

 PDF
PDF Citation
Citation Print
Print


Background
Stem cells, an emerging tool for regenerative medicine, are disparate from other cell types of our body owing to exclusive properties they embrace. Being unspecialized, these cells retain self-renewal capacity, hence regenerate their pool by cell division; symmetric or asymmetric which further leads to differentiation into diverse cell types. Based on the source of isolation, stem cells can be embryonic, fetal, or adult which can be cord blood, placental, bone marrow (BM), dental, adipose tissue, skin, etc. (1).
Among all the stem cells, hematopoietic stem cells (HSCs), responsible for hematopoiesis in the adult body are the utmost explored and researched one, owing to their enormous potential as well as therapeutic applications. Apart from bone marrow, which is their reservoir, HSCs can also be isolated from granulocyte-colony stimulating factor (G-CSF) mobilized peripheral blood. Also, it is feasible to obtain HSCs and various progenitors of the blood cells from fetal as well as embryonic tissues. Moreover, the umbilical cord blood (UCB) and placenta, generally discarded after childbirth, are rich sources of HSCs (2). Somatic cells can also become the source of HSCs by exploiting Induced pluripotent stem cells (IPSCs) (3). HSCs can presumptively characterize and isolated with the help of an anticipated set of clusters of differentiation antigens and cell surface marker present on their cell membrane as predicted and proven by Irving L. Weismann in 1988. These markers change consequently as HSCs get mature and differentiate into a specific set of blood lineages. Human HSCs are identified as Cluster of Differentiation (CD) 34+, CD38low/−, c-Kit−/low, CD59+, CD90+, Lin− whereas mouse HSCs are recognized as CD34low/−, CD38+, c-Kit+, SCA-1+, CD90+/low, Lin− (4).
Carrying the vital capacity to regenerate all blood cell types through making lineage-specific progenitor cells, HSCs are widely used to treat many human disorders (5). Bone marrow transplantations or Hematopoietic stem cell transplantation are HSCs based lifesaving procedures broadly used to treat disease ranging from cancer to autoimmune diseases. Subsequently, potential therapeutics of HSCs is increasing profoundly to life-threatening diseases such as autoimmune disease, leukemia, lymphoma, rheumatoid arthritis, multiple myeloma, hemophilia, diabetes and multiple sclerosis (6-8). Numerous clinical trials have been completed and still ongoing to observe the efficacy of HSCs in treating above mentioned disorders in which either whole BM, progenitors, or purified hematopoietic stem cells have been transplanted in patients and clinical improvement has been observed. However, to the best of our knowledge, none of the clinical studies has used cultured stem cells owing to ethical issues, risk of contamination, knowledge, and availability of suitable media besides culture conditions, etc.
The dynamic implication of HSCs for therapeutics instigates concern to combat the enormous demand of HSCs. Furthermore, whether we are investigating stem cell for fundamental research purpose markedly testing the toxicity of compounds, cancer research, vaccine production, drug screening, and discovery or for future therapeutics, a prerequisite is to culture stem cells under in-vitro conditions and mimicking similar environment as existing in the individual body from where these cells are isolated (9). Moreover, the accomplishment of a massive supply of HSCs for therapeutic applications demands cells to be expanded, self-renewed, and differentiated under in vitro conditions therefore, the robust HSCs production system is indispensable. However, it is a challenge to culture stem cells as these cells were usually lying quiescent inside their source for a prolonged period and to initiate their cell division with an anticipated output of self-renewal or differentiation in specific lineages is quite cha-llenging. Several factors alike stem cell source, cell count, purification levels like mononuclear fractions, progenitor cells, or highly purified stem cells based on cell surface markers govern the fate of cells in culture.
Stem cell culturing encompasses optimized protocols as well as good quality reagents; however, the most crucial requirement is media which supplies essential components like glucose, vitamins, amino acids, etc., which are vital constituents for the metabolism of stem cells to sustain growth and proliferation (10). Media for stem cell culture not only able to provide elementary nutrients to these cells, but it should also be suited for culture conditions. Pertaining to a fine balance between stem cell self-renewal and lineage commitment, stem cell expansion, as well as differentiation under the in vitro environment, demands a highly controlled system where media along its supplements play a critical and fate-deciding role. Hence, the need for standardization and consistency in stem cell culture medium quality is crucial. Along with media oxygen concentration, cytokine combinations also influence the self-renewal and differentiation potential of HSC’s culture under the in vitro condition (11). Various scientists and companies have optimized specialized stem cell culture media and different supplements for specific applications. Here in this review, we have summarized the different types of media developed and used for HSCs culture, self-renewal, expansion, and differentiation into different cell lineages that can be used for either research purpose or therapeutics.
Go to : 
HSCs Culture
Large numbers of basal media are accessible to culture stem cells, primarily serum-free, although we can include or exclude serum as well as other growth factors based on the requirement. The quality as well as quantity of the serum used in stem cell culture media play a significant role and affect the outputs. Moreover, growth and differentia-tion factors which are generally recombinant proteins such as Interleukin-3 (IL-3), stem cells factor (SCF), thrombopoietin (TPO), granulocyte-macrophage colony-stimulating factors and erythropoietin, also regulate the survival and self-renewal of HSCs along with proliferation and differentiation (12). These growth factors have an enormous impact on HSCs and their purity, excellence and concentration decide the fate of cultured HSCs (13). Studies of the above-said cytokines used in HSCs culture under in vitro conditions shown to have a crucial role as under in vivo condition HSCs are maintained by the SCFs by its c-kit receptor and by the synergic activity of other growth factors which are released by supporting niche cells (14, 15). HSCs also requires TPO for its survival, self-renewal, and platelet production during differentiating into megakaryocytes (16). TPO acts via its myeloproliferative leukemia receptor (17). Numerous types of media and supplements have been exploited in various HSCs studies to maintain self-renewal, proliferation, and culture HSCs under in vitro conditions as discussed.
DMEM is one of the widely used modified versions of Basal Media Eagle’s (BME), which comprise of four-fold higher concentrations of vitamins, amino acids, and other supplements than BME. The original formulation of DMEM used 1000 mg/L glucose, which is predominantly utilized to culture mouse embryonic cells; various modifications of this medium is present nowadays based on the combination of glucose, L-glutamine and sodium pyruvate. Scientists utilized Dulbecco’s Modified Eagle’s Medium (DMEM) supplemented with valproic acid and 10% Fetal Bovine Serum (FBS) to culture CD133+lin−CD45− hematopoietic cells isolated from umbilical cord blood. In an another study, DMEM supplemented with FBS (15%), hIL-6 (10 ng/ml), mSCF (100 ng/ml) and mIL-3 (6 ng/ml) has been utilized to culture mouse bone marrow cells (18).
Roswell Park Memorial Institute (RPMI) 1640 media primarily developed by Moore et al., at Roswell Park Memorial Institute, uses a bicarbonate buffering system besides glutathione as a reducing agent. RPMI-1640 has been exploited to culture and increase the yield of Peripheral Blood (PB) mononuclear cells for 5∼7 days. The media was supplemented with L-glutamine (200 mmol), 2-beta mercaptoethanol (50-mmol), gentamicin (20 μg/ml), FBS (10%) and growth factors like GM-CSF, IL-4, TNF-alpha, IL-1 alpha, IL-1 beta, c-kit ligand, IL-3, M-CSF, IL-2 (19). Iscove’s Modified Dulbecco’s Medium (IMDM) media, as depicted by Iscove and Guibert, consist of sodium bicarbonate buffered systems besides numerous supplements like albumin, additional amino acids, vitamins, lecithin, selenium and transferrin (20) is another signifi-cant HSCs culture media (21). IMDM media along with various combinations of supplements like IL-3 (5 ng/ml), EPO (3 IU/ml), SCF (100 IU/ml), hydrocortisone (10 μM), insulin (330 μg/ml), L-glutamine (2 mM) and human serum has been used in the culturing and the proliferation of CD 45+ cells (22). Along with IMDM, lymphocyte growth media have also been used for the culturing of HSCs along with IMDM media (23).
Although the above discussed basal media are exten-sively used by numerous stem cell researchers globally, still, precision in stem cell culture requirement has increased tremendously for the last two decades, henceforth necessity of specialized stem cell culture media increases. Apart from the above-mentioned basal media, various specialized media are also currently employed for the HSCs culture, self-renewal, and differentiation. Serum, mostly an animal product utilized along with basal media may pose hazards while culturing human stem cells, especially for therapeutic purposes urges serum-free media utilization. Stem pro-34 SFM one of the specialized serum-free media which is comprised of stem pro basal liquid media along with frozen stem pro-nutrient supplements and is exten-sively used to support BM, PB, or Cord Blood (CB) hematopoietic stem cells culture, expansion and differentiation (24). Numerous scientific studies have been published where this media has been used to culture hematopoietic stem cells. In one of the study, the CD34−/lowc-Kit+Sca-1+ lineage bone marrow cells have been cultured using StemPro-34 SFM supplemented with 0.05 mM 2-β-mer-captoethanol, 2 mM L-glutamine and cytokines cocktail of mouse SCF (10 ng/ml), mouse IL-3 (10 ng/ml), human TPO (100 ng/ml), human G-CSF (10 ng/ml), human IL-11 (100 ng/ml), and human IL-6 (100 ng/ml) till 14 days. Different cytokine cocktail possessed different efficiency in pertaining self-renewal of stem cells, SCF alone increases 25% of self-renewal but for a limited period of 2 weeks, While TPO alone increases the potential fewer than 10-5%. But the combination of TPO and SCF induced around 90% potential of cell division. IL-3, IL-6, and G-CSF in combination with SCF significantly increased the potential of cell division to 50∼60%, while alone did not prompt any cell division (25).
Go to :
HSCs Expansion
A substantial amount of hematopoietic stem cells is imperative for both stem cell researches as well as for therapeutics and can be acquired through in vitro stem cell expansion which is a long-term culture of HSCs without their exhaustion or differentiation which may happen during HSC culturing. Appropriate culture conditions and expansion media with necessary supplements are prere-quisites for this process of continuous maintenance of HSC’s features and CD markers. A variety of media has been used for various experimental studies for the expansion of HSCs as described in Table 1 (26-31), many of these are modified basal media and are available commercially too. Stem SpanTM Serum Free Expansion Media (SFEM) is a defined, serum-free medium consisting of IMDM, bovine serum albumin, rHuman insulin, human transferrin, 2-β-mercaptoethanol, and supplements. In one of the study 1×105 lin− cells/ml isolated from UCB have been cultured in SFEM with SCF (100 ng/ml), of Flt-3L (100 ng/ml), TPO (50 ng/ml) and low-density lipoproteins (1 μg/ml) for 8 days in the primary culture bag. The media was changed every four days and the resultant culture conditions lead to 3.3-fold expansion of HSCs (32). In another study, StemSpan medium supplemented with cytokine cocktail of SCF (100 ng/ml), TPO (100 ng/ml), Flt-3L (100 ng/ml), resveratrol (10 mM) and IL-6 (100 ng/ml) was utilized to expand HSCs which were later transplanted in mice model. Resveratrol maintained the HSCs phenotype, enhanced the expansion and engraftment potential of cultured HSCs (33). Likewise, SFEM II which is a modified version of SFEM also utilized to expand HSCs (26). Stem Pro SFM, a widely used media for HSCs culturing purposes and seems valuable for HSCs expansion. Stem pro-34 SFM along with FBS (10%), L-glutamine (2 mM), TPO (50 ng/ml), rhIL-3 (200 ng/ml) and rhFlt-3L (200 ng/ml) has been utilized for the expansion of UCB cells (27). In the experimentation, IL-3 enhanced the proliferation and expansion of HSCs (18). In another study Stem pro media was used to proliferate erythroid precur-sor cells (34).
Table 1
List of different culture media along with supplements for expansion of stem cells from various sources
| Media | Cell source | Cytokines combinations and supplements |
Phenotypic markers |
No. of cells | Passage day | Fold expansion | Ref. |
|---|---|---|---|---|---|---|---|
| SFEMa | UCBb | 100 U/ml pen/strep+ 2 mM L-glut+ 25 ng/ml SCFc + 25 ng/ml ILd-6 +25 ng/ml TPOe + 25 ng/ml FLTf ligand | CDg 34+/CD 38+ | 1.0×104/well | 5 days | ND | (26) |
| Stem Pro 34SFMh | UCBb | 10% FBSi+100 ng/mlPen/Strep+ 2 mM L-glutamine +hILd-3 peptide 8+50 ng/ml TPOe+ 200 ng/ml rhFLTf-3 | CD 34+ | 7.3×104 | 7 days | 19-fold | (27) |
| XVIVO 10 | PBMCj | 1% human serum albumin+ 50 ng/ml SCFc + 30 ng/ml FLTf3-ligand+ TPOe + 5-50 ng/ml human ILd -3 | CD 34+ | 2×104 | 2 to 4 days | 2.5-fold | (28) |
| IMDMk | UCBb | 20% FBSi and 5 ng/ml Sel + 50 ng/ml TPOe + 25 μg/ml insulin + 200 μg/ml transferrin, 100 ng/ml SCFc + 100 ng/ml FLTf+ 10 μg/mL FAm | CD 34+ | 1×105 | 5 days | 20-fold | (29) |
| SFEMa | BMn | 10% FBSi+ 1% pen/strep +100 ng/ml FLTf-3 + 100 ng/ml TPOe+ 100 ng/ml SCFc | CD 34+ | 1×105 | 24 hours | ND | (30) |
| HPGM° 1640 | PBMCj | 10 FBSi +100 U/ml Pen/Strep + 100 ng/ml SCFc + 20 ng/ml ILd-3+30 ng/ml | CD 34+ | 2.5×105 | Up to 30 days | 8-fold | (31) |
aSerum free expansion medium, bUmbilical cord blood, cStem cell factor, dInterleukin, eThrombopoietin, fFMS like tyrosine kinase, gCluster of differentiation, hSerum free media, iFetal bovine serum, jPeripheral blood mononuclear cells, kIscove’s modified dulbecco’s medium, lSelenium, mFolic acid, nBone marrow, °Hematopoietic growth media.
![]()
Hematopoietic Progenitor Growth Media (HPGM) is another specialized expansion media, which is xeno-free media, includes albumin, recombinant insulin, and pasteurized transferrin. HPGM is well suited for the expansion, growth, and differentiation of HSCs as well as hematopoietic progenitor cells from various sources (35). In one of the studies done by Kishimoto et al (36), HPGM media supplemented with 1× antibiotics/antimycotic solution, FBS (4%) and cytokine cocktail of TPO (20 ng/ml), Flt-3L (20 ng/ml) and SCF (10 ng/ml) with has been used for the culturing of 1×105 BM cells of the mouse/12 well plate, coated with low molecular weight heparin/protamine microparticles. The cells were cultured for 8 days under the above-said culture condition, and after 8 days there were 11-fold expansions of CD 34+ stem cells. The coated plates enhanced the expansion of CD 34+ stem cells by controlling the release of cytokines.
An auxiliary specialized media for HSCs expansion under in vitro condition is Stemline II Hematopoietic stem cell expansion media. This media is a modified version of Stemline media; hence it is a second-generation media that is generated to increase cell expansion besides to optimize the balance of differentiated and undifferentiated cells (37). Although human serum albumin is present, this is serum-free and xeno-free media with no animal-derived constituents. Ficoll sorted BM and PB CD34+CD38− cells from CML patients have been expanded using Stemline II media, supplemented with cytokine cocktail of TPO (20 ng/ml), SCF (100 ng/ml) and Flt-3L (100 ng/ml) in absence of retinoic acid. The absence of retinoic acid in the culture condition leads to an increase in the expansion of HSC’s and its self -renewal ability (38). In a yet another study the Stemline II media supplemented with human SCF (150 ng/ml), Flt-3L (100 ng/ml), IL-3 (50 ng/ml), TPO (100 ng/ml) and valproic acid (1 mM) was used to expand the CD34+CD38− cells for 7 days. Along with cytokine cocktail, valproic acid via inhibiting the histone deacetylase activity, shown to have a prominent role in HSCs cell survival and expansion (39).
X-VIVO hematopoietic cell medium is a specialized Serum-free chemically defined media, devoid of any exogenous growth factors, undefined or artificial stimulators, provides a balanced and complete environment for culturing HSCs (40). In one of the stem cell expansion studies, CD34+ PB mononuclear cells have been cultured to expand into X-VIVO 10 media supplemented with human SCF (50 ng/ml), 1% serum albumin and cytokine cocktail of human IL-3, Flt-3L, and TPO (28).
Go to :
HSCs Differentiation
Each type of stem cell requires an exclusive culture medium supplemented with specific growth factors to sustain cell proliferation and differentiation into precise lineages. HSCs differentiate in all mature blood cell lineages and this differentiation process is earnestly regulated by intracellular signaling pathways which in turn governed by growth factors and cytokines. Various scientific studies involving in vitro HSCs differentiation into particular lineages, blood, or other tissue types, indicate the use of numerous basal as well as specialized media along with several supplements including cytokines and growth factors cocktails under stringent culture conditions as depicted in Fig. 1 (19, 41-43).
 | Fig. 1HSCs differentiation into various lineages. (a) Neuronal differentiation using Methylcellulose (MCM) and α-MEM with Inter-leukin-3(IL-3), β-Mercaptoethanol (BM), 1% bovine serum albumin (BSA), sodium L-glutamine (Na L-G), human recombinant SCF, GM-CSF, 20% FBS, Gentamicin (Gent) and Retinoic acid (42). (b) Red Blood Cells (RBCs) differentiation using IMDM with L-Glutamine (L-G), L-monothioglycerol (L-MT), SCF., 1% Pen/Strep, 1% Human serum albumin (HAS), Ferric nitrate (FN), Transferrin (T), Hydrocortisone (HC), IL-3, Erythropoietin (EPO), Insulin (I) and BM-MSCs (43). (c) Endothelial Progenitor cells differentiation using IMDM and essential basal medium (EBM), SCF+, Flt-3 L, Thrombopoietin (TPO), IL-3, GM-CSF, Vascular endothelial growth factor (VEGF), Ascorbic acid (AA), heparin, Hydrocortisone (HC), IGF, EGF, Ferric nitrate (FN), L-GlutamineL-G and FGF (41). (d) Dendritic cells differentiation using RPMI-1640, L-GlutamineL-G, β-Mercaptoethanol (BM), Gentamicin (Gent), human recombinant (rh)-IL-4, rh-GM-CSF, rh-Tumor necrosis factor alpha (TNF-a) rh-IL-lα, IL-1β, c-kit, rh-IL-3, rh-M-CSF, rh-IL-2 (19).
|
In one of the granulocyte-monocyte to dendritic cells differentiation study, BM cells were cultured in RPMI-1640, supplemented with 1× streptomycin/penicillin, FBS (10%) and GM-CSF (20 ng/ml) for 6 days. After 6 days the induced cells were cultured in IMDM supplemented with 1x penicillin/streptomycin, 10% FBS, Flt-3L 200 ng/ml, mercaptoethanol (50 μM) and sodium pyruvate (1 mM) (44). RPMI1640 is also used for the differentiation of HSCs into insulin-producing cells (45). IMDM has already been used for a variety of cells like hematopoietic stem progenitor cells from bone marrow, B-lymphocytes, T lymphocytes, B cells stimulated with lipopolysaccharide, etc. In another study where an attempt has been made to differentiate CB CD 34+ cells into endothelial progenitors cells, for proliferation phase, 2×105 CD 34+ cord blood cells/ml were placed in IMDM containing cytokines cocktail of IL-3 (10 ng/ml), SCF (200 ng/ml), Vascular Endothelial Growth Factor (VEGF) (50 ng/ml), Flt-3 L (200 ng/ml), TPO (20 ng/ml), and GM-CSF (12.5 ng/ml), seeded into the 24-well plates and the cells were sub-cultured when the cell density reached 1×106 cells/ml. These cells were further expanded and differentiated into endothelial progenitor cells (41). Moreover, using IMDM media megakaryocytes have been generated from mobilized PB and hematopoietic progenitor cells from BM (46).
X-VIVO 10, a widely HSCs expansion media can also be utilized by expanding cells for differentiation purposes. In a study conducted by Stec et al (47), X-VIVO 10 media supplemented with 4% of fetal calf serum (FCS), TPO (15 ng/ml), Flt-3L (30 ng/ml), SCF (50 ng/ml), IL-3 (30 ng/ml) was used as the first step for the expansion of CB hematopoietic CD34+ mononuclear cells. Every 3 to 5 days the cells were sub-cultured and enriched with fresh media. These expanded cells were later differentiated into monocytes by using IMDM media supplemented with SCF (25 ng/ml), IL-3 (30 ng/ml), 20% FCS, monocyte colony-stimulating factor (30 ng/ml), Flt-3L (30 ng/ml) for 3∼14 days.
Methylcellulose is also one of the varieties of diverse media available for in vitro culturing of HSCs and progenitor cells. Methylcellulose is used to enumerate the mature differentiated blood lineage generating capacity of HSCs, which is known as Colony-forming cell assay which generates colony-forming erythroid, myeloid, and mixed lineage progenitors (48). This media provides diverse progeny on different regions of the plate and thus, it is easy to distinguish the mixture of progenitors generated from the HSCs. Methocult is the variant of methylcellulose media used in several stem cell studies (49). Human methylcellu-lose enriched media is also a modification of methylcellu-lose and is used to distinguish the mixed cell population into respective progenitors. For the assay of erythroid, myeloid, and mixed lineage progenitors from a BM isolated mixed lin− sample, 0.8% methylcellulose in the alpha medium was used in a study done by Lemieux M et al. The cells were cultured along with the various supplemented such as 1% bovine serum albumin, 30% FCS, 0.1 mmol/L β mercaptoethanol, 2% pokeweed mitogen-stimulated mouse spleen cell-conditioned media and 3 U/ml human erythropoietin. The cells were assayed after 14 days (50).
Go to :
Conclusions
Stem cells are an emerging therapeutic tool to cure numerous life-threatening human ailments. With escalating research and clinical use, rises the need for a large amount of stem cells, which can significantly fulfill through stem cell culture under in vitro conditions in which culture media plays an imperative role. Stem cell culture media has diverse applications and utilities ranging from self-renewal, proliferation, expansion, and differentiation. For reproducible results in stem cell experiments meant for research purposes as well as therapeutics, one must use quality controlled standard media which will enhance the validity and consistency in this highly desirable field. Media has been formalized from time to time as per the need for stem cells to deliver an improved in vitro environment for stem cell development. Numerous scientists in addition to biological industries are continuously moving forward in this direction and formulating improved as well as ready to use appropriate stem cell culture media. It is mandatory to observe stem cell behavior and culture condition requirements while culturing cells either in basal media with various supplements or commercially available specialized medium.
Here in this review we have summarized diverse kinds of media available and used by several scientific groups for in vitro culturing HSCs. We have cited most of the media available and used in culturing hematopoietic stem cells along with the commercial suppliers as given in Table 2. As per the analysis of various HSCs culturing media in this review article, depicted in Fig. 2 it has been noted that IMDM and RPMI are extensively exploited basal media, also X-VIVO, SFEM are the foremost used serum-free specialized media. Keeping in mind the experiments and applications of HSCs culture, stem cell researchers can choose from the conferred media in this article. However, stem cell scientific community need more derived media for the efficient culturing of HSCs, so that researchers can work on stem cell field proficiently and exploit the field extensively.
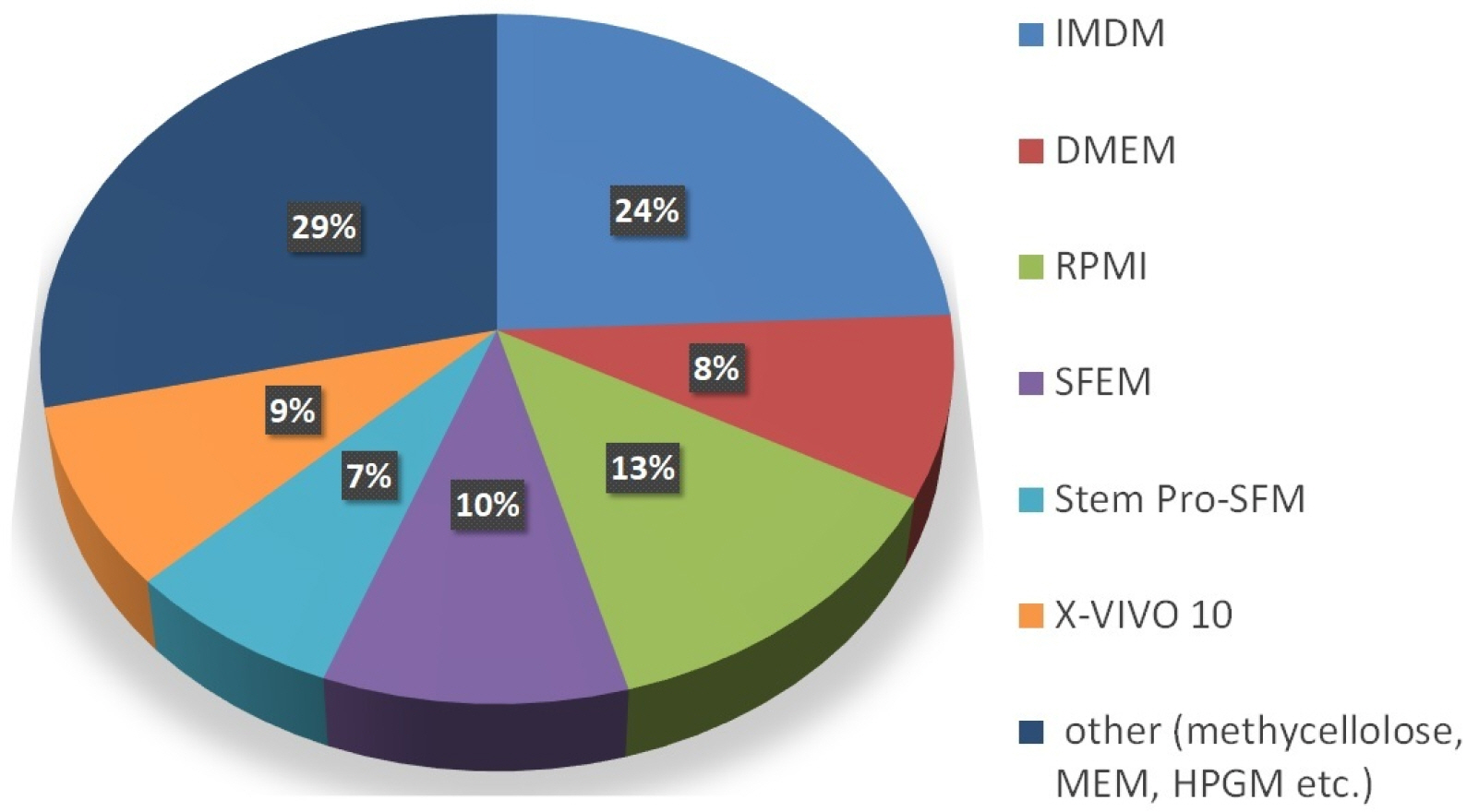 | Fig. 2Pie chart depicting percentage distribution: various culture media for HSC proliferation, expansion and differentiation based on hematopoietic research papers included in this review.
|
Table 2
Different stem cell media and their commercial suppliers
| Media | Name of the suppliers/company |
|---|---|
| AIM V (19) | Invitrogen |
| DMEMa (21) | Lonza, Thermofischer, Hyclone, Biochrom, Sigma-Aldrich, Cellgro, GIBCO Invitrogen, STEMCELL Technologies, BioWhittaker, PAN Biotech etc. |
| LGMc-3 (23) | Clonetics Corp., Walkersville |
| StemPro CDd 34+ (25) | Thermofischer, Gibco |
| Stem X-VIVO (28) | Lonza |
| IMDMb (29) | Thermo Fischer, Lonza, Himedia, Gibco, ATCC, Life Technologies |
| Stemspan SFEMe and SFEM II (30) | STEMCELL Technologies |
| HPGMf (31) | Lonza |
| Stemline and Stemline II (38) | Sigma |
| Alpha-MEMg (41) | American Type Culture Collection, BioWhittaker, Gemini Bio-Products, HaartBio Ltd, Gibco, Lonza, Mediatech, Irvine Scientific and Sigma etc. |
| RPMIh (45) | GIBCO Invitrogen, HiMedia, Biochrom, BioWhitaker, Cambrex, STEMCELL Technologies, Cellgro, Biowest, CRUK, HyClone, ATCC, Irvine, Lonza, Gibco etc. |
![]()
Go to :
 XML Download
XML Download