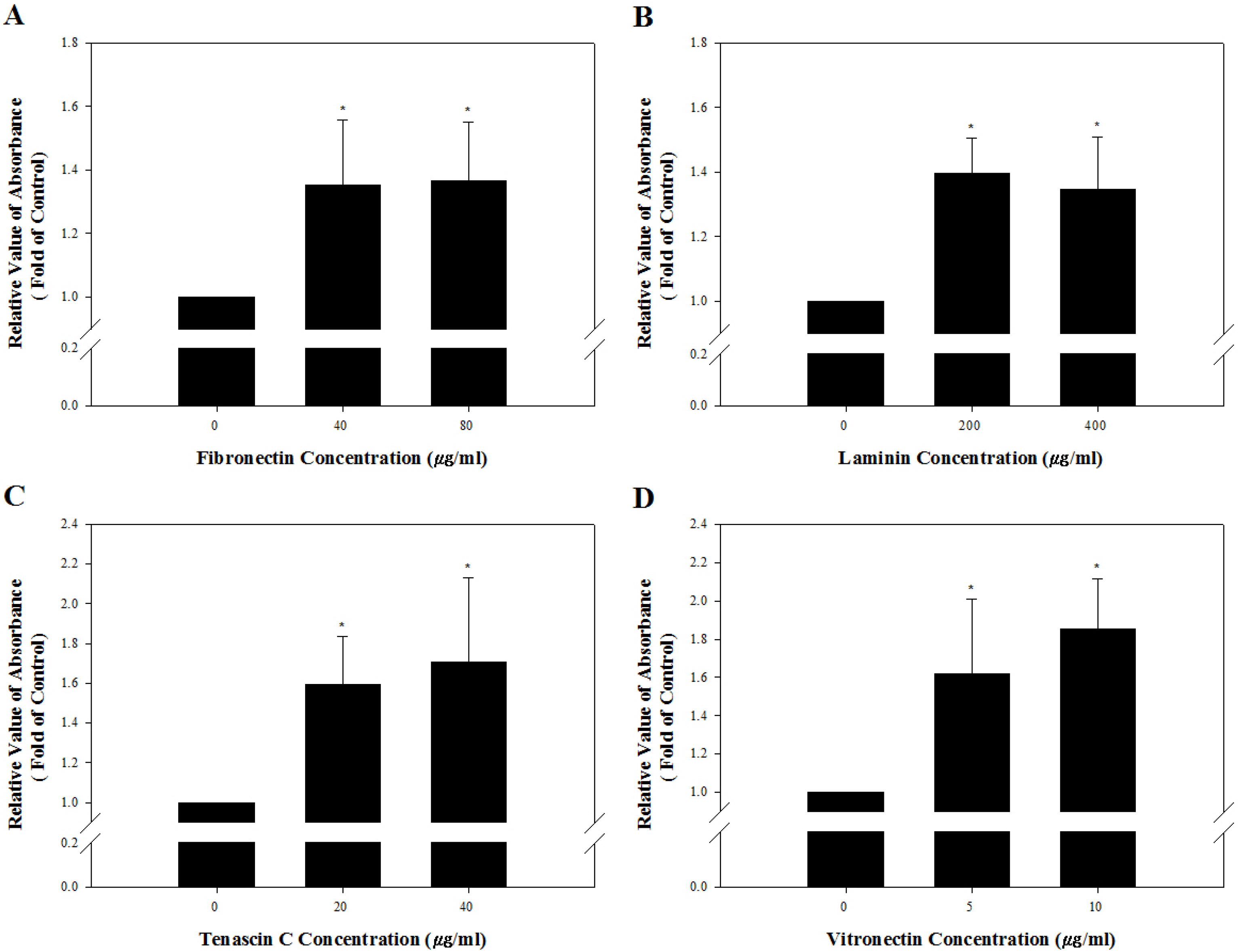
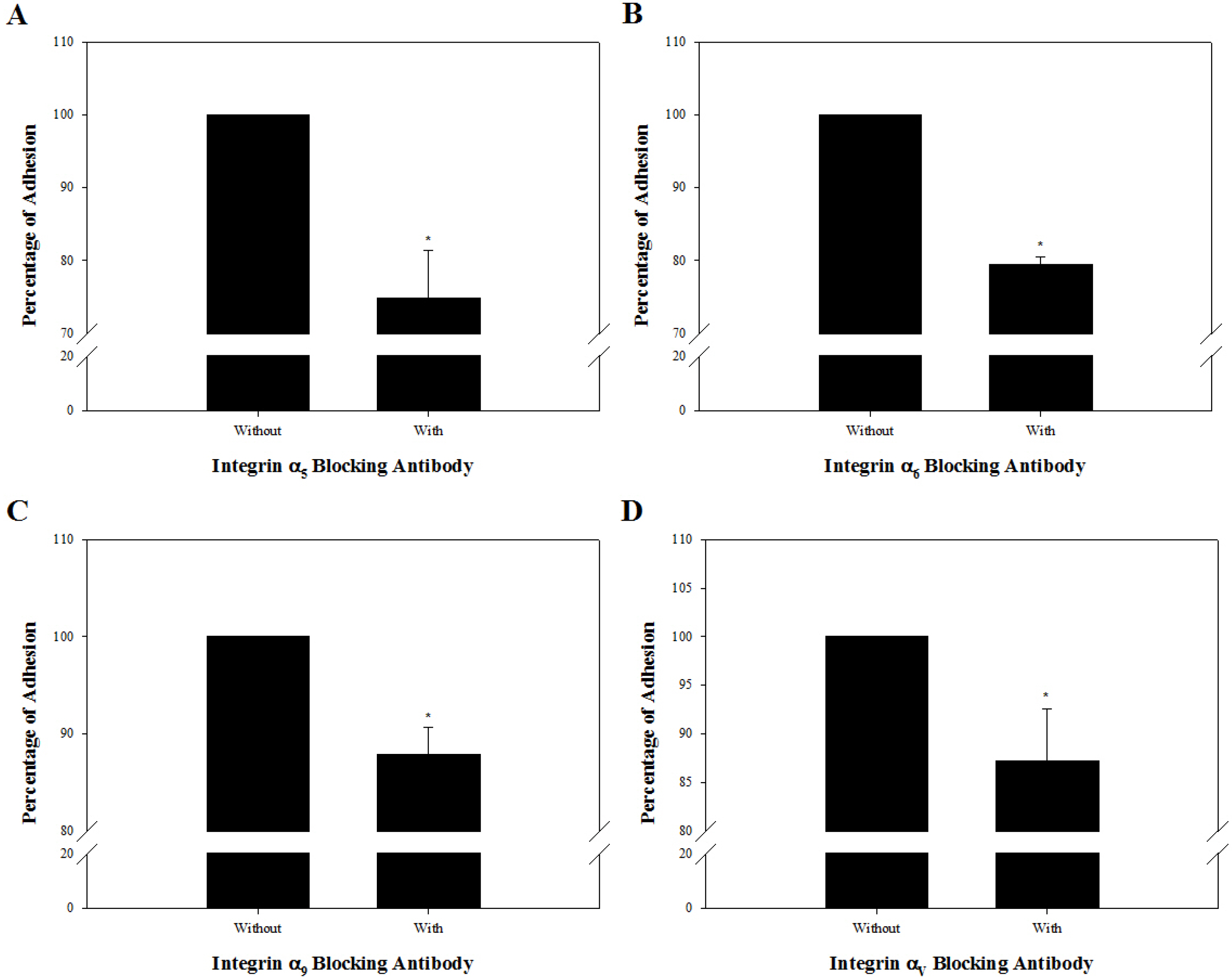
Fig. 4
Identification of the presence of integrin heterodimers interacting with fibronectin, laminin, tenascin C and vitronectin on the cell membrane of undifferentiated outbred ICR mouse SSCs. A 96-well tissue culture plates was coated with 0, 40, or 80 μg/ml fibronectin (A), 0, 200, or 400 μg/ml laminin (B), 0, 20, or 40 μg/ml tenascin C (C), and 0, 5, or 10 μg/ml vitronectin (D). Subsequently, SSC population were prepared by sorting testicular cells retrieved enzymatically from testis derived from ICR mice using a MACS technique based on anti-Thy1 antibody. Then, 1×104 cells in SSC population were resuspended in SSC culture medium and plated in each well. After incubation for 2 h at 37℃, adherent cells were stained with crystal violet, and the adhesion level was quantified using a microplate reader. The percentage of maximum adhesion is represented as the optical density of cells plated on ECM protein-free plates. Mouse SSCs cultured on fibronectin-, laminin-, tenascin C- and vitronectin-coated culture plates had significantly higher levels of adhesion than those on ECM protein-free culture plates. However, increasing concentrations of ECM on the culture plates did not induce a significant improvement of mouse SSC adhesion levels. All of the data shown are means±standard deviation of three independent experiments. *p<0.05.
![]()
Fig. 5
Functional analysis of integrin heterodimers suspected to function on the cell membrane of undifferentiated outbred ICR mouse SSCs. SSC population were prepared by sorting testicular cells retrieved enzymatically from testis derived from ICR mice using a MACS technique based on anti-Thy1 antibody. Subsequently, 1×104 cells in SSC population were incubated in the absence or presence of anti-integrin α5 (5H10–27 [MFR5]) (A), anti-integrin α6 (NKI-GoH3) (B), anti-integrin α9 (Y9A2) (C), or anti-integrin αV (RMV-7) (D) blocking antibody, plated on 40 μg/ml fibronectin-, 200 μg/ml laminin-, 20 μg/ml tenascin C, and 5 μg/ml vitronectin-coated wells, and incubated for 8 h at 37℃. After staining adherent cells with crystal violet, quantification of adhesion level was conducted using a microplate reader. As a parameter of functional blocking by antibodies, the percentage of maximum adhesion, which is represented by the optical density of cells plated on each ECM protein-coated well in the absence of any blocking antibodies, was determined. Mouse SSCs treated with integrin α5β1, α6β1, α9β1, and αVβ1 blocking antibodies showed significantly lower rates of attachment to each ECM component compared to mouse SSCs without blocking antibody. All of the data shown are means±standard deviation of three independent experiments. *p<0.05.
![]()
Discussion
In constructing niches for driving the self-renewal or differentiation of SSCs, information regarding the extracellular signals in the microenvironments around undifferentiated or differentiated SSCs is essential. Parti-cularly, extracellular signals derived from ECM proteins composing STBM that interact directly with SSCs in the seminiferous tubule (25) play an important role in determining the fate of these cells. However, intracytoplas-mic transduction of these ECM-derived signals is mediated by cell-surface receptors belonging to the integrin family (26). Therefore, information on integrins present in undifferentiated or differentiated SSCs should be reflected in fabricating niches customized to each type of SSC. Here, we confirmed the types of integrin heterodimers expressed on the surface of undifferentiated SSCs from outbred ICR mice. Through transcriptional analysis of 24 integrin subunits, followed by confirmation of their expression at the translational level, attachment to ECM proteins, and inhibition with blocking antibodies, integrin α5β1, α6β1, α9β1 and αVβ1 were found to be expressed on the cell membrane. These results suggest that the fibronectin-interacting integrin α5β1, laminin-interacting integrin α6β1, tenascin C-interacting integrin α9β1, and vitronectin-interacting integrin αVβ1 may be valuable extracellular signals in maintaining outbred ICR mouse SSCs in the undifferentiated state, and that niches incorporating fibronectin, laminin, tenascin C and vitronectin analogs may provide support for self-renewal of SSCs derived from outbred ICR mice.
The basement membrane in the seminiferous tubule consists of fibronectin (27), laminin (28), collagen IV (29, 30), entactin (31, 32), and perlecan (33). Accordingly, the expression of integrin α5β1 and α6β1 on the surface of the undifferentiated outbred ICR mouse SSCs (Fig. 4, 5) suggest that fibronectin and laminin within the STBM may be key extracellular signals for maintaining the self-renewal of outbred ICR mouse SSCs. Interestingly, outbred ICR mouse SSCs also expressed integrin α9β1 and αVβ1 in the undifferentiated state (Fig. 4, 5). However, there are no tenascin C or vitronectin interacted with integrin α9β1 or αVβ1 within the STBM. Therefore, we speculated that the tenascin C- and vitronectin-derived extracellular signal may be derived from the ECM of neighboring SSCs, supporting maintenance of outbred ICR mouse SSC self-renewal by combining with extracellular signals derived from fibronectin and laminin within the STBM. In contrast, integrin α9β1 and αVβ1 may be an inducer of spermatogenesis due to its localization on the membrane as active heterodimers rather than inactivated subunits (Fig. 4, 5), as supported by previous reports that tenascin C and vitronectin is expressed in the seminiferous tubules of the adult testes showing normal spermatogenesis (34, 35). However, further studies are required to validate these speculations.
In conclusion, transcriptional, translational, and functional screening of integrins demonstrated the expression of integrin α5β1, α6β1, α9β1 and αVβ1 on the surface of the undifferentiated SSCs derived from outbred ICR mice. The identified integrin heterodimers may be useful for maintaining mouse SSCs from outbred strains in undifferentiated state. Moreover, these achievements will greatly improve the applicability of niches with regard to the maintenance of SSC self-renewal.
Supplementary Materials
Supplementary data including two tables and four figures can be found with this article online at http://pdf.medrang.co.kr/paper/pdf/IJSC/IJSC-13-s20061.pdf.
 XML Download
XML Download