

 PDF
PDF Citation
Citation Print
Print


Introduction
The liver is the main metabolic and detoxifying organ of the body. Liver diseases occur due to multifactorial causes including viral infection, excessive alcohol consumption, metabolic malfunction associated with obesity, hereditary genetic disorder, and drug-induced liver injury, which leads to approximately two million deaths per year globally (1). Therefore, to understand the etiology of diseases and develop therapies, various liver models have been established (2, 3). Specifically, human cell-based liver models are required, because of interspecies differences with animal models despite their merits as in vivo platforms (4). Primary human hepatocytes (PHHs) have been considered the gold standard model for hepatotoxicity prediction and drug evaluation owing to their mature functionality. However, useful human liver cell sources are still urgently needed due to the low availability and difficulty in long-term functional maintenance of PHHs in culture.
Recently, stem cell technologies have been proposed as novel methods for obtaining human hepatic cells; such technologies include the following: 1) acquisition of expandable hepatic cells from somatic cells by genetic (5) and small molecules-mediated-(6) reprogramming approaches, 2) hepatic differentiation from pluripotent stem cells (PSCs) (7-9), and 3) three-dimensional (3D) organoid generation (10-13). Organoids are 3D stem cell-derived-miniature tissues recapitulating the structure and functions of native organs (14). Liver organoids have been developed using various approaches (15) whereby hepatic cells derived from liver tissue (16, 17) or PSCs (18-20) were cultured in a 3D extracellular matrix such as Matrigel. We also generated PSC-derived expandable 3D human hepatic organoids (21).
Organoid generation is a spatiotemporal niche-reproducing process that follows developmental stages (22). Organoids derived from stepwise differentiation of PSCs usually represent immature structural phenotypes and functions (23). Further maturation was enhanced in human intestinal organoids (24) and in liver organoids (18) after in vivo transplantation. Alternatively, long-term in vitro prolonged culture of organoids resulted in functional maturation with diverse cell compositions in PSC-derived human brain organoids (25). Therefore, we performed long-term culture of hepatic organoids and optimized long-term expansion and differentiation methods.
Materials and Methods
Hepatic organoids generation
Human induced pluripotent stem cells (hiPSCs) generated from human foreskin fibroblasts (CRL-2097, the American Type Culture Collection), using a CytoTuneⓇ-iPS 2.0 Sendai Reprogramming Kit (Thermo Fisher; A16517), were routinely maintained on a γ-irradiated mouse embryonic fibroblast feeder in iPSCs culture medium (21) at 37℃, 5% CO2. To assess hepatic differentiation, hiPSCs were seeded onto MatrigelTM (Corning; 354234)-coated dishes supplemented with PSC medium and cultured for 2∼3 days until cells reached 90% confluence. The medium was exchanged with RPMI 1640 (Thermo Fisher; 11875093) based 1×B-27 supplement, minus insulin (Thermo Fisher; A1895601) and 100 ng/mL recombinant human activin A (PeproTech; 120-14e). Cells were further incubated for six days to differentiate into definitive endoderm (DE). Cells were differentiated into hepatic endoderm (HE) by treatment with RPMI 1640 based 1×B27 supplement (Thermo Fisher; 17504044), 10 ng/mL basic fibroblast growth factor (bFGF) (PeproTech; 100-18B), and 20 ng/mL recombinant human bone morphogenetic protein (BMP)4 (PeproTech; 120-05ET) under 5% hypoxia for four days. For hepatic maturation, the medium was replaced with Hepatocyte Culture Medium (Lonza; CC-3198) without epidermal growth factor (EGF), mixed with Endothelial Cell Growth Medium-2 (Lonza; CC-3162) in a 1:1 ratio, supplemented with 2.5% fetal bovine serum (RMBIO; FBS-BBT-5XM), 100 nM dexamethasone (Sigma-Aldrich; D4902), 20 ng/mL recombinant human Oncostatin M (R&D system; 295-OM-050), and 10 ng/mL recombinant human hepatocyte growth factor (HGF) (PeproTech; 100-39) for four days under 5% hypoxia, and subsequently under normoxic conditions for a further eight days or more. Arpproximately 25 days after seeding, cyst-shaped 3D organoids were spontaneously generated from 2D monolayers of mature hepatocytes. The organoids, including a few free-floating organoids, were collected and embedded in Matrigel supplemented with Hepatic Medium (HM). These conditions were previously optimized for maintaining functional hepatic organoids (Supplementary Table S1) (21).
Immunostaining
The organoids were washed with phosphate-buffered saline (PBS), fixed with 4% paraformaldehyde in PBS for 15 minutes, and permeabilized with 0.25% Triton X-100 in PBS for 15 minutes at room temperature (RT). The organoids were incubated with 4% bovine serum albumin in PBS for one hour at RT for blocking and then stained with primary antibodies (Supplementary Table S2) diluted in blocking buffer at 4℃ overnight. The samples were washed three times with 0.05% Tween-20 (Sigma-Aldrich; P9416) in PBS and then stained with Alexa FluorⓇ conjugated secondary antibodies for 40 minutes at RT. The nuclei were stained with DAPI reagent (Sigma-Aldrich; D5942). Florescence images were obtained with an Olympus microscope.
Real-time polymerase chain reaction (PCR)
Total RNA was purified using by Trizol reagent (Thermo Fisher; 15596018) according to the manufacturer’s instructions. Reverse transcription was performed with a TOPScriptTM RT DryMIX (Ezynomics; RT200). Quantitative real-time PCR was performed using Fast SYBRⓇ Green Master Mix (Applied Biosystems; 4385614) with gene-specific primers (Supplementary Table S3) in a 7500 Fast Real-Time PCR System (Applied Biosystems). β-ACTIN was used as an internal control.
Long term expansion of hepatic organoids with passaging
Organoids were routinely cultured under HM medium, which was replenished every 2∼3 days depending on the culture density. The organoids were mechanically split every seven days; the Matrigel was removed with cold PBS and the organoids were cut into 200∼250 μm sections using a sterile surgical blade under a dissecting microscope and resuspended in fresh Matrigel at a 1:3∼1:5 ratio. After solidification of Matrigel for at least 5 minutes, HM medium was gently added.
Hepatic organoids freezing and thawing
To prepare for cryopreservation, the small sections of the split organoids were mixed with mFreSRTM (Stem Cell Technology; 05855), and freezing/thawing was carefully performed by standard procedures. After thawing, HM medium containing 10 μM Y-27632 (Tocris; 1254), a ROCK inhibitor, was used for three days. To determine the viability of the organoids, cells were dissociated into single cells using TrypLE Express (Thermo Fisher Scientific; 12605010) at 37℃ for 5∼10 minutes and stained with trypan blue. Live and dead cells were counted by Countess II Automated Cell Counter (Thermo fisher; AMQAX1000).
Karyotyping
The organoids were dissociated into single cells and cultured in Matrigel-coated T25 flasks containing DMEM/F-12 supplemented with 50 ng/mL EGF and 10 ng/mL bFGF for 3∼4 days. A chromosomal GTG banding karyotype analysis was performed at 550 resolution by GenDix, Inc. (Seoul, Korea).
Hepatic differentiation of the organoids
For further hepatic differentiation, the organoids maintained on HM were cultured in expansion medium (EM) (21) supplemented with 20 ng/mL recombinant BMP7 (PeproTech; 120-03) for 2∼3 days until the organoids were enlarged. The medium was replaced with differentiation medium (DM) (21) and the organoids were incubated for an additional six days. The medium was replaced every two days.
Results and Discussion
Generation of hiPSCs-derived hepatic organoids
As we previously descried (21), 3D hepatic organoids were generated from hiPSCs through the stepwise hepatic differentiation process (Fig. 1A). hiPSCs of 90% confluence on Matrigel-coated dishes at 2∼3 days after seeding were differentiated into definitive endoderm (DE) by treatment with Activin A for six days. The monolayer of cells with a cobblestone/petal-like prototypical morphology of DE were differentiated into hepatic endoderm (HE) by treatment with bFGF and BMP4 under hypoxia for four days (Fig. 1A and 1B). For hepatic maturation, cells in HE stage were differentiated into immature hepatocytes (IH) and mature hepatocytes (MH) by treatment with dexamethasone, Oncostatin M, and HGF under hypoxia for four days and subsequently, under normoxic condition for eight days or more (Fig. 1A). Approximately 25 days after seeding, cyst-shaped 3D organoids were spontaneously generated from 2D monolayers of mature hepatocytes (Fig. 1B); the organoids (including a few free-floating organoids) were collected and embedded in Matrigel supplemented with HM. These conditions for maintaining functional hepatic organoids were previously optimized (Supplementary Table S1) (21). The organoids increased in size during culture with HM (Fig. 1C) and expressed the epithelial and hepatic markers, E-cadherin and Albumin (ALB), respectively (Fig. 1D).
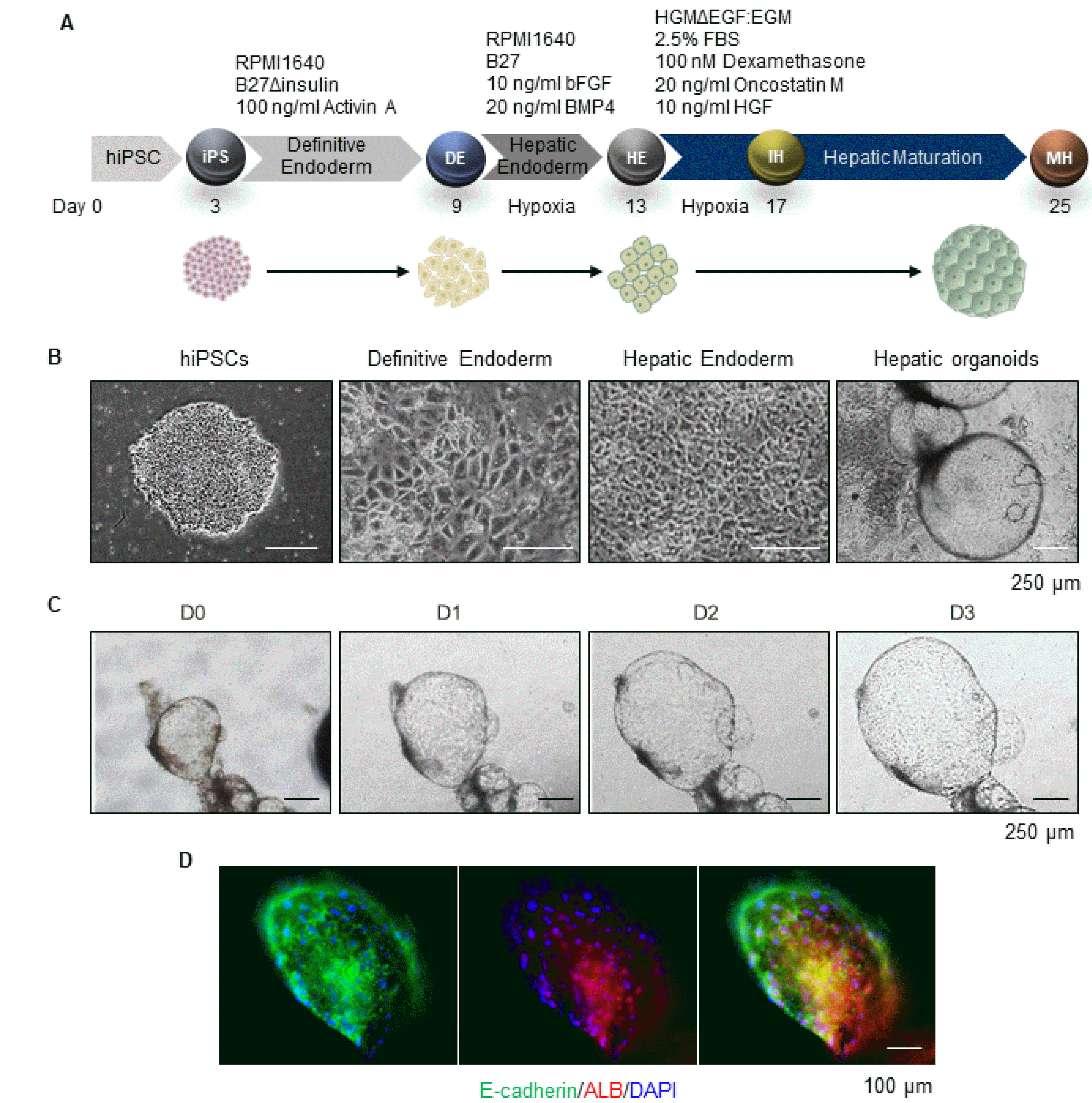
Fig. 1
Generation of hiPSCs-derived hepatic organoids. (A) Schematic diagram of the generation protocol from hiPSCs to hepatic organoid. (B) Representative bright field image of hiPSCs, definitive endoderm, hepatic endoderm, and hepatic organoids. (C) Bright field image of organoids immediately after Matrigel embedding (D0) and in the same field during culture (D1 to D3). (D) Representative immunofluorescence images of the hepatic organoids stained with E-cadherin and ALB.
![]()
Long-term culture of hepatic organoids without passaging
Matrigel-embedded organoids were maintained for four weeks and the medium was changed every alternate day (Fig. 2A, upper), since prolonged culture of organoids could enhance their maturity (25). The occurrence of damaged organoids markedly increased, and the Matrigel loosened and degraded over time with the appearance of dark granules after two weeks in culture (Fig. 2B, upper). Therefore, we replated the organoids weekly at a 1:2 ratio in fresh Matrigel (Fig. 2A, lower). Organoids in fresh Matrigel reached over 1.5 mm in size at four weeks and appeared healthier with a thick layer (Fig. 2B, lower) compared to those in Matrigel that had not been renewed (Fig. 2B, upper). The gene expression level of the functional mature hepatocyte marker ALB was sufficiently higher in the group grown on regularly renewed Matrigel, until three weeks, compared to that in the group grown on the same Matrigel. However, after four weeks, gene expression levels decreased drastically in both groups (Fig. 2C). Additionally, after five to six weeks, as the organoid size reached 2 mm, dark granules inside the organoids increased and organoids with dense morphology were appeared (Supplementary Fig. S1). Furthermore, the gene expression levels of the hepatic marker CK18 decreased after two to four weeks in both groups by half of that in 1-week control. The levels of the biliary/progenitor cell marker CK19 and the fetal hepatocyte marker AFP were considerably increased after long-term culture of the organoids (Fig. 2C). Therefore, we performed serial passaging of the organoids to resolve the size limit challenge and improve the functional maturity.
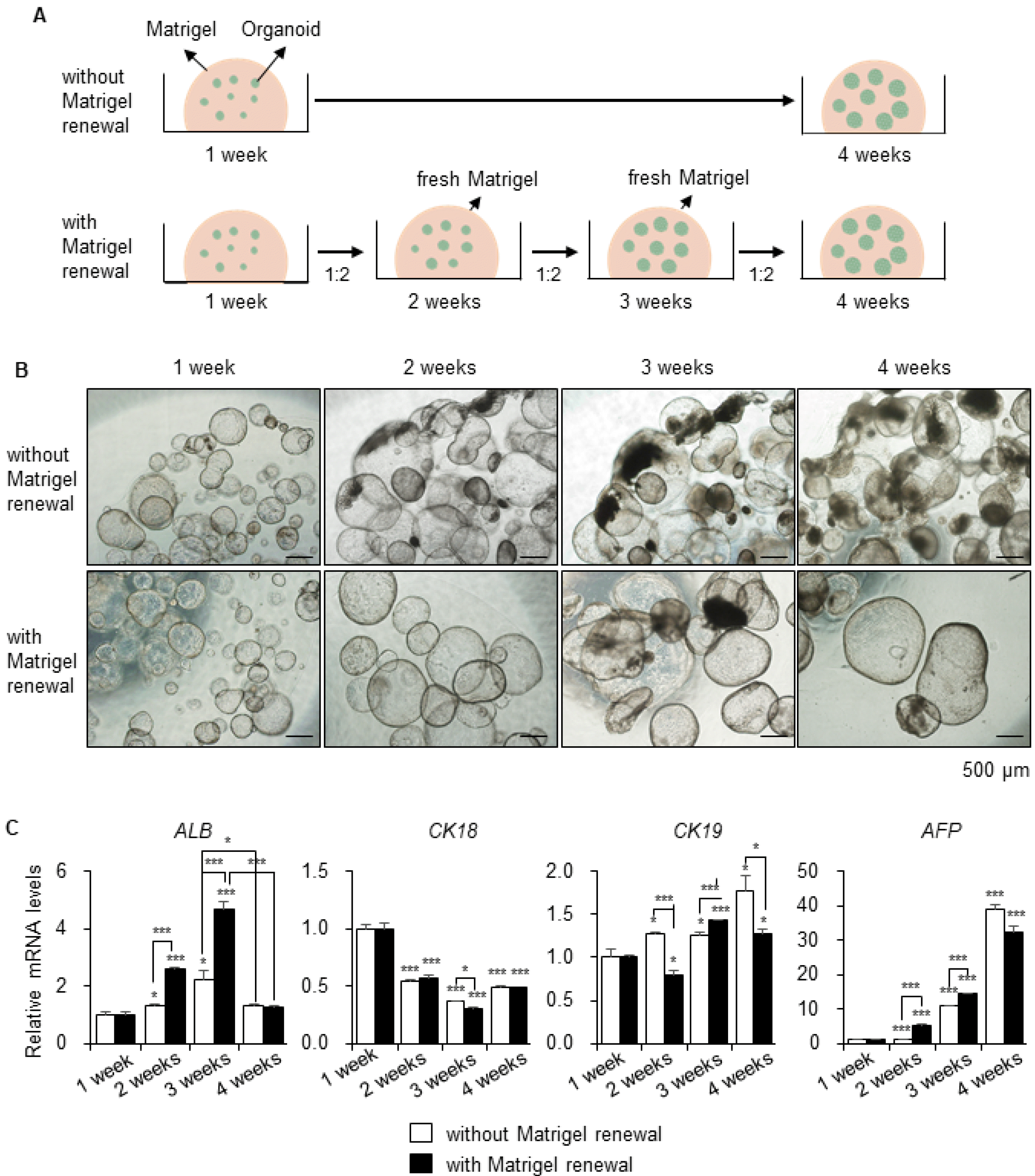
Fig. 2
Long-term culture of hepatic organoids without passaging. (A) Scheme of long-term culture of the organoids. Matrigel-embedded organoids were maintained for four weeks without Matrigel renewal (upper). Organoids were divided at a 1:2 ratio and replated on fresh Matrigel weekly (lower). (B) Morphology of the organoids in the same field from week 1 to 4 without Matrigel renewal (upper) and with Matrigel renewal (lower). (C) mRNA expression levels of ALB, CK18, CK19, and AFP in organoids without Matrigel renewal and with Matrigel renewal weekly. Data are the mean±SEM (n=3) and analyzed by Student’s t-test, *p<0.05 and ***p<0.001.
![]()
Long-term expansion of hepatic organoids by serial passaging
For long-term expansion of functional hepatic organoids, the organoids were mechanically split into 200∼250 μm sections by a sterile surgical blade under a dissecting microscope and resuspended them in fresh Matrigel at a 1:3∼1:5 ratio (Fig. 3A). Assessment of the morphology of small pieces of the organoids that were recovered, showed growth to over 0.6 mm in size after one week (Fig. 3A). Therefore, we passaged the organoids every seven days, which continued to expand far over passage 55, or 1 year, without growth retardation (Fig. 3B). Theoretically, approximately 6.5×1022 hepatic cells can be obtained from one hiPSC for 1 year. Moreover, organoids are susceptible to long-term cryopreservation, whereas freezing and thawing of 2D-cultured PHHs were challenging. The viability of organoids was 73±2.56% after thawing and they regrew well (Fig. 3C). Additionally, organoids were stably maintained a normal karyotype until at passage 40 and 50 (Fig. 3D). Finally, the gene expression levels of the functional mature hepatocyte marker ALB were maintained over long periods, and the levels of fetal hepatocyte marker AFP were remarkably decreased by passaging (Fig. 3E). Therefore, these data demonstrated that organoids can be exponentially expanded by serial passaging over long time periods, which decreases hepatic immaturity and also cryopreserved for long-term storage.
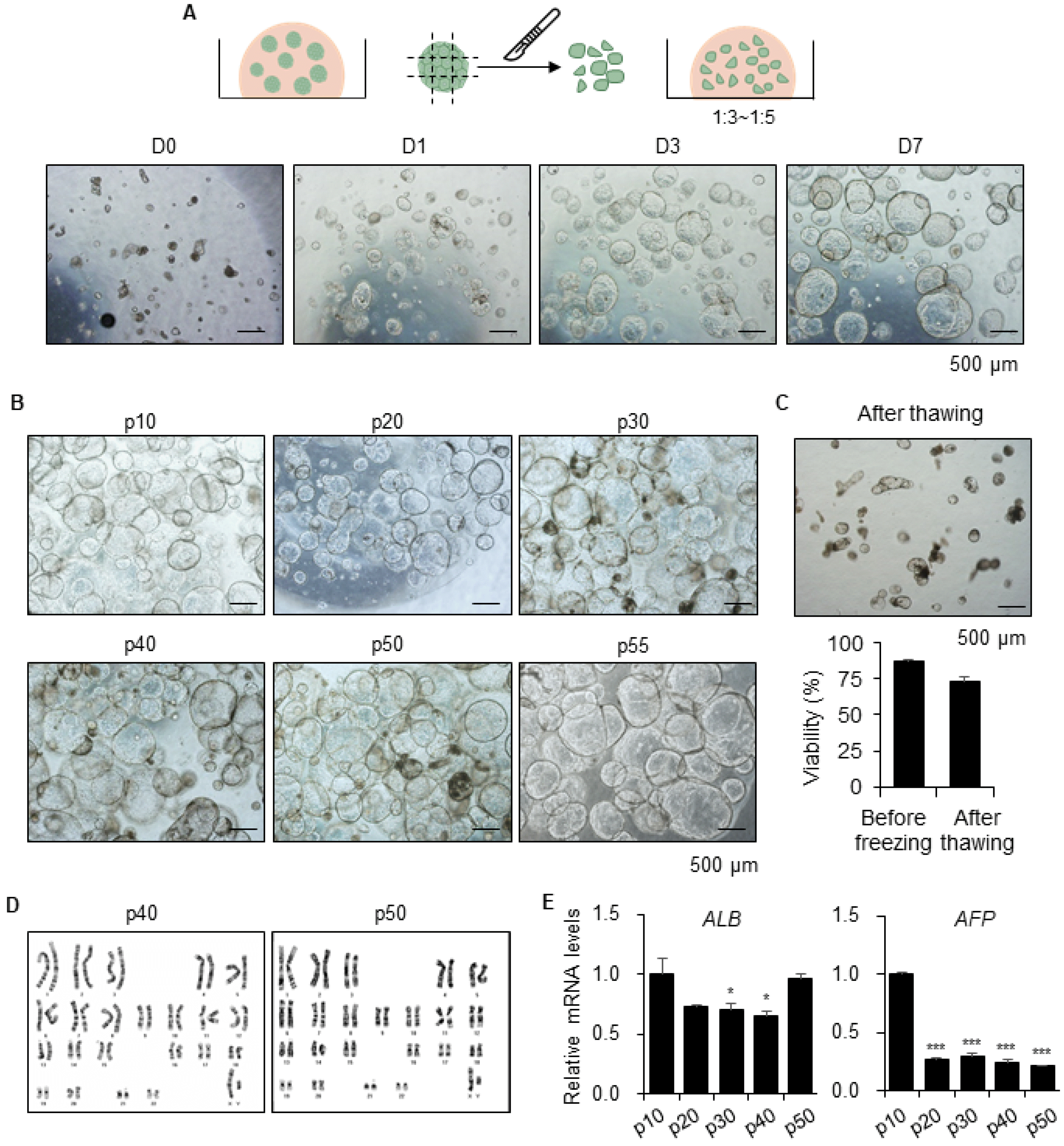
Fig. 3
Long-term expansion of hepatic organoids by serial passaging. (A) Schematic diagram of long-term culture of organoids by passaging (upper). Representative bright field image of hepatic organoids after passaging at day 0, 1, 3 and 7 in the same field (lower). (B) Representative morphology of each passage of hepatic organoids at day 7. (C) Representative morphology of organoids a day after thawing (upper). Cell viability was determined by cell counting with Trypan blue staining before freezing and 12 hours after thawing (lower). Data are the mean±SEM (n=12). (D) Karyotype analysis of the organoids at passage 40 and 50. (E) mRNA expression levels of ALB and AFP at every 10 passages. Data are the mean±SEM (n=3) and analyzed by Student’s t-test, *p<0.05 and ***p<0.001.
![]()
Differentiation potential of the long-term expanded hepatic organoids
A previous study found that organoids undergo hepatic maturation by further differentiation under Differentiation Medium (DM) (Fig. 4A) (21). The levels of gene expression of mature hepatic markers ALB, RBP4, and CYP3A4 were substantially increased in DM-cultured organoids at passage 10 (Fig. 4B). Consequently, we compared the differentiation potential of long-term expanded hepatic organoids after every 10 passages in DM, which revealed small and dense morphologies (Fig. 4C). Moreover, expression of ALB was maintained and that of RBP4 and CYP3A4 continuously increased (Fig. 4D). Therefore, these results implied that late passage organoids also have full differentiation potential until at least passage 50, and differentiation can be induced at any time point in vitro.
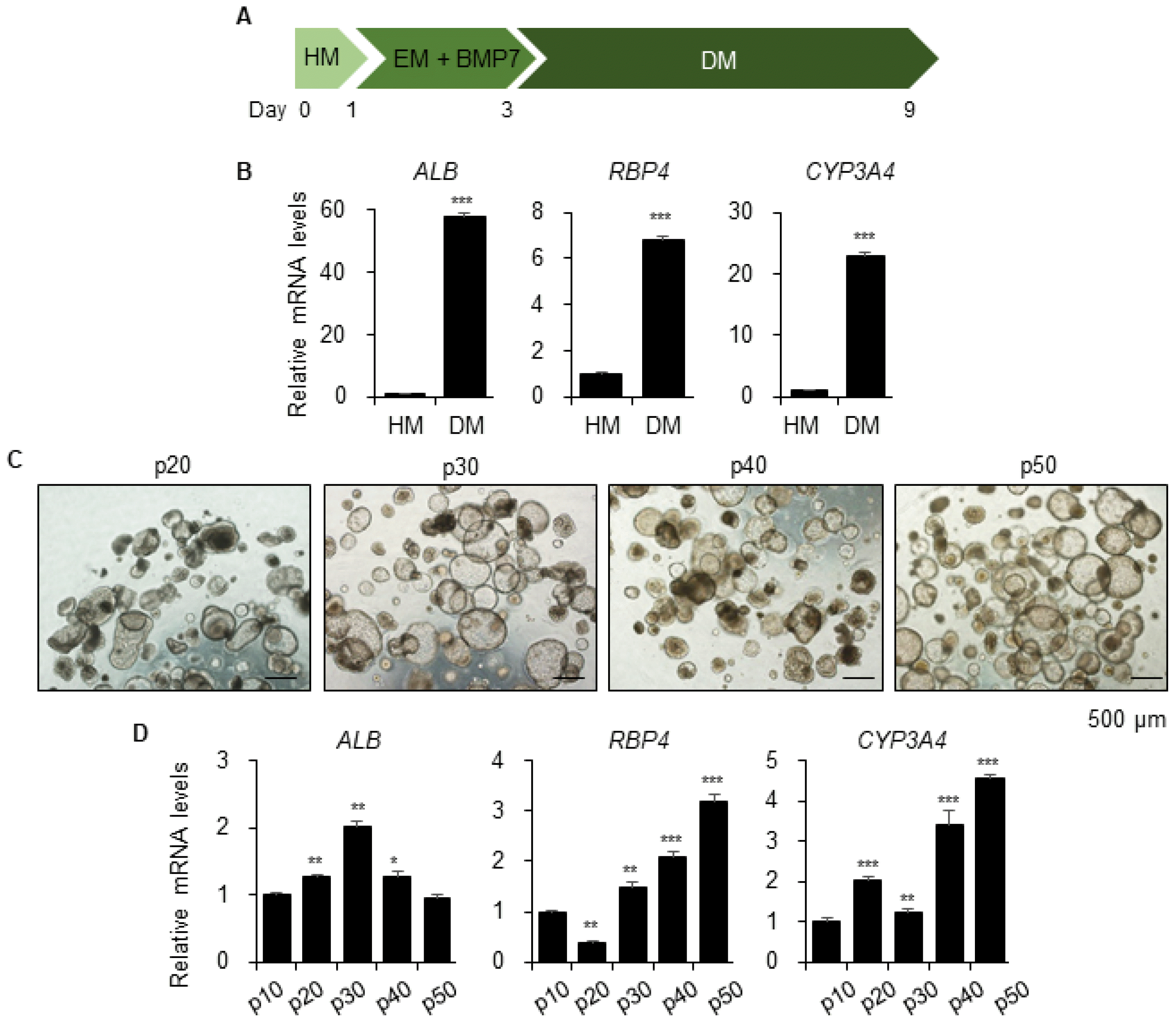
Fig. 4
Differentiation potential of the long-term expanded hepatic organoids. (A) Schematic diagram of organoid differentiation for further hepatic maturation. Hepatic medium (HM); Expansion medium (EM); and Differentiation medium (DM). (B) mRNA expression levels of ALB, RBP4, and CYP3A4 in HM- or DM-cultured organoids at passage 10. (C) Representative morphology and (D) mRNA expression levels of ALB, RBP4, and CYP3A4 of differentiated hepatic organoids at each indicated passage. Data are the mean±SEM (n=3) and analyzed by Student’s t-test. *p<0.05; **p<0.01; and ***p<0.001.
![]()
In this study, we described the methods for exponential and long-term expansion of our novel and functional human hepatic organoids by serial passaging. The proliferation and differentiation capacities of the organoids were maintained for long-term use. There was no evidence to cease proliferation and lose differentiation potential until passage 55. Therefore, this method can be used to address the challenge of a shortage of human hepatocytes. Hepatic organoids can be a practical and renewable source of liver cells for human cell-based and personalized in vitro 3D liver models.
Supplementary Materials
Supplementary data including three tables and one figure can be found with this article online at http://pdf.medrang.co.kr/paper/pdf/IJSC/IJSC-13-s20060.pdf.
 XML Download
XML Download