

 PDF
PDF Citation
Citation Print
Print


Introduction
Intervertebral disc (IVD) degeneration and consequent low-back pain have been the leading cause of disability especially in people aged less than 45 years old (1, 2), which presents a costly burden to the healthcare system. Conventional treatments for IVD degeneration are limited, since conservative or surgical therapies do not restore IVD tissue properties. Stem-cell based therapies that focus on rejuvenating and functionalizing tissue are becoming a promising strategy for IVD degeneration (3). Exogenous mesenchymal stem/stromal cells (MSCs), especially umbilical cord derived MSCs (UCMSCs), are currently the most popular graft cells for IVD regeneration because UCMSCs provide a nearly unlimited cell source with self-renewal capability and multilineage differentiation potential (4, 5). However, IVD microenvironment is characterized by high osmolarity, limited nutrition, acidic pH, and low oxygen tension (6, 7). Some studies have reported that exogenous stem cells are not able to survive well under IVD harsh microenvironment, as it negatively influences the biological and metabolic vitality of stem cells (8, 9). The IVD is composed of an inner nucleus pulposus (NP) surrounded by the annulus fibrosus (AF) and thin hyaline cartilaginous end-plates (EP) between the IVD and the adjacent vertebral bodies. Increasing evidence showed that cells isolated from the various compartments of human IVD, such as NP, AF, and EP expressed most of the phenotype markers that define MSCs, thus supporting the existence of resident stem/progenitor cells within IVD (10, 11). As seed cell candidate, the endogenous stem cells inside the IVD, especially nucleus pulposus-derived stem/progenitor cells, exhibit better adaptability to the harsh IVD microenvironment than the adipose-derived MSCs in tolerating hypoxic (12), hypertonic (13), and acidic culture conditions (14) after being transplanted into IVD. Therefore, the endogenous stem cells inside the IVD had been regarded as attractive seed cells and might perform better than exogenous MSCs for IVD regeneration (15). Nonetheless, the number of stem/progenitor cells that can be harvested and expanded from nucleus pulposus (NP) tissues in vitro are insufficient to meet requirements for successful treatment (16). In addition, culturing primary NP stem/progenitor cells for disc repair is challenging due to their limited expansion capability (17, 18). These issues have motivated the exploration of rejuvenating regenerative capacity of NP derived stem/progenitor cells from degenerated IVD.
It has been reported that MSCs exhibit a number of trophic functions to enhance tissue regeneration (19). Recent studies have suggested that MSCs can promote the viability of NP cells in animal models and in preclinical studies (20, 21). Sun et al. (22) noted that adipose-derived stem cells protect compressive load induced NP cell death and degradation by inhibition of activated caspase-9 and -3 activity in the co-culture system. Yamamoto et al. (23) observed that bone marrow derived MSCs may improve the viability of NP Cells in vitro. Results from these studies indicated that the mechanism underlying MSCs efficacy depends mostly on their paracrine activity.
However, researches reporting the ability of MSC secretome to promote the functionality of NP cells are just emerging. Whether the trophic factors secreted by MSCs can ameliorate the biological function of NP derived stem/progenitor cells from degenerated disc (D-NPSCs) is not fully understood, and continuous laboratory investigations are indispensable to explore the possibility of employing trophic factors secreted by MSCs to promote the regenerative capacity of D-NPSCs and further develop a stem cell-based treatment for degenerated IVD.
Despite the presence of many different types of stem cells, UCMSCs have been recently considered more potent as cell source than bone marrow MSCs or adipose-derived MSCs since they proliferate/differentiate more efficiently and have a noninvasive harvesting procedure as well as abundant cell sources (24). Thus, in this study, we collected conditional medium from human UCMSCs (UCMSCs-CM) to treat D-NPSCs, which were isolated from degenerated NP tissues obtained from patients who underwent micro endoscopic discectomy for degenerative spine diseases. We found that UCMSCs-CM exposure restored biological activities of D-NPSCs detected by examining the pluripotent markers, proliferative capacity, multi-lineage differentiation potential, and the expression of NP-specific cell surface markers. Our finding will lay a foundation for the development of optimal therapeutic strategy for IVD degeneration.
Materials and Methods
Ethics approval
This study was approved by the medical ethics committee of Jinan University, Guangzhou, China. All procedures performed for the present work were approved specific informed consent was obtained in all cases.
Sample collection
Mildly as well as severely degenerated NP tissue samples were obtained from patients (n=8) who underwent micro endoscopic discectomy for degenerative spine diseases. The details of all samples and patients are shown in Table S1 of the Supplementary File.
Umbilical cord were obtained from newborns (n=5) who exhibited no complications throughout pregnancy and were delivered full-term by Caesarean section.
Isolation of cells from NP tissues or umbilical cord
Isolation of cells from NP tissues: The disc tissue samples were collected under sterile conditions immediately following surgery, washed with phosphate-buffered saline (PBS, pH=7.2). The NP tissue were obtained by removing anulus fibrosus tissue under stereoscopic microscope. The explant culture method was employed to isolate NPSCs from NP tissue as previously described (25). In brief, NP tissues were finely minced into small fragments of approximately 1 mm3, and incubated at 37℃ in a humidified atmosphere containing 5% CO2 without culture medium for two hours to allow the tissue pieces to attach. We used Dulbecco Modified Eagle Medium (DMEM)/F-12 (Gibco, USA) supplemented with 10% fetal bovine serum (FBS) (Gibco, USA), 1% L-glutamine, and 1% penicillin-streptomycin (Invitrogen, USA) as complete culture medium, which was added to the tissue culture dishes and incubated for an additional 15∼18 days. The medium was changed every two days. The pieces were removed when the cells reached 80% confluency, and the cells were passaged by a 2-min treatment with 0.25% trypsin and 0.02% EDTA at 37℃. Cells at Passage 2∼4 cells were used in the experiment.
Isolation of MSCs from human umbilical cord: UCMSCs were isolated and cultured as described previously (26). Briefly, umbilical cord tissue was washed twice with PBS. A 10 cm umbilical cord was cut into small pieces of about 1 cm in length. The pieces were washed thoroughly to remove blood and blood clots, the Wharton’s jelly was separated and dissected into small pieces of approximate 1∼2 mm3 pieces, then plated in the complete culture medium and cultured for 12 days. The medium was changed every third day. At 80%∼90% confluence, UCMSCs were detached and passaged. Cells at Passage 3∼6 were used for all experiments.
Collection of conditioned medium from UCMSCs cultures: UCMSCs were cultured and expanded in the complete culture medium. At passage 3∼6, a total of 1×106 cells were seeded into 100 mm2 cell culture dishes. When the cells reached 60% confluence, they were washed with PBS and then cultured in serum-free complete culture medium for an additional 24 h. The cell supernatant was collected and filtered with 0.22 micron filter to remove living cells, dead cells and cellular debris as conditioned medium, stored at −80℃ until use. When used, the conditioned medium would be mixed with DMEM/F12 at the concentration of 40%, which was further supplemented with 10% FBS, 1% L-glutamine and 1% penicillin-streptomycin, as UCMSCs-CM.
To determine the optimal percentage of the conditioned medium, we performed the pre-experiment to evaluate the effects of 20%, 40% and 80% of the conditioned medium on the proliferative capacity of D-NPSCs by using CCK8 method. The result showed that the OD values of 40% were the highest compared with other groups (data shown in Fig. S1 of the Supplementary File).
D-NPSCs in passage 2∼4 were plated at 5×104 cells/well in 24-well culture plates, then treated with UCMSCs-CM, as experimental group (CM group), the untreated D-NPSCs was taken as negative control (D-NPSCs group) and UCMSCs was acted as positive control (UCMSCs group). The media used in D-NPSCs and UCMSCs group were complete culture medium, which contained DMEM/F12 supplemented with 10% FBS, 1% L-glutamine and 1% penicillin-streptomycin.
Flow cytometry assay
The expression of specific surface markers: The characteristics of the cells from D-NPSCs, CM and UCMSCs group were investigated further by analyzing the cell surface markers with flow cytometry. Cells at passage 2∼4 were harvested and washed with PBS after 72 h treatment of UCMSCs-CM, and every 1×106 cells were resuspended in 100 μl cold PBS. Fluorochrome-coupled anti-mouse monoclonal antibodies (including CD29-FITC, CD44-PE, CD73-PE, CD90-PC5, CD105-PE, CD11b-PE, CD14-FITC, CD24-FITC, CD34-PE, CD45-PC5, CD45-PerCP and HLA-DR-FITC) were added to the corresponding tubes, and were incubated with the cell suspension at 4℃ for 30 min in the dark. Isotype control antibodies served as controls. Finally, labeled cells were washed three times with PBS and surface markers expression was detected using flow cytometry (BD Biosciences, USA). The percentage of positively stained cells was calculated relative to the isotype control.
Cell cycle analysis: Cells from D-NPSCs, CM and UCMSCs group were harvested after 72 h treatment of UCMSCs-CM, washed twice with cold PBS, and fixed in 75% pre-chilled ethanol for over 12 hours at 4℃. The fixed cells were washed twice with ice-cold PBS, then stained with propidium iodide (PI, 500 ml/106 cells) at room temperature for 30 min in the dark. The intensity of fluorescence was determined by flow cytometry, and the percentages of cells in G0/G1, G2/M and S phases were analyzed using BD CellQuest software. The experiment was repeated three times to ensure reproducibility.
Apoptosis analysis: Cell apoptotic fraction was detected by staining with FITC-conjugated annexin-V and PI using the Annexin-V/PI Apoptosis Detection kit. According to the manufacturer’s instructions, all cells were harvested after 72 h treatment of UCMSCs-CM, washed twice with PBS, and resuspended in 300 μl of binding buffer. 5 μl Annexin V-FITC solution was added to the cells and incubated for 30 min at 4℃ in the dark, which was followed by further incubation with 5 μl PI for 5 min. The percentage of apoptotic cells were analyzed immediately by flow cytometry, and the experiment was repeated three times independently.
Cell viability and proliferation analysis
Growth curve: Growth curve was plotted to evaluate growth kinetics of the cells after 13 days incubation of UCMSCs-CM. Cells for each group were seeded at a density of 3×104 cells/well in 24-well plates. The cells were harvested by trypsinization from each well, and a duplicate was collected and counted with a hemocytometer count at day 1, day 3, day 5, day 7, day 9, day 11 and day 13. The growth curve was plotted with the cell culture time as the horizontal axis and the number of cells as the vertical axis. The results were analyzed by GraphPad Prism software.
Cell viability: Cell viability was determined using the Cell Counting Kit 8 (CCK-8, Dojindo Laboratories, Japan). A total of 3×104 cells/well were seeded in 24-well plates, CCK-8 solution was added at day 1, 3, 5 and 7, and incubated for a further 2.5 h. The absorbance of the supernatant was measured with a micro plate reader (Bio-Rad, USA) at wavelength of 450 nm. The CCK8 assays were performed in triplicate.
EdU incorporation assay: The EdU (5-ethynyl-2’-deoxyuridine) proliferation assay was performed to further explore the role of UCMSCs-CM using Cell-LightTM EdU Apollo 567 In Vitro Kit (Ribobio, China) following the manufacture’s recommendation. In brief, cells were seeded into 96-well plates at 5×103 cells/well, and cultured for 72 h. Then 100 μl medium containing 50 μM EdU was added into each well and cells were incubated for 2 h at 37℃, fixed with 4% par formaldehyde (PFA), then stained with Hoechst 33342 and Apollo reaction cocktail. All images were processed using Image J software and the proportion of EdU incorporated cells was calculated. Three independent experiments were performed for quantification.
Multilineage differentiation
Osteogenic and adipogenic differentiation of sub confluent cells was induced by induction media for 21 days as previously described by Richardson et al. (27) after 72 h treatment of UCMSCs-CM. For chondrogenesis, 3.5×106 cells were treated with UCMSCs-CM for 72 h, then centrifuged to form a three-dimensional aggregate in a 15 ml conical tube, and incubated with chondrogenic media for 21 days. The induced medium was replaced every 2∼3 days. For Alizarin Red S (ARS) staining, cells were fixed in 4% PFA for 10 minutes and stained for 5∼10 min with 40 mM ARS solution. Oil red O solution was used to stain intracellular lipid vacuoles for 5∼10 min after fixation with PFA. To identify chondrogenic differentiation, the cell pellets were stained with Alcian blue, which were fixed in PFA, then frozen in OCT freezing medium (Sakura Finetek), sectioned into 5μm-thick slices. The results were photographed under phase contrast microscope, and all images were processed using Image J software, the proportion of area positively stained was calculated. The experiment was repeated 3 times independently.
The expression of marker genes detected with quantitative real-time polymerase chain reaction (qRT-PCR)
Quantitative real-time polymerase chain reaction (qRT-PCR) was used to determine relative gene expression. The expression of NP-specific markers genes Aggrecan (ACAN), COL2A1, SOX9, Tie2, and CD24, and pluripotent stem cell-related genes (OCT4 and Nanog) were evaluated after 72 h incubation of UCMSCs-CM. All primers were designed according to the sequences in GenBank using software prime 5.0. Sequences of primers were shown in Table 1. Cells were harvested, and total RNA was extracted using Trizol reagent (Invitrogen, USA), and quantified by a Nanodrop 2000 spectrophotometer (Thermo Fisher, USA). RNA (1 μg) was used for cDNA synthesis using a Cell to cDNA Kit. Real-time PCR was then performed in a final volume of 20 μl with 2×SYBR Green qRT-PCR Master Mix. Each reaction was performed in triplicate. The housekeeping gene GAPDH was used for endogenous control. The 2−△△Ct method was used to describe the relative expression levels of the gene.
Table 1
Primers used for this study
![]()
Results
To investigate effects of the conditioned medium derived from UCMSCs on regenerative capacity of NP resident stem/progenitor cells from degenerated IVD, D-NPSCs were treated with the UCMSCs-CM at 40% of the conditioned medium and compared with untreated D-NPSCs and UCMSCs in term of immunophenotype, proliferation, multi-lineage differentiation abilities, and the expression levels of NP related markers.
UCMSCs-CM enhanced expression of MSC surface markers in D-NPSCs
Flow cytometry results showed that UCMSCs were highly positive for CD29, CD44, CD73, CD90 and CD105 at rates greater than 95% , and negative for CD11b, CD14, CD34, CD45 and HLA-DR (Fig. 1), which fulfilled the international Society for Cell Therapy (ISCT) requirements for MSC definition; The D-NPSCs before UCMSCs-CM treatment, although expression of CD73, CD44 and CD90 were at rates greater than 95% and expression of CD11b, CD14, CD34, CD45 and HLA-DR were negative, expression of CD29 and CD105 showed interindividual variability and all at rates lower than 95%. However, CD29 and CD105 were significantly up-regulated at rates greater than 95% after 72 h UCMSCs-CM treatment (Fig. 1, p<0.05), therefore CM group exhibited a very similar expression pattern of surface markers with UCMSCs group.
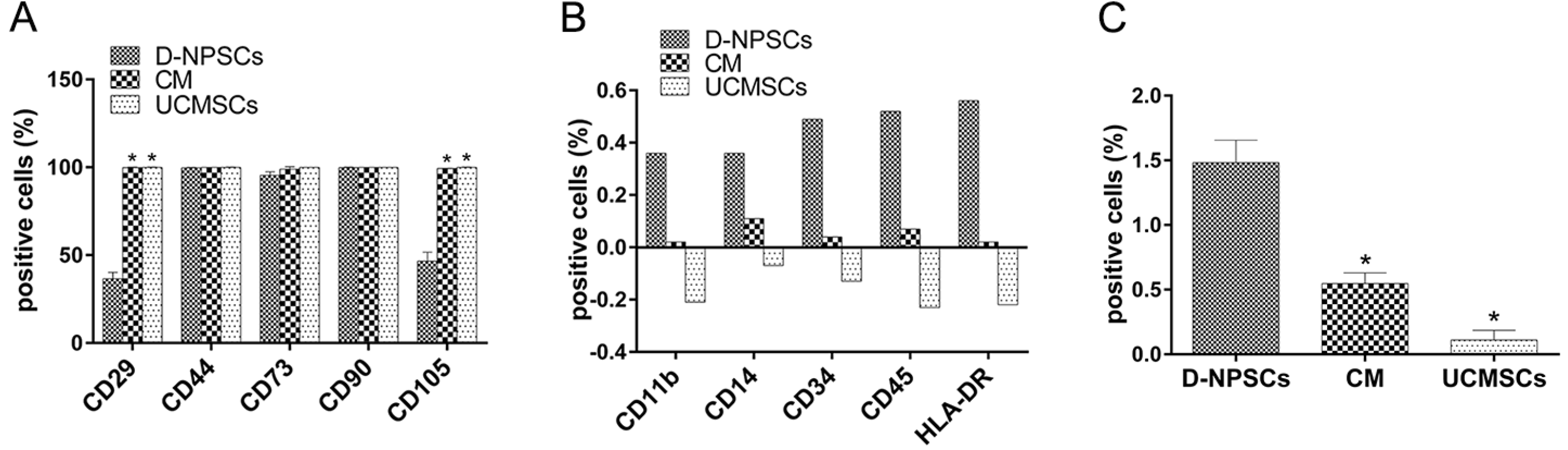
Fig. 1
The MSC and NP cell surface markers expression were analysis by flow cytometry after 72 h treatment with UCMSCs-CM. (A) MSC positive surface markers, (B) MSC negative surface markers, (C) NP cell surface marker CD24. The data are presented as the means±SD, n=3. *p<0.05, compare with D-NPSCs group. UCMSCs were positive for CD29, CD44, CD73, CD90 and CD105 at rates greater than 95%. For D-NPSCs, the expression of CD29, and CD105 showed interindividual variability but all at rates lower than 95%, however, CD29 and CD105 in CM group were at rates greater than 95%, and the CM group revealed a very similar expression pattern of surface markers with UCMSCs group; Cells in all three groups were negative for the hematopoietic stem cell surface markers CD11b, CD14, CD34, CD45 and HLA-DR. Cells in all three groups expressed CD24 at low rates, while the rate in D-NPSCs significantly higher than that in both CM group and UCMSCs group.
![]()
UCMSCs-CM promote proliferation and viability in D-NPSCs
Growth curve: According to the growth curve, all cells proliferation was slow during the first day of culture (latent phase) as shown in Fig. 2A. Both UCMSCs and CM groups were accelerated rapidly during day 3∼5 (logarithmic phase), and slowed down thereafter (stationary phase), while D-NPSCs proliferated still slowly on day 3, and then entered the logarithmic growth phase at day 5, which continued for 5∼6 days, and reached cell growth plateau at day 10∼13. These results demonstrated that the growth ability of cells in CM group was markedly higher than that in D-NPSCs group.
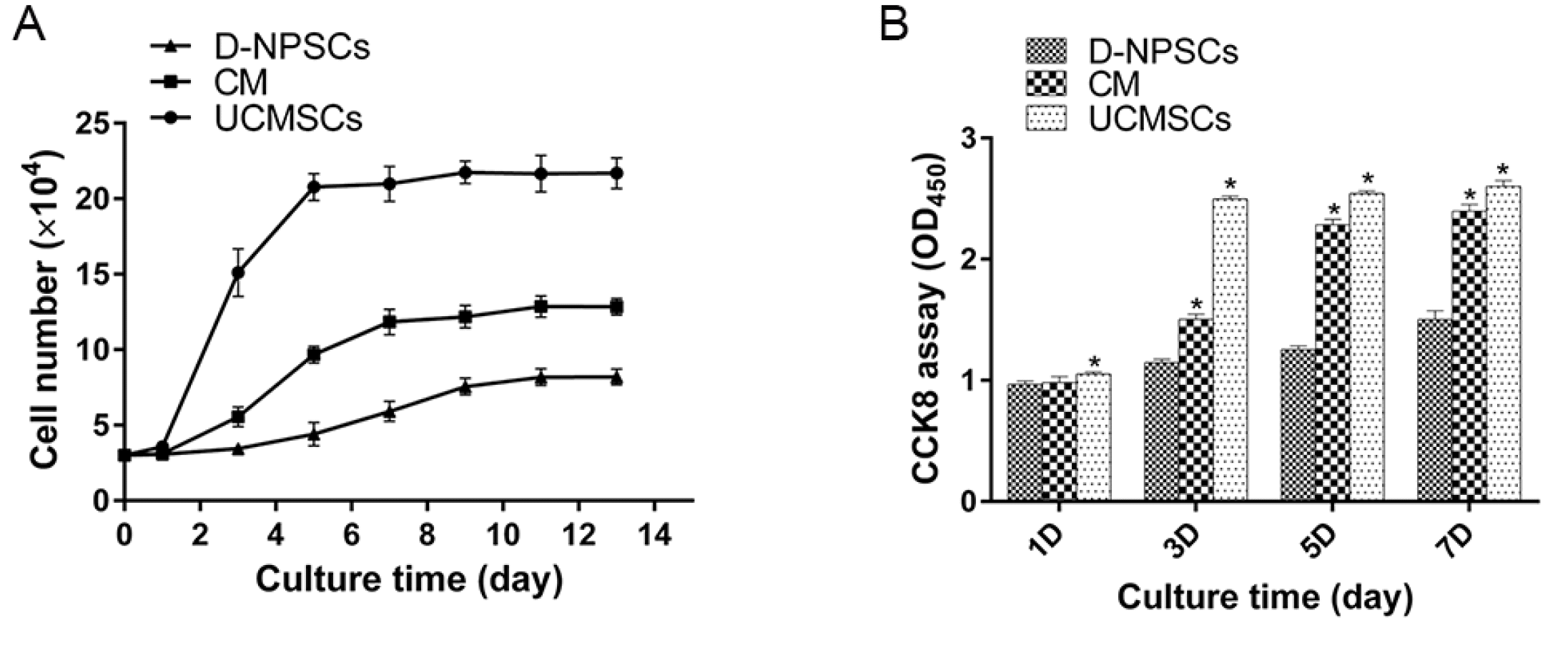
Fig. 2
Proliferation capability analysis of cells in the three groups. (A) Growth curves, (B) Cell viability was analyzed by cell counting kit-8 on the 1st, 3rd, 5th, and 7th day. The data are expressed as mean±SD, n=3, *p<0.01 vs. D-NPSCs. (A) Both UCMSCs and CM group were accelerated rapidly during days 3∼5 (logarithmic phase), and slowed down thereafter (stationary phase), while D-NPSCs in the days 3, cells proliferated slowly and then entered the logarithmic growth phase, which continued for 5∼6 days, and reached cell growth plateau in 9∼13 days. (B) Cells from CM group exhibited a greatly increased OD value compared with D-NPSCs group at day 3, 5 and 7.
![]()
Cell viability analyzed by CCK8 method: The viability of D-NPSCs and UCMSCs was assessed with CCK8 method as shown in Fig. 2B. The OD values of cells from both CM group and UCMSCs at day 3, 5 and 7 were significantly higher than D-NPSCs, which was consistent with the results of growth curves. The CM group reached to a similar OD value with UCMSCs group at day 5, 7.
EdU analysis: The results showed that cells in CM group had markedly higher proportion of EdU incorporated cell than D-NPSCs group after 72 h UCMSCs-CM treatment (Fig. 3, p<0.01), although lower than UCMSCs group, which suggested that UCMSCs-CM promoted the DNA replication and cell growth in D-NPSCs.
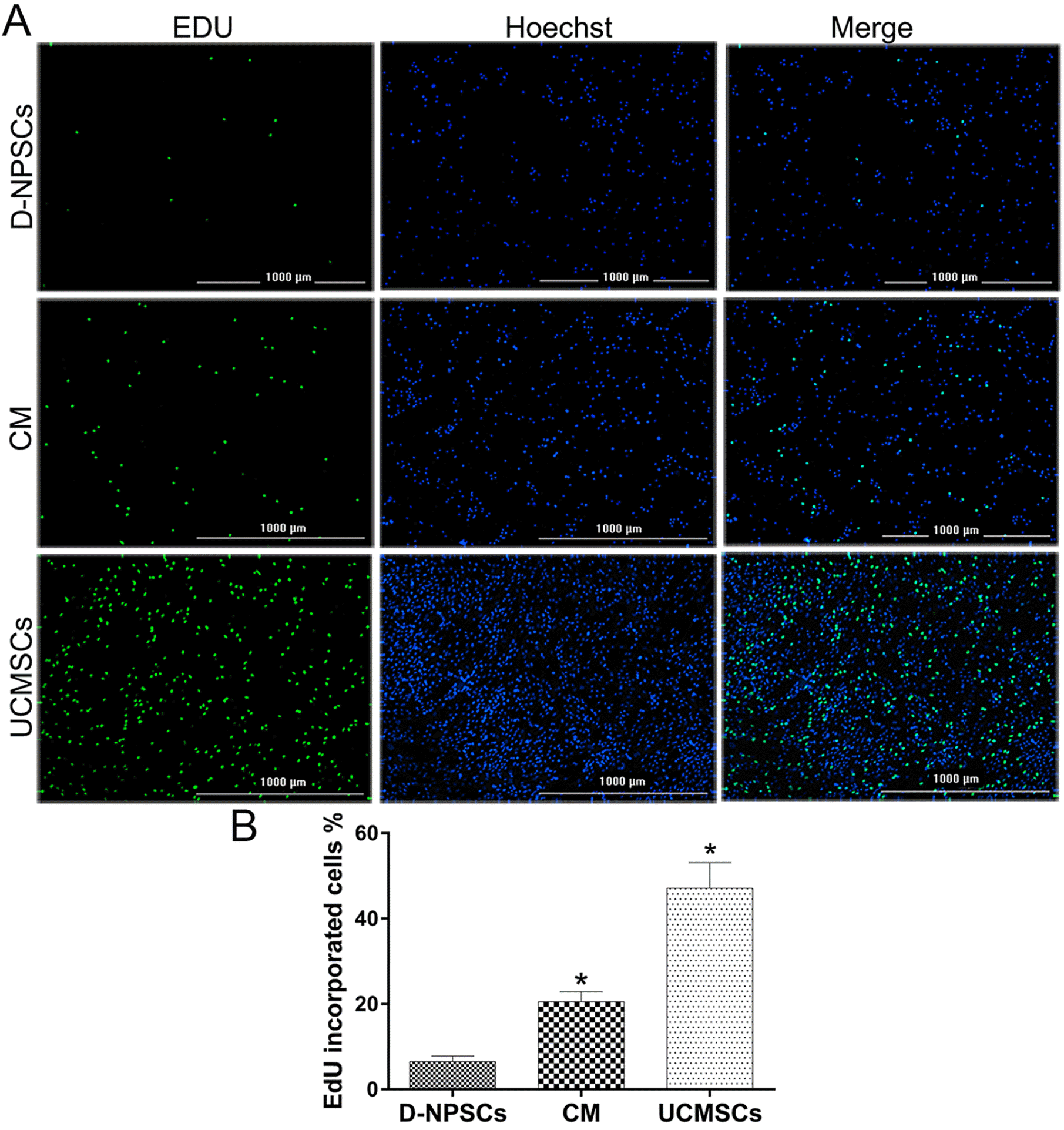
Fig. 3
EdU proliferation assay after 72 h treatment with UCMSCs-CM. (A) EdU incorporated cells in the three groups. (B) Comparative analysis of the percentage of EdU incorporated cells in the three groups. Scale bar=1000 μm (magnification 40×). The data are presented as the means±SD, n=3. *p<0.01, compare with D-NPSCs group. (A) Cells in D-NPSCs group had markedly lower proportion of EdU incorporated cell than CM group and UCMSCs group. (B) Further quantitative analysis also revealed that the percentage of EdU incorporated cell was significantly lower in D-NPSCs than that in CM group and UCMSCs group.
![]()
Cell cycle and apoptosis analysis: Cell cycle were assessed using flow cytometry, the percentage of cells in the G1/G0, G2/M and S phases of cell cycle was then analyzed at 72 h following UCMSCs-CM treatment. The results revealed that cells in the CM group had significantly higher percentages of cells in the S phases and lower percentages of cells in the G1/G0 phase than that in the D-NPSCs group (Fig. 4A, p<0.05), which is similar to cells in the UCMSCs group. The frequency of cell apoptosis in the CM group was significantly decreased when compared with the D-NPSCs group after 72 h UCMSCs-CM treatment (Fig. 4B, p<0.05), and tended to be higher compared with the UCMSCs group.
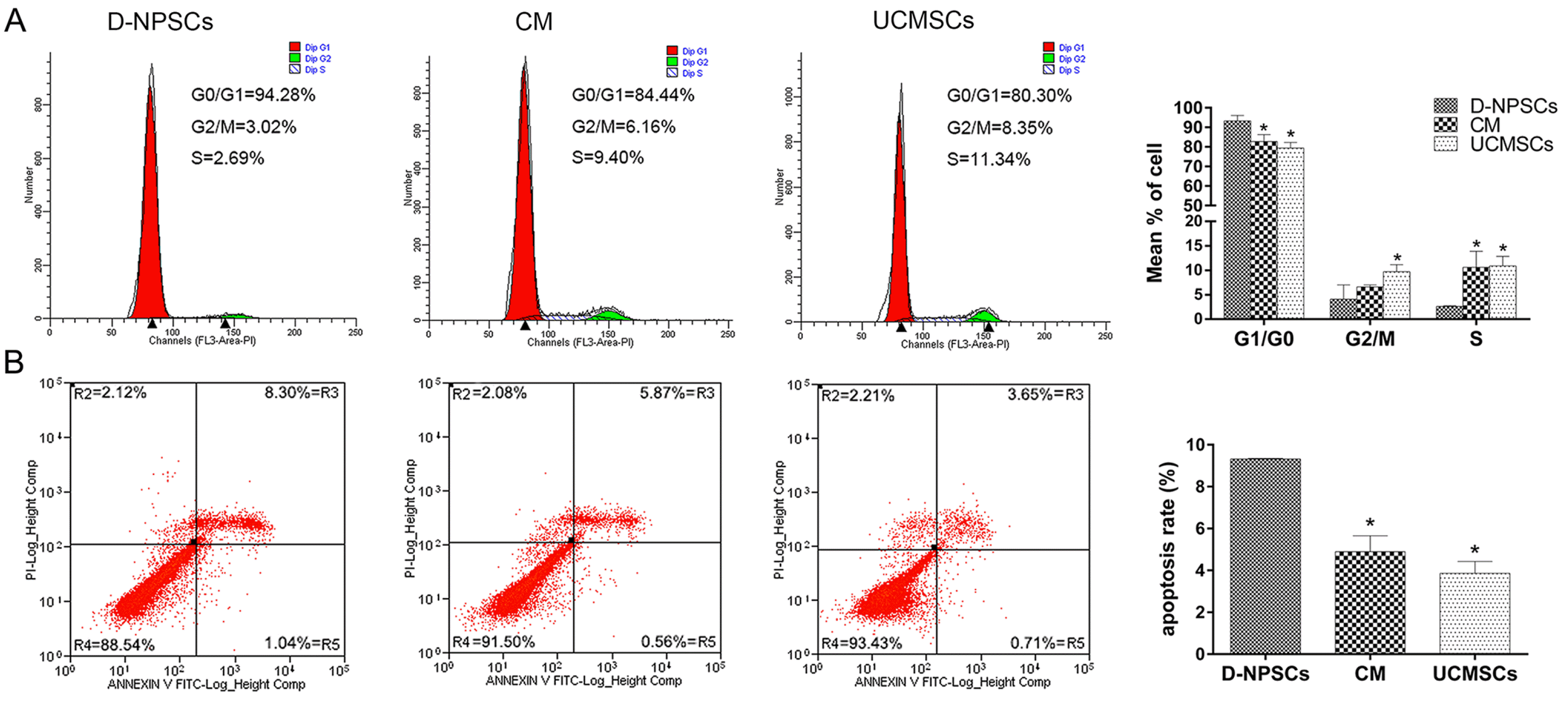
Fig. 4
The cell cycles (A) and apoptosis (B) were assessed after 72 h treatment with UCMSCs-CM by flow cytometry. The data are expressed as means±SD, n=3, *p<0.05 vs. D-NPSCs group. CM group had significantly higher percentages of cells in the S phases and lower percentages of cells in the G1/G0 phase than D-NPSCs group, and showed a similarity with UCMSCs group (A). The cell apoptosis rate in CM group was significantly decreased compared with D-NPSCs group, and tended to be higher compared with UCMSCs group (B). Data are presented as the means±SD, n=3. *p<0.05, compare with D-NPSCs group.
![]()
Collectively, the proliferation and viability of cells in CM group were greatly higher than that of D-NPSCs group, indicated that UCMSCs-CM promoted stem/progenitor cell growth from degenerated nucleus pulposus by slowing down the process of cell apoptosis and driving more cells into the DNA synthesis phase.
Multilineage differentiation potential analysis
Multilineage differentiation potential were analysised when the cells were incubated for 21 days in adipogenic, osteogenic and chondrogenic media following UCMSCs-CM treatment.
Osteogenic Differentiation D-NPSCs exhibited few calcium deposition stained by ARS as observed in Fig. 5A, whereas the cells from the CM group displayed larger and more intensely stained mineralized nodules (p<0.01) though it presented less intense staining than UCMSCs.
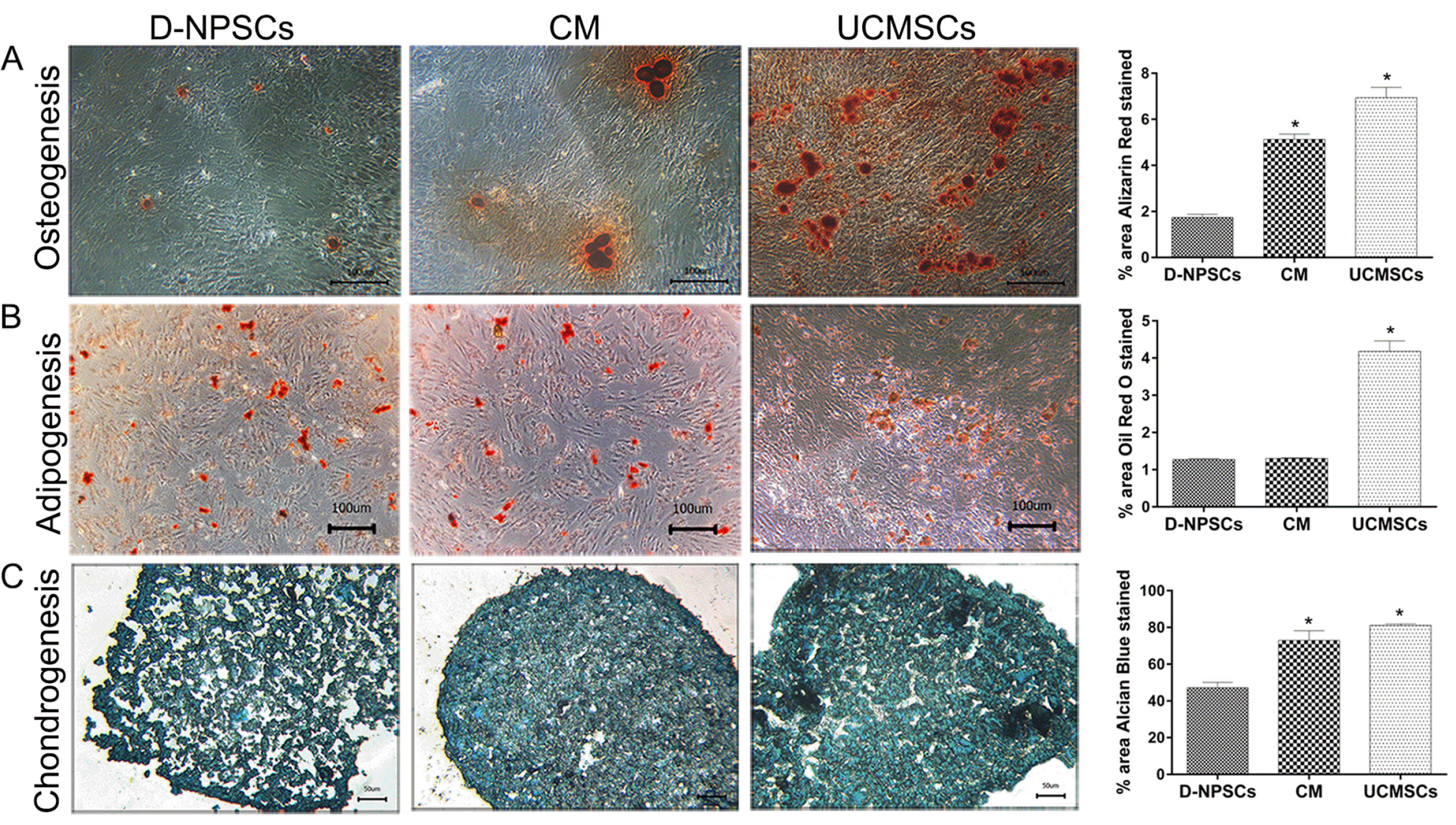
Fig. 5
Multipotent differentiation potential analysis after 72 h treatment with UCMSCs-CM. (A) Osteogenic differentiation for 21 days, Scale bar=100 μm (magnification 100×), (B) adipogenic differentiation for 21 days, Scale bar=100 μm (magnification 100×), (C) chondrogenic differentiation for 21 days, Scale bar=50 μm (magnification 200×). The data are presented as the means±SD, n=3. *p<0.01, vs. D-NPSCs group. D-NPSCs exhibited few calcium deposition stained by Alizarin red S, whereas the cells from CM group displayed larger and more intensely stained mineralized nodules though it presented less intensely staining than UCMSCs. (A) There were no significant difference in Oil red O positive staining area between the CM group and D-NPSCs group, both appeared to form less fat drops than UCMSCs as shown in (B); Cells from CM group produced more intensely stained extracellular matrix than D-NPSCs group, showed similar intensity levels with UCMSCs group. (C) For osteogenic and chondrogenic differentiation, further quantitative analysis also revealed that the percentage of area stained positively was significantly lower in D-NPSCs group than that in both CM group and UCMSCs group.
![]()
Adipogenic Differentiation Oil red O was used to stain lipid-rich vacuoles to analyze for adipogenesis. Cells from all the three groups showed adipogenic differentiation. However, there were no significant difference in positive staining area between the CM and D-NPSCs groups (Fig. 5B), and cells from both groups appeared to form less fat drops than UCMSCs.
Chondrogenic Differentiation Cells from CM group produced more intensely stained extracellular matrix than D-NPSCs group (Fig. 5C, p<0.01), but a similar staining intensity compared with UCMSCs group.
These data indicated that UCMSCs-CM improved the osteogenic and chondrogenic potential of D-NPSCs.
UCMSCs-CM altered the expression level of NP related markers
Results in Fig. 6 showed that the expression levels of NP cell phenotypic markers, SOX9, COL2A1 and ACAN in CM group were significantly lower than those in D-NPSCs group after 72 h UCMSCs-CM treatment (Fig. 6, p<0.01). Meanwhile, the mRNA expression of Tie2, a NP progenitor cell specific marker, and NP cell marker CD24, were also evaluated. The results showed that Tie2 expression was significantly up-regulated, while CD24 was significantly down-regulated in CM group compared with D-NPSCs group (Fig. 6, p<0.01).
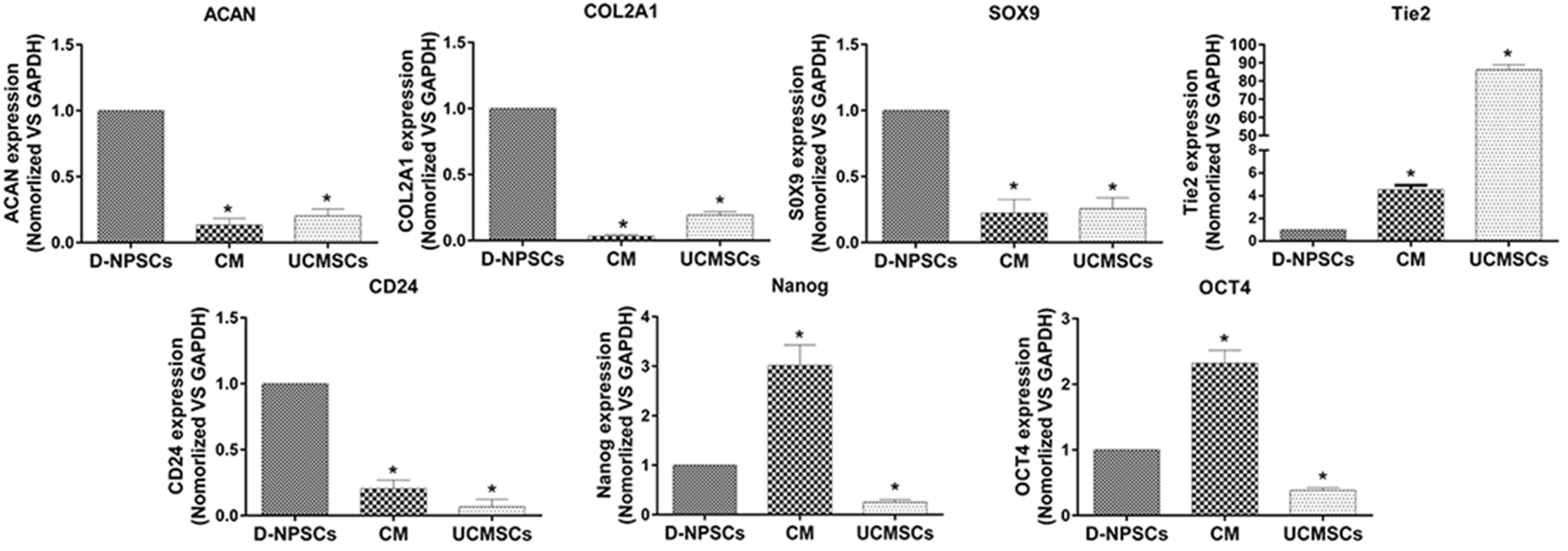
Fig. 6
qRT-PCR for ACAN, COL2A1, SOX9, CD24, Tie2, OCT4 and NANOG was performed after 72 h treatment with UCMSCs-CM and data were presented as relative ratio to GAPDH. The data are expressed as mean±SD. n=3, *p<0.01 vs. D-NPSCs. The expression levels of NP cell phenotypic markers, SOX9, COL2A1, ACAN and CD24 in CM group were significantly lower than those in D-NPSCs, while the level of Tie2, OCT4 and NANOG were significantly upregulated in CM group compared to D-NPSCs group.
![]()
Additionally, we had chosen CD24 for flow cytometry analysis, found that CD24 in CM group had a significantly lower positive rate than that in D-NPSCs group (Fig. 1C, p<0.05), although higher relative to UCMSCs group, and this result was in accordance with the qRT-PCR analysis results.
OCT4 and Nanog were evaluated as pluripotent stem cell markers and the results indicated that the expression levels of these two genes were greatly up-regulated after 72 h UCMSCs-CM treatment (Fig. 6, p<0.01).
Discussion
MSCs-based therapies had been proposed as novel treatments for IVD degeneration. Although the initial applications conceived for MSCs therapy were based on their multilineage differentiation capacity, subsequent research showed that MSCs engraftment and differentiation at injury sites were very low and transient (28). Increasing evidence indicated that the cytokines and growth factors secreted by MSCs were antiapoptotic, proangiogenic and had the potential to enhance the regenerative capacity of other cells (29-32). Thus, it was hypothesized that MSCs exert their therapeutic effects mainly through secreted trophic factors rather than differentiation. Several studies have shown that MSC-CM can promote the viability and migration of a variety of cells, such as keratinocytes, myocardial cells, endothelial cells and fibroblasts in vitro and in vivo (33-35). However, few studies focused on the therapeutic effects of MSC-CM on degenerated tissue specific progenitor cells.
In the present study, we explored whether MSCs secreted factors can exert beneficial stimulatory effects on D-NPSCs, and characterized changes in immunophenotyping, expression of characteristic NP markers, proliferative capacity and differentiation potentials following treatment of UCMSCs-CM. We observed the expression of MSC specific surface markers CD29 and CD105 was dramatically increased after treating with UCMSCs-CM. Our previous data suggested that NPMSCs from young health tissues expressed all the MSC markers at the same rates as UCMSCs, while the expression of CD29 and CD105 in NPSCs were greatly down-regulated with IVD degeneration. These results indicated that trophic factors in UCMSCs-CM may stimulated D-NPSCs for self-repair, and regained healthier MSCs phenotype.
Similarly, the qRT-PCR results showed that the expression of Tie2 was significantly up-regulated after UCMSCs-CM treatment. Tie2, a cellular membrane receptor tyrosine kinase of the Tie family, was identified recently as a marker of NP precursor cells, which exhibited mulitpotency and self-renewal capacity (36). Notably, the expression of OCT4 and Nanog, the markers of pluripotency were also found greatly up-regulated in D-NPSCs after UCMSCs-CM treatment. Studies have shown that OCT4 and Nanog contributed to the stemness and multi-potency of human MSCs, and knockdown of OCT4 and Nanog in human MSCs reduced cell proliferation rate and differentiation potential (37, 38). These results indicated that UCMSCs-CM may improve the stemness and multi-potency of D-NPSCs.
It is noticeable that although the expression of Tie2 significantly up-regulated, while the expression of classic chondrogenic markers, SOX9, COL2A1 and ACAN, and NP cell specific marker CD24 were greatly down-regulated after UCMSCs-CM treatment. Moreover, the expression of OCT4 and Nanog were also significantly up-regulated following UCMSCs-CM treatment. Based on these results, we speculate that UCMSCs-CM improved the undifferentiated state and stemness of D-NPSCs, while reduced the expression of differentiated genes, such as SOX9, COL2A1 and ACAN. UCMSCs-CM may play an important role in dedifferentiation of the D-NPSCs, and consequently increase their multipotency.
Cell growth and proliferation assay fully confirmed that the proliferation capacity of D-NPSCs was remarkably improved by UCMSCs-CM, which was in alignment with the results of cell cycle and apoptosis analysis. As for multilineage differentiation potentials, the results demonstrated that UCMSCs-CM seemed to benefit osteogenesis and chondrogenesis more than adipogenesis in D-NPSCs. The exact underlying mechanism need to be further explored.
Our data demonstrated that UCMSCs-CM improved the stemness, proliferation capacity and potential for osteogenesis and chondrogenesis in D-NPSCs. Previous studies had also revealed that chemokines and growth factors secreted by MSCs were essential for promoting the regenerative capacity of other cells (39). Moreover, Song et al. (40) reported that adipose-derived stem cells improved the viability of nucleus pulposus cells from degenerated IVD in co-culture system. According to above studies, we speculate that CM derived from UCMSCs could promote the regenerative capacity of resident stem/progenitor cells from degenerated disc. The available data demonstrated that IVD degeneration involves not only a declining cell number, but a considerable functional impairment of NP cells. It is therefore evident that enhancing the functionality of NP resident/progenitor cells will be a crucial component in designing future regenerative treatment strategies.
In conclusion, Our finding demonstrated that CM derived from UCMSCs could promote the regenerative capacity of D-NPSCs by improving their stemness and proliferative capacity, and suggests the possibility of therapeutic strategies to rejuvenate IVD stem/progenitor cells via paracrine signaling, by soluble factors derived from MSCs. This study provides insights and perspectives for the development of a novel therapeutic strategy for IVD degeneration.
Supplementary Materials
Supplementary data including one table and one figure can be found with this article online at http://pdf.medrang.co.kr/paper/pdf/IJSC/IJSC-13-s20027.pdf.
 XML Download
XML Download