

 PDF
PDF Citation
Citation Print
Print


Structure and Function of Tendon and Ligament
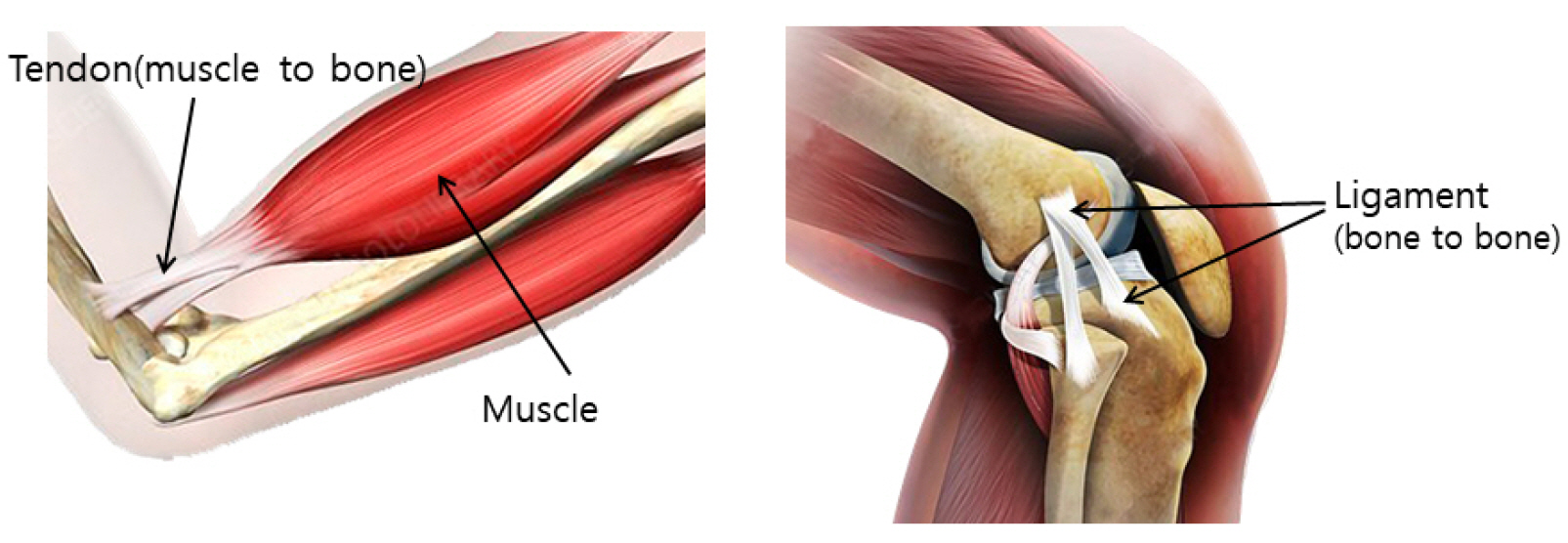
Tendons are structures connecting muscles to the bones in our body and transmit the force generated by contraction of the muscles to the bones. Ligaments connect bones to bones, with histological properties similar to tendons. Both tendons and ligaments can preserve elastic energy and withstand high tension (Fig. 1).
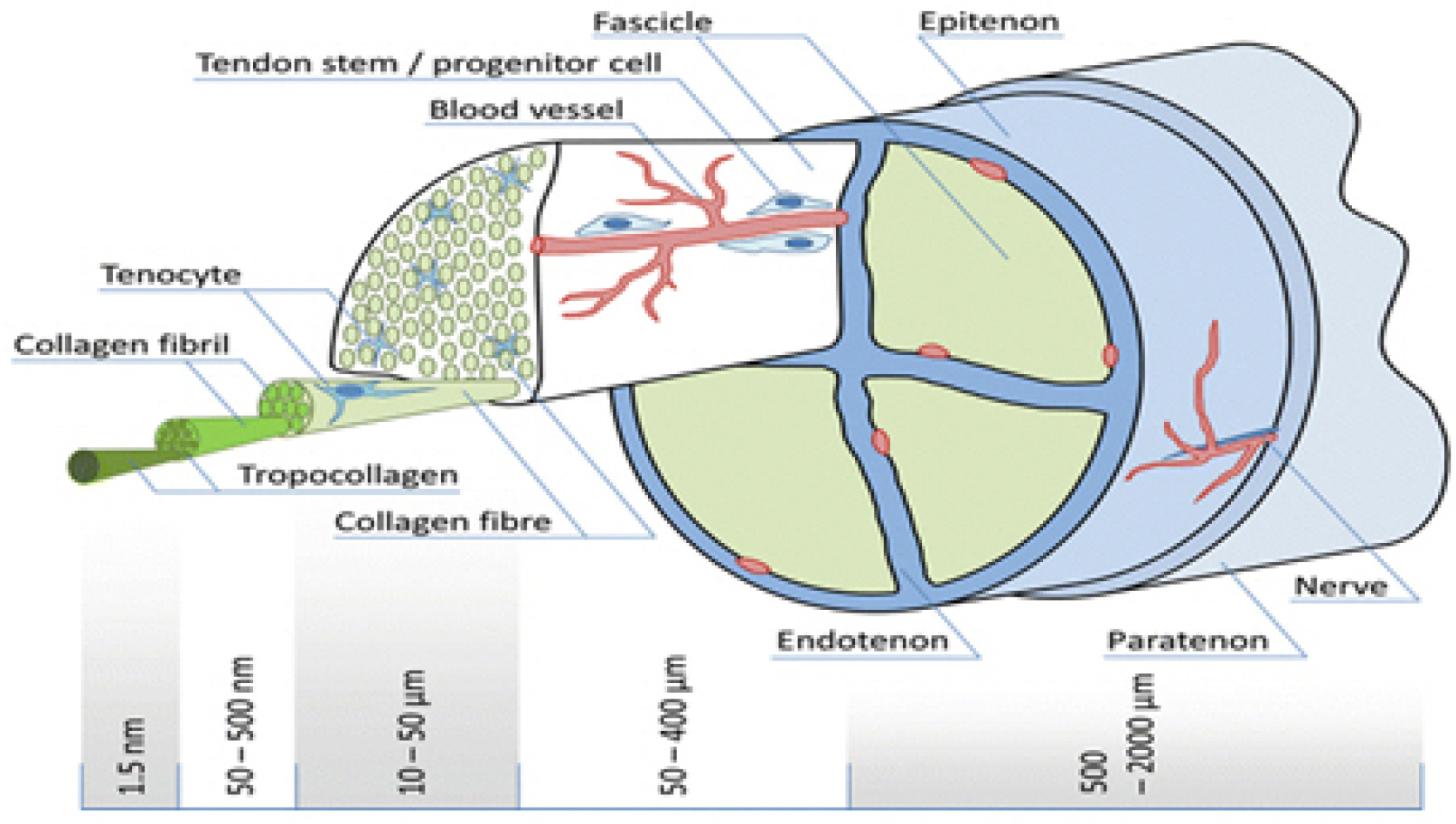
In the microstructure of the tendon, three tropocollagen chains are intertwined with one another to form microfi-brils. The microfibrils are stacked in parallel to form fibrils, and the fibrils gather to form fascicles that are wrapped by endotenon. The fascicles gather to form tendons, which are surrounded by epitenon. Epitenon is wrapped by paratenon, which contains blood vessels and nerves that feed the tendon (Fig. 2).
Collagen primarily provides resistance to tensile forces. While type 1 collagen comprises 95% of the whole collagen, there are also types 3, 5, 6, 12, 14, and 15 collagen. Type 3 collagen is the most common type of collagen in tendons after type 1 collagen. Type 3 collagen tends to increase in the course of tendinitis or tendon healing. Elastin endows an elastic property to tendons. Type 1 collagen accounts for 65∼80% of the dry weight of tendons, while elastin contributes 2%. Extracellular matrix (ECM) provides lubrication and viscoelasticity to the tendon and maintains the fibrous structure. The ECM of tendon is comprised of glycoproteins, including tenascin, decorin (DCN), biglycan (BGN), fibromodulin (FMOD), and lumican (LUM).
Tenomodulin (Tnmd) is a type 2 transmembrane glycoprotein specific to the tendon that plays an important role in cell proliferation and tendon maturation. Tnmd is regulated by scleraxis (SCX), a tendon-specific transcription factor. When Tnmd is deficient, the proliferative capacity of tendon progenitor cells decreases, and aging progresses rapidly. As it is not found in other musculoskeletal cells, Tnmd is a good marker of tenogenic differentiation.
In normal tendons, the fibers are parallel and there are few cells. In tendinosis, the number of cells increases with the proliferation of blood vessels and mucus degeneration of the tendon substance (1-3).
Go to : 
Tendon/Ligament-Specific Stem Cells
In tendon and ligament tissue, there are very small numbers of cells similar to mesenchymal stem cells (MSCs), called tendon stem/progenitor cells (TSPCs) or tenogenic stem cells (4, 5). TSPCs have all the properties of general MSCs, such as the presence of specific surface markers, self-replication ability, and the ability to differentiate into bone, cartilage, and fat. Unlike bone marrow stem cells (BMSCs) or adipose stem cells (ASCs), SCX is always expressed in TSPCs. When implanted, TSPCs form ectopic tissue. When SCX is introduced into bone marrow stem cells, these cells acquire the characteristics of TSPCs. The expression of SRY-Box transcription factor 9 (SOX9), a marker of cartilage formation, as well as osteogenic potential, is significantly reduced in these cells (6). In TSPCs, biglycan and fibromodulin proteins in the ECM of tendon maintain the microenvironment (stem cell niche) and control bone morphogenetic protein (BMP) signals to prevent osteogenic differentiation (4, 5). While TSPCs do not proliferate under normal conditions, they generate ECM under dynamic stimulation (Fig. 3) (4, 5).
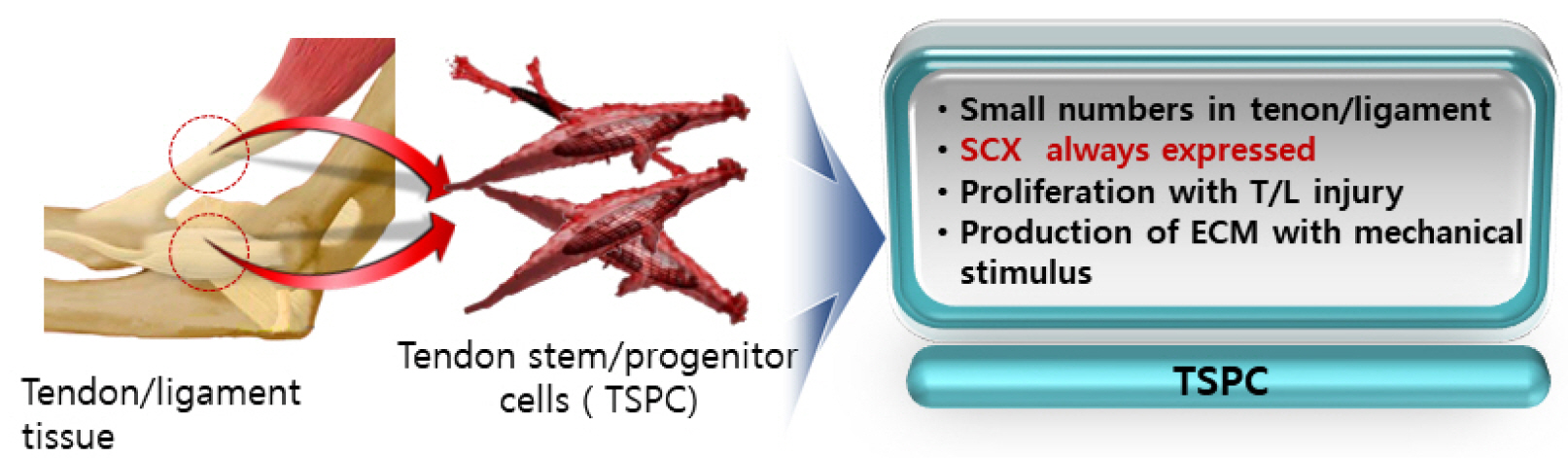
Go to :
Transcription Factors Involved in the Differentiation of Tendon
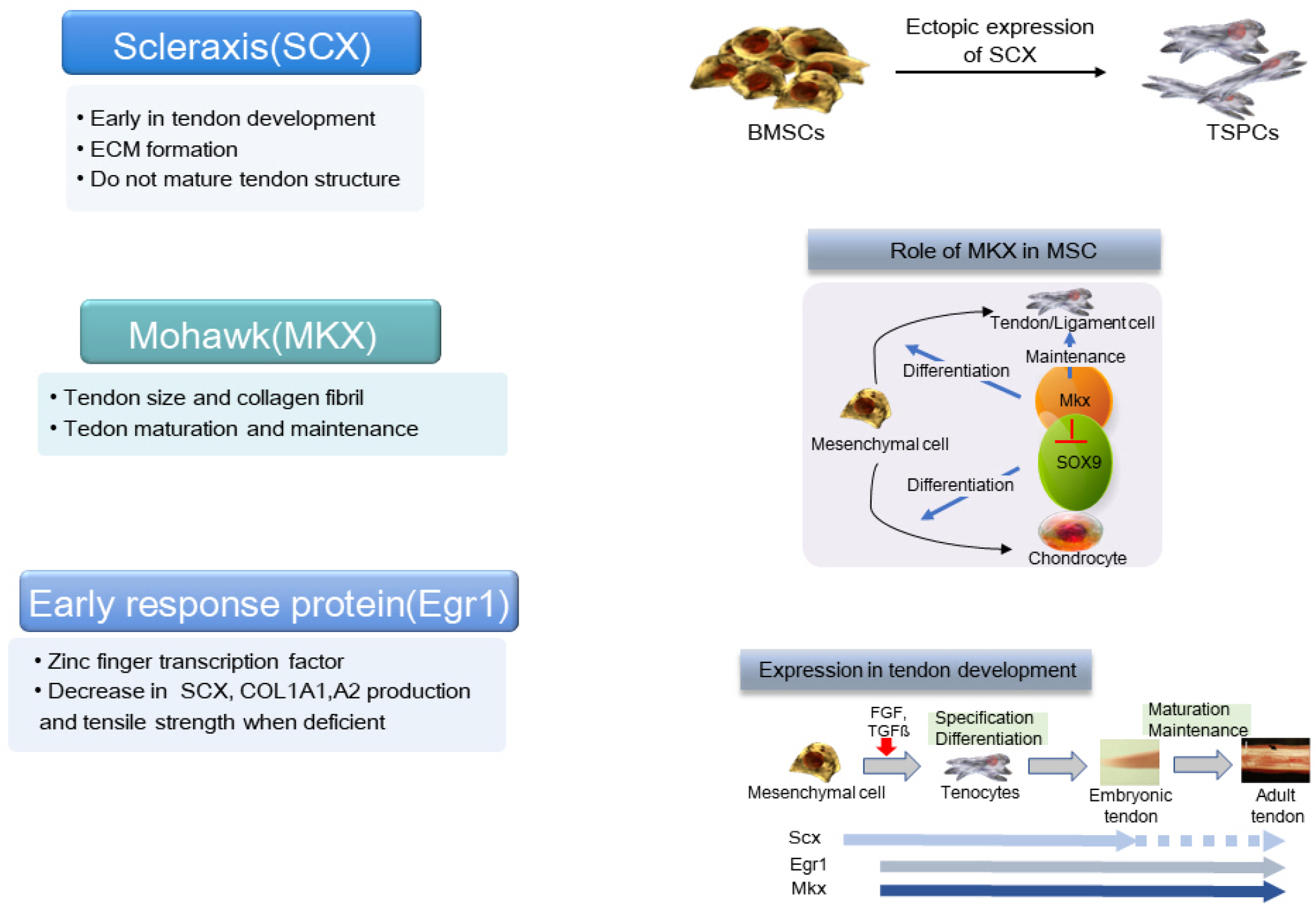
While the role of specific growth factors and transcrip-tion factors is well established in the osteogenic and chondrogenic differentiation of stem cells, surprisingly, a consensus has not been established for tenogenic differentia-tion (1, 7). The currently known transcription factors necessary for tendon development include SCX, Mohawk (MKX), and early response protein (Egr1). SCX is a helix-loop-helix transcription factor, first known as a tendon/ligament specific factor, expressed in the early stages of tendon development. It promotes the transcription of type 1 collagen a1/a2, aggrecan, and Tnmd genes. When SCX is knocked out, tendon development is suppressed due to the absent aggregation of tendon progenitor cells, resulting in the disorderly arrangement of fibrils. The development of bone tubercles to which ligaments are attached is also inhibited. The expression of SCX in ectopic embryonic stem cells initiates tenogenic differentiation. SCX increases the production of ECM but does not mature tendon structures (1, 7).
MKX also regulates the expression of type 1 collagen in tendon development. The deficient expression of MKX does not change the number of cells, but it reduces the size of the tendon and the diameter of the collagen fibers. MKX promotes the expression of SCX by binding to the transforming growth factor (TGF)-β2 promoter. MKX inhibits osteogenic, chondrogenic, and adipogenic differentiation, as well as the proliferative potential of MSCs. MKX plays a more important role in the maturation and maintenance of tendons, whereas SCX initiates tenogenic differentiation (7-9). A reduction in MKX expression was reported in the anterior cruciate ligament (ACL) of patients with tendinopathies or knee osteoarthritis. MKX also inhibits chondrogenic metaplasia of mature tendons by inhibiting SOX9 (7, 9).
Egr1 is a zinc finger transcription factor, which targets the expression of the TGF-β2 promoter. Egr1 knockout suppresses the production of SCX and type 1 collagen a1/a2, thus reducing the tensile strength of the tendon (Fig. 4) (1, 7).
Go to :
Growth Factors that Induce Tendon Proliferation and Differentiation
Growth factors that were investigated for TSPC proliferation are connective tissue growth factor (CTGF), insulin-like growth factor -1/2, platelet-derived growth factor BB (PDGF-BB), and basic fibroblast growth factor (bFGF). Connective tissue growth factor (CTGF/CCN2) increases the expression of type 1 collagen and tenascin. It also promotes the production of SCX and tenomodulin by modulating FAK and ERK1/2 signaling (10). CTGF supplementation enriched a CD146 (tendon stem cell marker)+ TSPC subpopulation, and induced proliferation and tenogenic differentiation from rat patellar tendon-derived cells (11). CTGF also recruited CD146+cells and enhanced tendon healing in vivo (11). While either PDGF-BB or bFGF significantly increased cell proliferation and matrix synthesis in tendon fibroblast culture (12, 13), co-treatment of PDGF-BB combined with bFGF dose-dependently led to an accelerated cell proliferation (13). When combinations of growth factors are utilized, unexpected interactions can be engendered with activation of different signaling pathways. Of several different combinations, IGF1 plus GDF5 demonstrated the most spectacular increase in the expression of tenogenic marker and collagen synthesis (14).
TGF-β signaling pathway is involved in inducing and maintaining tendon progenitors in the developing embryo: disturbance in either ligands or receptors resulting in loss of all tendon structure (14, 15). TGF-β ligands are potent inducers of tendon markers including SCX (14, 15). Myostatin, another TGF-β Smad2/3 subfamily member, also has similar tenogenic properties (16). While several BMPs inhibit tenogenic induction, a subset of BMPs positively regulate tendon-specific differentiation (17-19). Growth differentiation factors (GDF)-5,6,7 relatively specific growth factors for tendon differentiation, also called bone morphogenetic protein (BMP) -14,13,12 or cartilage-derived morphogenetic protein (CDMP) -1,2,3 respectively, are closely related factors belonging to the TGF-β superfamily (20-23). The treatment of MSCs with these factors was shown to promote ectopic tendon/ligament differentiation, whereas other BMPs promoted osteogenic or chondrogenic differentiation (23). GDF factors induce tenogenic differentiation through Smad8 signaling system. Without these GDF factors, the size of collagen fiber decreases, and healing is delayed in the case of tendon injury. A deficiency in one factor can be compensated by other factors. Sutures coated with GDF5/BMP14 were reported to promote collagen synthesis. When treated with GDF5/BMP14, rat TSPCs had increased SCX expression with reduced adipogenic and chondrogenic potential (24). GDF5/BMP14 treatment also increased cell proliferation and matrix content of murine tendon fibroblasts (25, 26). Collagen sponges containing GDF7/BMP12 promoted the healing of tendon damage in patients with rotator cuff injuries in clinical trials (14). Similarly, GDF7/BMP12 upregulated tendon markers SCX, Tnmd, type 1 collagen and tenascin in rat TSPCs (27). BMP12 combined with CTGF overexpression had an additive effect on tenogenic gene expressions (27). TSPCs co-transfected with BMP12 and CTGF genes had increased SCX, type 1 collagen a1, type 3 collagen a1, and tenascin expression while osteo-, chondro-, and adipogenic markers were inhibited (28).
Go to :
Mechanical and Environmental Factors for Tenogenic Differentiation
Another strategy to induce tenogenic differentiation is mechanical stimulations similar to those seen in native microenvironments (29, 30). Although individual tendons undergo specific loading subject to different anatomic locations, the general loading type for most tendons is uniaxial tension. When uniaxial and biaxial stimulations were directly compared, uniaxial loading was more strongly tenogenic to TSPCs than biaxial loading, which induced a mixed osteogenic, adipogenic, and chondrogenic signals (30). Uniaxial loading of three-dimensional TSPC significantly promoted tenogenic differentiation, neo-tendon formation, and mechanical properties of formed tendon (30).
TSPCs typically show reduced proliferation and ECM production as well as decreased tendon and stemness markers at later passages of culture (31, 32). TSPCs cultured at low oxygen levels showed enhanced cell proliferation and increased gene expression of stemness and tendon markers while osteogenic, adipogenic, and chondrogenic gene expressions were reduced, avoiding problems seen in long-term culture (33-35).
Go to :
Induction of Tenogenic Differentiation from Pluripotent Stem Cells
Tenogenic stem cells can be induced from pluripotent stem cells. Komura et al. (36) developed a protocol for inducing tenogenic differentiation from mouse induced pluripotent stem cells (miPSCs) using serial treatment of Wnt3a and activin A, basic FGF, and TGF-β1 and bFGF. They created reporter mice that express enhanced green fluorescent protein (EGFP) driven by SCX promoter and generated iPSC from these mice. The iPSC-derived EGFP-positive cells exhibited elevated expression of tendon-specific genes, including SCX, MHK, Tnmd, and FMOD, indicating that they have tenocyte-like properties.
McClellan et al. (37) differentiated equine embryonic stem cells (eESCs) to tenocytes by culturing without feeders in ESC media lacking LIF and in the presence of 20 ng/ml TGF-β3 for 14 days. Dale et al. (38) induced tenogenic differentiation of hESCs using BMP12 and BMP13 in the presence of ascorbic acid (AA). A tenocyte-like elongated morphology was observed in hESC after 40-days continuous supplementation with BMP12, BMP13 and AA.
Go to :
Current Status of Treatment for Tendon and Ligament Injuries
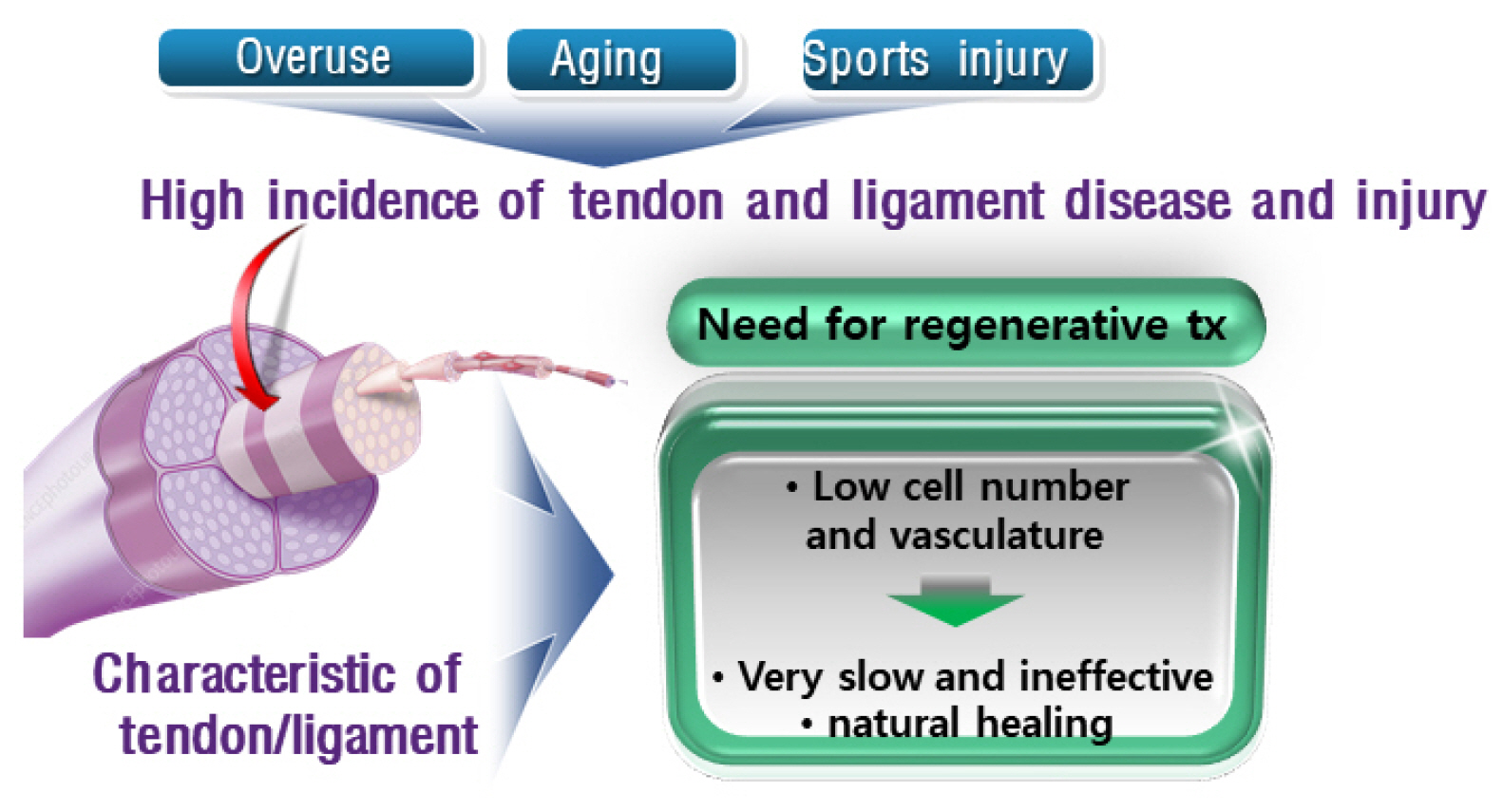
Tendon and ligament damage is very common, with about 3∼5 million cases worldwide each year, accounting for 50% of the physical injuries (39). Tendon and ligament injuries are mostly caused by degenerative changes due to overuse, aging, or sports injuries. Natural healing is very slow and inefficient due to the low vascularization of tendons and ligaments (1, 4, 5). When a tendon ruptures, it undergoes a healing process that consists of inflammatory, proliferative, matrix formation, and remodeling phases. While surgical repair is usually performed, re-rupture frequently occurs due to incomplete tendon regeneration and it is exceedingly difficult to regain pre-rupture strength.
ACL rupture is one of the most common sports injuries. It occurs frequently at 1 in 3,000 people, with 200,000 surgical procedures performed every year in the United States alone (40). ACL rupture is currently treated with reconstructive procedures using autologous or allograft grafts. Complications, such as rupture, donor site morbidity, instability, and secondary osteoarthritis, are problems after ACL rupture and repair.
Rotator cuff tears (RCTs), the most common degenerative tendon disease, occur in about 30% of people over the age of 60. Of them, 35% progress to the extent that daily life is difficult (41, 42). Around 200,000 operations are performed for RCTs each year in the United States, costing about 500 million dollars (43). Unfortunately, the re-rupture of large size RCTs after surgical repair is quite common, leading to treatment failure. Regenerative treatment has been considered or applied, including growth factors, biomaterials, platelet-rich plasma, and cell therapy (1, 5).
Go to :
Need for Regenerative Treatment for Tendon and Ligament Injuries
As a regenerative cell therapy for tendon and ligament regeneration, autologous bone marrow aspirate concentrate, autologous stromal vascular fraction or platelet-rich plasma have been applied. It should be noted that they are not specific treatments for tendon regeneration. Also, there are great differences in the method of administration depending on the treating physicians, with controversial effects.
There is currently no adequate method of treatment for extensive tendon defects seen in RCTs. A treatment is necessary that can respond to the unmet need to augment tendon suturing in wide tendon defects with additional regenerative effects. In ACL rupture, as a direct repair is mostly unsuccessful, reconstruction using autogenous or allogeneic tendon is currently the standard treatment, which is a larger operation than primary suture. Also, autografting is associated with donor site morbidity while the latter has an associated re-rupture rate. Methods are necessary that can reduce treatment morbidity by increasing the success rate of tendon suturing and reduce the re-rupture rate of allograft reconstruction. In the case of tendinopathies, a successful treatment method is needed to induce regeneration of the degenerative tendon causing discomfort and deterioration in the quality of life (Fig. 5).
There are sporadic reports on the clinical application of stem cell for tendon regeneration. Hernigou et al. (44) evaluated the efficiency of biologic augmentation of rotator cuff repair with iliac crest BMSCs. Forty-five patients in the study group received concentrated BMSCs (51,000± 25,000) as an adjunct to rotator cuff repair at the time of arthroscopy. BMSC injection during rotator cuff repair enhanced the healing rate and improved the quality of repair. At the most recent follow-up of 10 years, intact rotator cuffs were found in 39 (87%) of the 45 patients in the MSC-treated group, but just 20 (44%) of the 45 patients in the control group (44).
Jo et al. (45) assessed the result of intratendinous injection of ASCs for partial-thickness rotator cuff tear at mid-term follow-up. The first part of the study consisted of 3 dose-escalation groups, with 3 patients each: low-dose (1.0×107 cells), mid-dose (5.0×107), and high-dose (1.0× 108) groups. For the second part, 10 patients with high dose treatment were added. Intratendinous injection of ASCs reduced shoulder pain by approximately 90% at 1 and 2 years in the mid- and high-dose groups. The strength of rotator cuff muscles significantly increased by greater than 50% at 2 years in the high-dose group. Magnetic resonance imaging showed that the volume of tendon defects in the high-dose group nearly disappeared at 1 year and did not recur at up to 2 years (45).
Go to :
Future Strategy: Generation of Tendon and Ligament-Specific Stem Cells by Gene Transfer of Tendon-Specific Transcription Factor
Considering that the number of TSPS available from tendon harvest is limited, a viable strategy would be to derive therapeutic tenogenic stem cells from BMSCs or ASCs. While it is possible to induce MSCs into tenogenic stem cells by incubating them with growth factors such as GDFs, the in vitro culture time is longer, and the cost of serum and growth factors required for culture is increased accordingly. In comparison, if tendon-differentiated transcription factor genes can be transferred to MSCs with high efficiency, these MSCs are expected to have the properties of tenogenic stem cells. It has already been confirmed that BMSCs and ASCs, which have SCX and MHK genes transferred using adenovirus, have properties similar to TSPCs (1, 6, 8). MSCs to which the gene for differentiation transcription factor has been transferred can be used as cell therapy for tendon and ligament regeneration without separation or additional culture. Therefore, it is expected that the gene transfer of tendon-specific transcription factors will be effective in generating tendon and ligament-specific stem cells. Use of nonviral vectors or adeno-associated viral vector should be considered because of nonlethal nature of the tendon or ligament disease. Combining the application of these tenogenic stem cells with current surgical repair techniques may provide a less expensive and technically feasible regenerative technology for tendon and ligament injuries resistant to current treatment.
Go to :
 XML Download
XML Download