

 PDF
PDF Citation
Citation Print
Print


Introduction
Mesenchymal stem cells (MSCs) are pluripotent stem cells that can differentiate into many cell types, such as osteoblasts, chondrocytes, adipocytes, and muscle cells. In addition, it has been found in recent years that MSCs have immunomodulatory capacity and the potential to treat a variety of inflammation-related diseases (1-3). Currently, studies on immune regulation of T cells by MSCs have been extensively provided (1, 4, 5), but reports on the effects of MSCs on B cell function are rare and present conflicting data (2,6-11). Some studies have reported that MSCs can promote B cell proliferation, activation, differentiation and antibody secretion (7-9,12,13), while others have shown opposite effects (10,11,14-18). The main reason for this contradiction is that the immune regulation mechanism of MSCs on B cells remains unclear. Consequently, the use of MSCs for the effective treatment of certain immune diseases, such as systemic lupus erythematosus, multiple sclerosis and type 1 diabetes caused by abnormal B cells, is greatly hindered.
Suppressor of cytokine signaling 1 (SOCS1) is an important intracellular negative regulator of cytokine-induced signal transduction. SOCS1 encoding transcripts are usually present in cells at low levels, but their expression is rapidly induced by a variety of inflammatory cytokines, including IL-2, IL-3, IL-4, IL-6, IFN-γ and IFN-α (19-21). The up-regulated expression of SOCS1 protein down-regulates the signals induced by these cytokines in negative feedback loops (19, 21). Recent studies have shown that SOCS1 is involved in the formation and differentiation of various immune cells and plays an important role in the regulation of innate and adaptive immune responses (20-22).
Studies have shown that the concentration of inflammatory cytokines affects the immunomodulatory effects of MSCs on T cells (23-25). High concentrations of inflammatory factors can cause MSCs to exertan immunosuppressive effect, while insufficient levels of inflammatory factors can cause MSCs to play an immunological enhancement role (23). Zhang et al. (26) indicated that inflammatory factors can stimulate SOCS1 gene expression in MSCs. Currently, the role of SOCS1 in immune regulation of MSCs is still poorly understood. Zhang et al. (26) indicated that interference with SOCS1 expression in MSCs enhanced the immunosuppressive effect of MSCs on T cells. However, the role of SOCS1 in B cells immunomodulation by MSCs is unclear. Here, we found that, unlike its immunosuppressive effect on T cells, the SOCS1 gene knockdown of MSCs reversed the inhibition of B cell differentiation into plasma cells and promoted IgA+ B cell production.
Go to : 
Materials and Methods
Animals
6∼8 weeks-old male mice (C57BL/6) were purchased from the Laboratory Animal Center, Institute of Basic Medical Sciences, Beijing, China and maintained under specific pathogen-free conditions. The studies were approved by Animal Care and Use Committee of Tongren University.
Cells
The MSCs without (MSC/CTLsh) or with SOCS1 knockdown (MSC/SOCS1sh) are identical to those used in our previously published article (26). MSCs were cultured with α-MEM (Invitrogen) supplemented with 4 mM L-glutamine, 100 U/ml penicillin, 100 U/ml streptomycin and 10% fetal bovine serum (FBS) at 37℃ in a 5% CO2 incubator. B cells were separated from murine spleens with B220 MicroBead Kit (Miltenyi Biotec) as described previously (27).
Co-cultures
The cells were incubated in RPMI 1640 medium (Invitrogen) containing 10% FBS in the presence of different stimulators (group I: 10 μg/ml LPS; group II: 10 μg/ml LPS+25 ng/ml IL4; group III: 10 μg/ml LPS+1 ng/ml TGF-β). For the B cell proliferation assay, B cells were stained with CFSE and stimulated with LPS, LPS+IL4, LPS+TGFβ for 22 hours and then cultured alone or with different proportions (1:10, 1:20, 1:40, 1:80) of MSC/CTLsh or MSC/SOCS1sh. After co-culturing for 36 h, B cells were harvested for proliferation measurement using flow cytometry. For B cell activation assay, B cells were stimulated with LPS, LPS+IL4, LPS+TGFβ for 11 hours and then co-cultured by adding MSC/CTLsh or MSC/SOCS1sh at a ratio of 1:10. After 9 hours of co-culture, B cells were labeled with anti-CD40 and anti-CD86 antibodies and assayed for activation by flow cytometry. To evaluate the differentiation of B cells into plasma cells, B cells were stimulated with LPS, LPS+IL4, LPS+TGFβ for 23 hours and then co-cultured by adding 1:10 MSC/CTLsh or MSC/SOCS1sh. After two days of co-cultivation, B cells were collected for analysis of plasma cell formation. Finally, for IgA+ B cell formation assay, B cells were stimulated with LPS for 24 hours and then co-cultured for 3 days with MSC/CTLsh or MSC/SOCS1sh at a ratio of 1:10. The whole co-culture was stimulated with LPS. After 3 days of co-cultivation, B cells were collected for analysis of IgA+ B cells.
Monoclonal antibodies and FACS analysis
The antibodies used were: monoclonal Abs conjugated to APC: -anti-CD220, -anti-CD86; Biotin-anti-IgA; and PE: -anti-CD40, -anti-CD138 (all from BD Biosciences). Cells labeled with biotinylated antibodies were visualized by incubation with Phycoerthyrin (PE) conjugated streptavidin. For cell proliferation assays, B cells were labeled with carboxyfluorescein diacetate, succinimidyl ester (CFSE, Invitrogen) as described previously (27). Data were collected at FACS Calibur (Becton Dickinson, San Jose, CA, USA) and analyzed with FlowJo software (TreeStar).
Prostaglandins E2 (PGE2) determination
MSC/SOCS1sh and MSC/CTLsh were incubated with different doses (0, 0.5 and 5 ng/ml) of TNF-α/IFN-γ at 37℃, 5% CO2 for 12 h. Supernatants were collected and precipitated by centrifugation to remove cell debris, and the remaining supernatants were used for PGE2 determination. PGE2 measurement was performed by ELISA (Cayman Chemical, Ann Arbor, MI) according to the manufacturer’s instructions. All samples were measured in triplicate.
Statistical analysis
Statistical analyzes were performed by unpaired two-tailed Student’s t-test and one-way analysis of variance (ANOVA) followed by Tukey’s multiple comparison post-test. p values<0.05 were considered statistically significant.
Go to :
Results
SOCS1 knockdown enhances the inhibitory effect of MSCs on B cell proliferation
To verify the effect of MSCs on B cell proliferation and the role of SOCS1 therein, B cells were co-cultured with different ratios of MSCs (1:10, 1:20, 1:40 and 1:80) under the stimulation of LPS (Fig. 1A), LPS+IL4 (Fig. 1B) and LPS+TGFβ (Fig. 1C). The results showed that under the stimulation of the above three groups of different cytokines, regardless of the MSC/CTLsh or MSC/SOCS1sh group, the B cell proliferation rate was inversely proportional to the amount of MSCs added. When the ratio of MSCs to B cells was 1:10, B cell proliferation was the lowest of all MSCs ratios, regardless of the MSC/CTLsh group or the MSC/SOCS1sh group. Besides, in this MSCs/B cells ratio, the B cell proliferation was significantly less than that of the isolated B cell group. On the other hand, when the ratio of MSCs to B cells was reduced to a minimum of 1:80, B cell proliferation rate was the highest of all ratios. Moreover, the B cell proliferation rate was also higher than that of the isolated B cell group. For both LPS (Fig. 1A) and LPS/TGFβ (Fig. 1C) cytokine-stimulated groups, B cell proliferation in the MSC/SOCS1sh group was significantly lower than the MSC/CTLsh at a ratio of 1:10. This difference between the two groups was gradually reduced in 1:40 and 1:80 ratios. These results indicated that MSCs regulate B cell proliferation in a dose-dependent manner. Low doses of MSCs showed an immune-promoting effect, while high doses of MSCs exerted an immunosuppressive effect. Moreover, the immunosuppression was more pronounced in the MSC/SOCS1sh group.
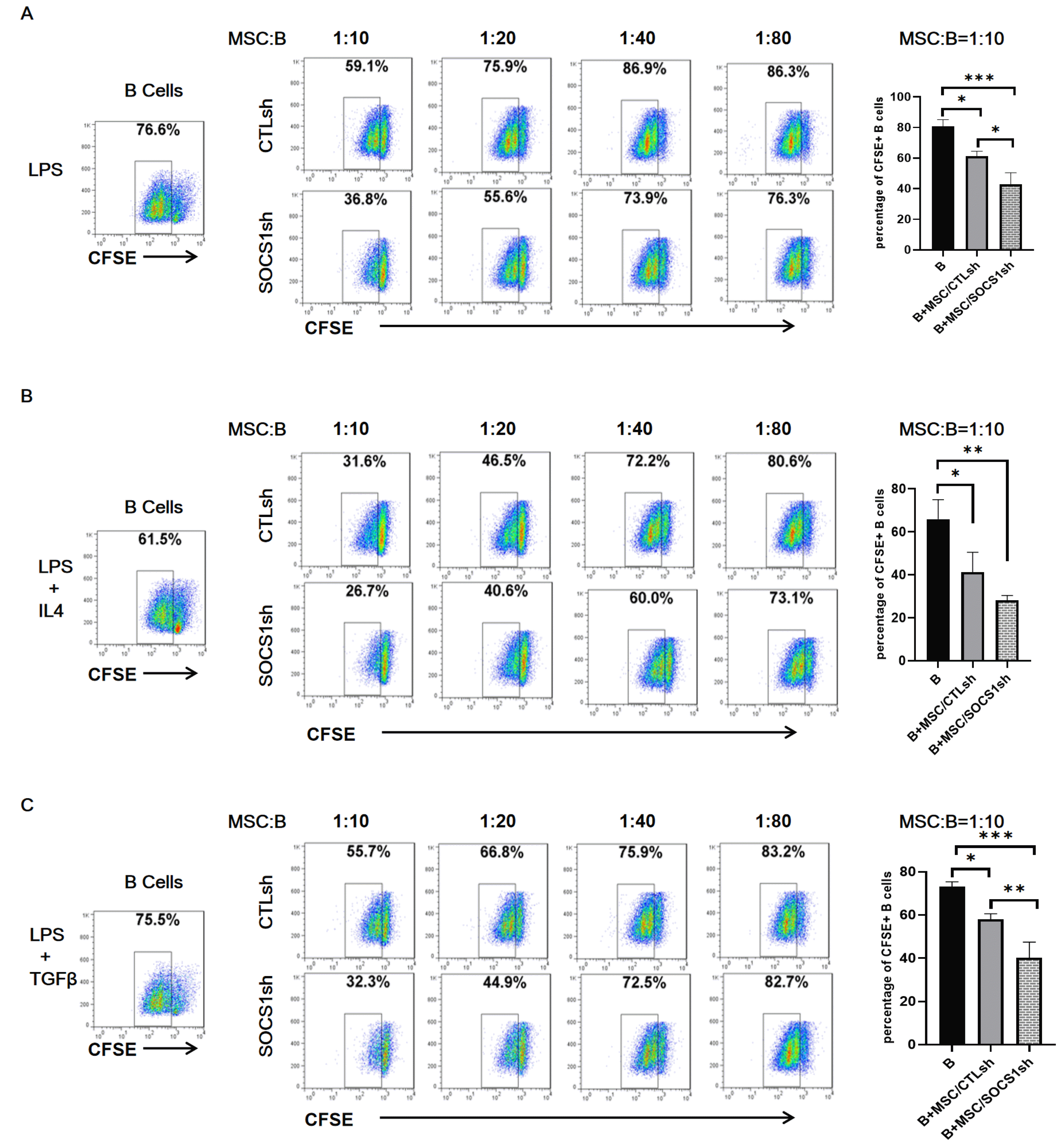 | Fig. 1SOCS1 knockdown aggravates the inhibitory effect of MSCs on B cell proliferation. The B cells isolated from the mouse spleens were labeled with CFSE and stimulated with LPS (A), LPS+IL4 (B), LPS+TGFβ (C) for 22 hours and then cultured alone or co-cultured with different ratios of MSC/CTLsh or MSC/SOCS1sh under the stimulation of the above cytokines. After 36 hours of co-culture, the B cell proliferation was analyzed by flow cytometry. Data were representative of three independent experiments. *p<0.05, **p<0.01, ***p<0.001. Note: CTLsh, MSC/CTLsh; SOCS1sh, MSC/SOCS1sh.
|
SOCS1 downregulation has no effect on inhibition of B cell activation by MSCs
We evaluated the expression of CD40 and CD86 activation markers in B cells to further reveal the role of SOCS1 from MSCs in B cell activation. B cells were co-cultured with MSC/CTLsh or MSC/SOCS1sh under the stimulation of LPS (Fig. 2A), LPS/IL4 (Fig. 2B) and LPS/TGFβ (Fig. 2C). The results showed that the percentage of CD40+ and CD86+ B cell in MSC/CTLsh group was generally lower than in the isolated B cell groups in any of the cytokine-stimulated groups. However, there were no differences between MSC/CTLsh and MSC/SOCS1sh groups. These results indicated that MSCs inhibited B cell activation and that SOCS1 knockdown had no influence on this effect.
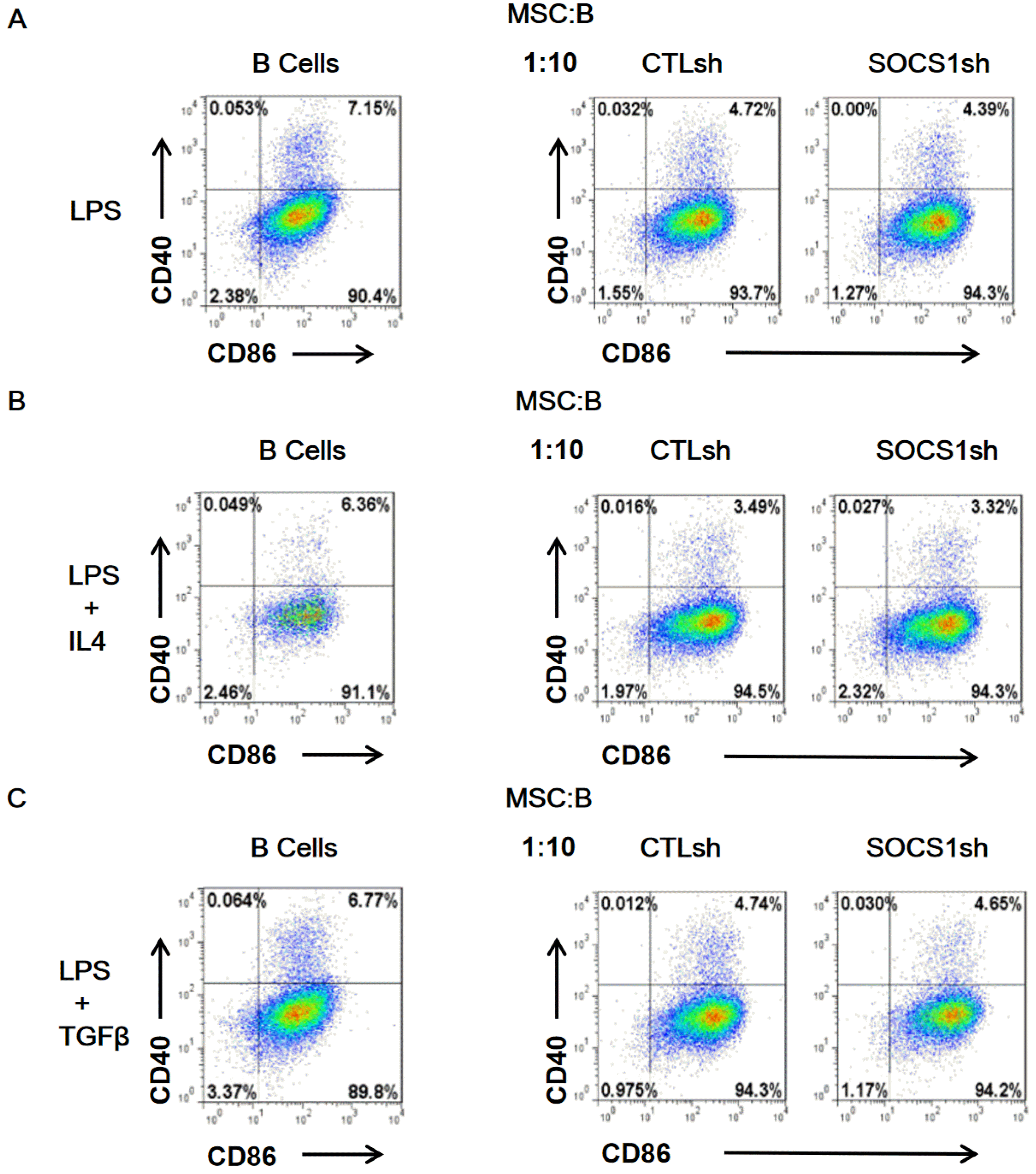 | Fig. 2SOCS1sh has no effect on the inhibition of B cell activation by MSCs. B cells isolated from mouse spleens were stimulated with LPS (A), LPS/IL4 (B) and LPS/TGFβ (C) for 11 hours and then co-cultured by adding MSC/CTLsh or MSC/SOCS1sh at a ratio of 1:10. After 9 hours of co-culture, B cells were labeled with anti-CD40 and anti-CD86 antibodies. B cell activation was analyzed by flow cytometry. Note: CTLsh, MSC/CTLsh; SOCS1sh, MSC/SOCS1sh.
|
SOCS1 knockdown reverses the inhibition of B cell differentiation into plasma cells by MSCs
To further clarify the effect of MSCs on B cell differentiation, B cells were cultured with MSCs at a ratio of 1:10 under stimulation of LPS (Fig. 3A), LPS/IL4 (Fig. 3B) and LPS/TGFβ (Fig. 3C). The results showed that, compared to the isolated B cells group, the MSC/CTLsh co-culture inhibited the expression of the plasma cell marker CD138, regardless of the cytokine-stimulated group. Remarkably, MSC/SOCS1sh has abolished the inhibition of CD138 expression induced by MSC/CTLsh. These results indicated that SOCS1 knockdown could reverse the inhibitory effect of MSCs in the last stage of B cell differentiation.
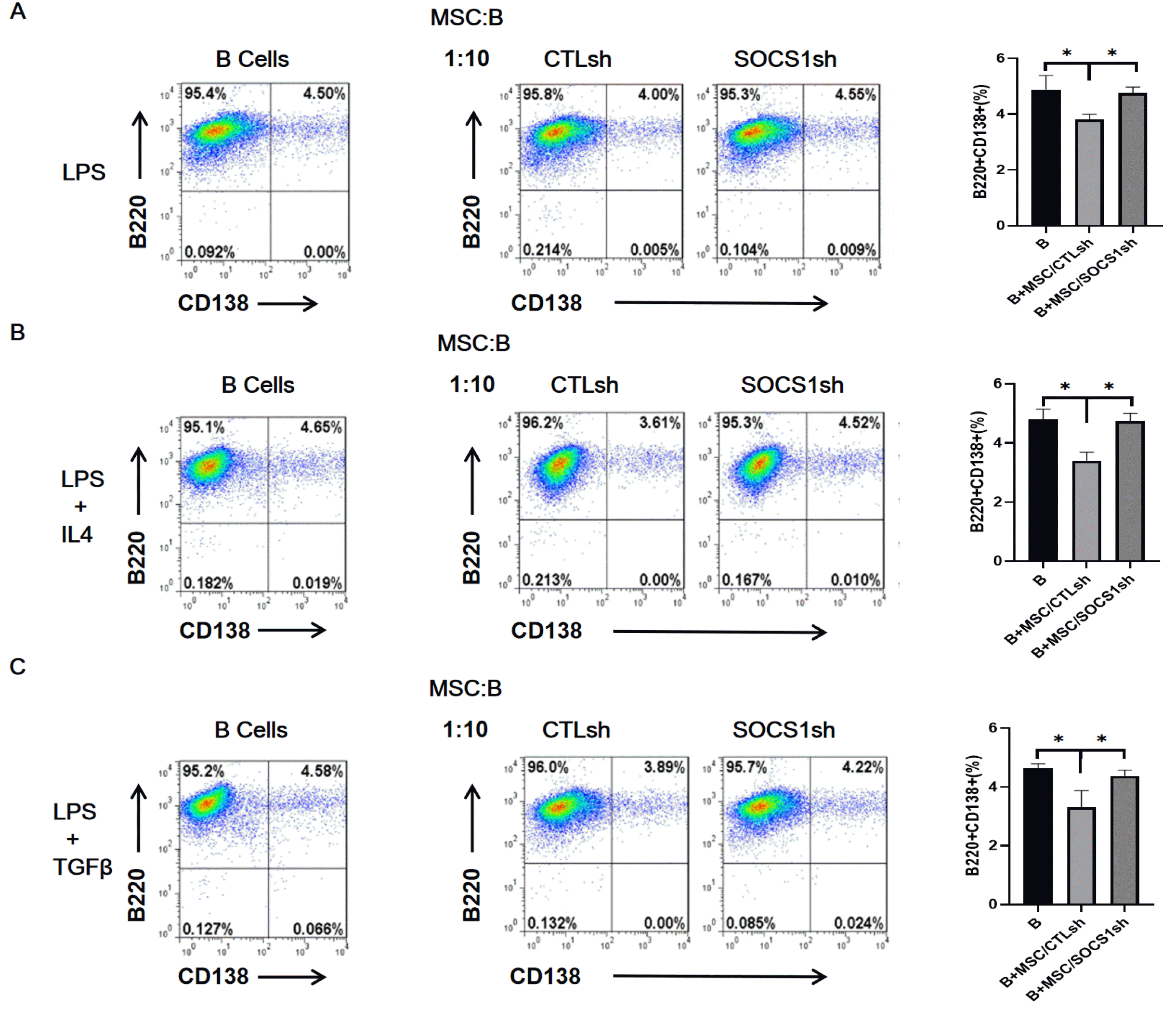 | Fig. 3Knockdown of SOCS1 reverses the inhibition of plasma cell formation by MSCs. B cells were stimulated with LPS (A), LPS/IL4 (B), LPS/TGFβ (C) for 23 hours and then co-cultured by adding MSC/CTLsh or MSC/SOCS1sh at a ratio of 1:10. After two days of co-cultivation, B cells were harvested, labeled with anti-B220 and anti-CD138 antibodies and analyzed for plasma cell formation by flow cytometry. Data were representative of three independent experiments. *p<0.05. Note: CTLsh, MSC/CTLsh; SOCS1sh, MSC/SOCS1sh.
|
MSCs promoted IgA+ B cell formation after SOCS1 knockdown
IgA is the most abundant immunoglobulin in the human body and has an irreplaceable role in mucosal immunity (28). More than 80% of plasma cells produce IgA in mucosa-associated lymphoid tissues (28). The effect of MSCs on IgA+ B cells production is still unclear. Here, we examined the formation of IgA+ B cells co-cultured with MSC/CTLsh or MSC/SOCS1sh (Fig. 4). We found no significant differences in IgA+ B cell production in these two groups compared to the isolated B cells group. However, we showed that the IgA+ B cell production was significantly higher in the MSC/SOCS1sh than in the MSC/CTLsh group (Fig. 4). These results indicated that MSCs promoted the production of IgA+ B cells after SOCS1 knockdown.
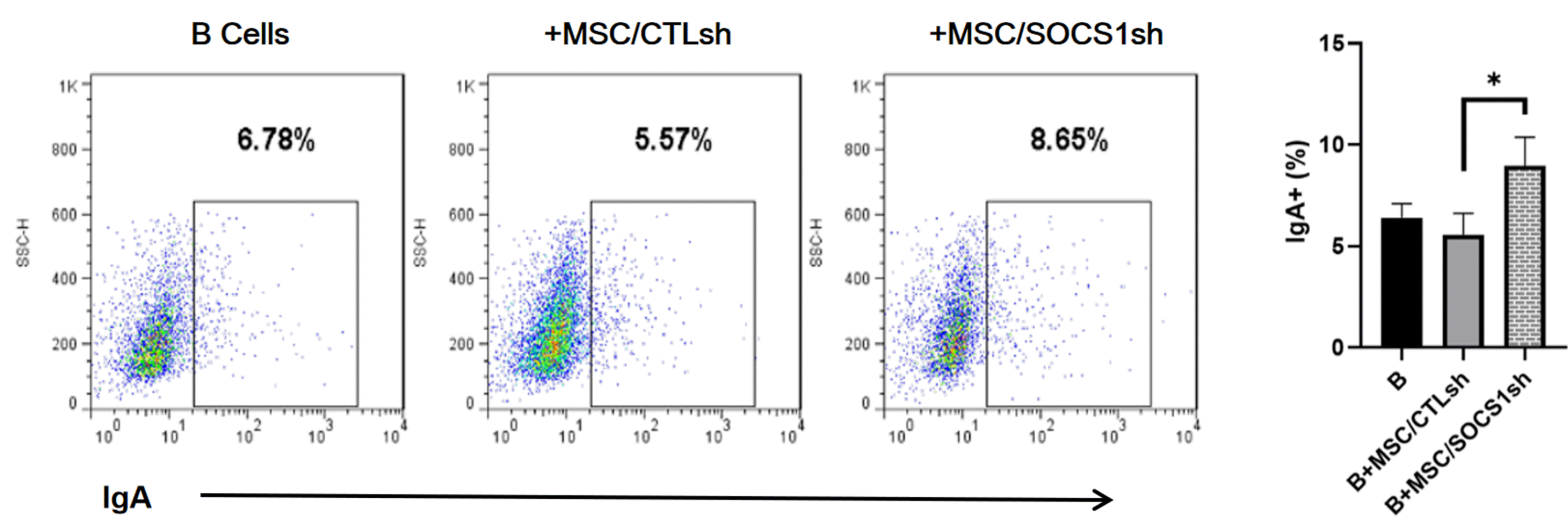 | Fig. 4MSC/SOCS1sh promotes IgA+ B cell formation. Purified B cells were stimulated with LPS for 24 hours, then MSC/CTLsh or MSC/SOCS1sh was added at a ratio of 1:10 and co-cultured for 3 days. All cultures were stimulated with LPS. After co-cultivation for 3 days, the percentages of IgA+ B cells were determined by FACS, which are showed within the squares. Data were representative of three independent experiments. *p<0.05. Note: CTLsh, MSC/CTLsh; SOCS1sh, MSC/SOCS1sh.
|
SOCS1 knockdown increases PGE2 secretion of MSCs
We evaluated PGE2 secretion in MSCs stimulated by TNF-α/IFN-γ inflammatory factors (Fig. 5), because PGE2 has been reported to be involved in B cell development (29). The results showed that the secretion of PGE2 increased in the MSC/CTLsh and MSC/SOCS1sh groups after TNF-α/IFN-γ stimulation. Besides, the PGE2 production in MSC/SOCS1sh was significantly higher than in the MSC/CTLsh group (Fig. 5). These results indicated that SOCS1 knockdown can promote PGE2 production in MSCs.
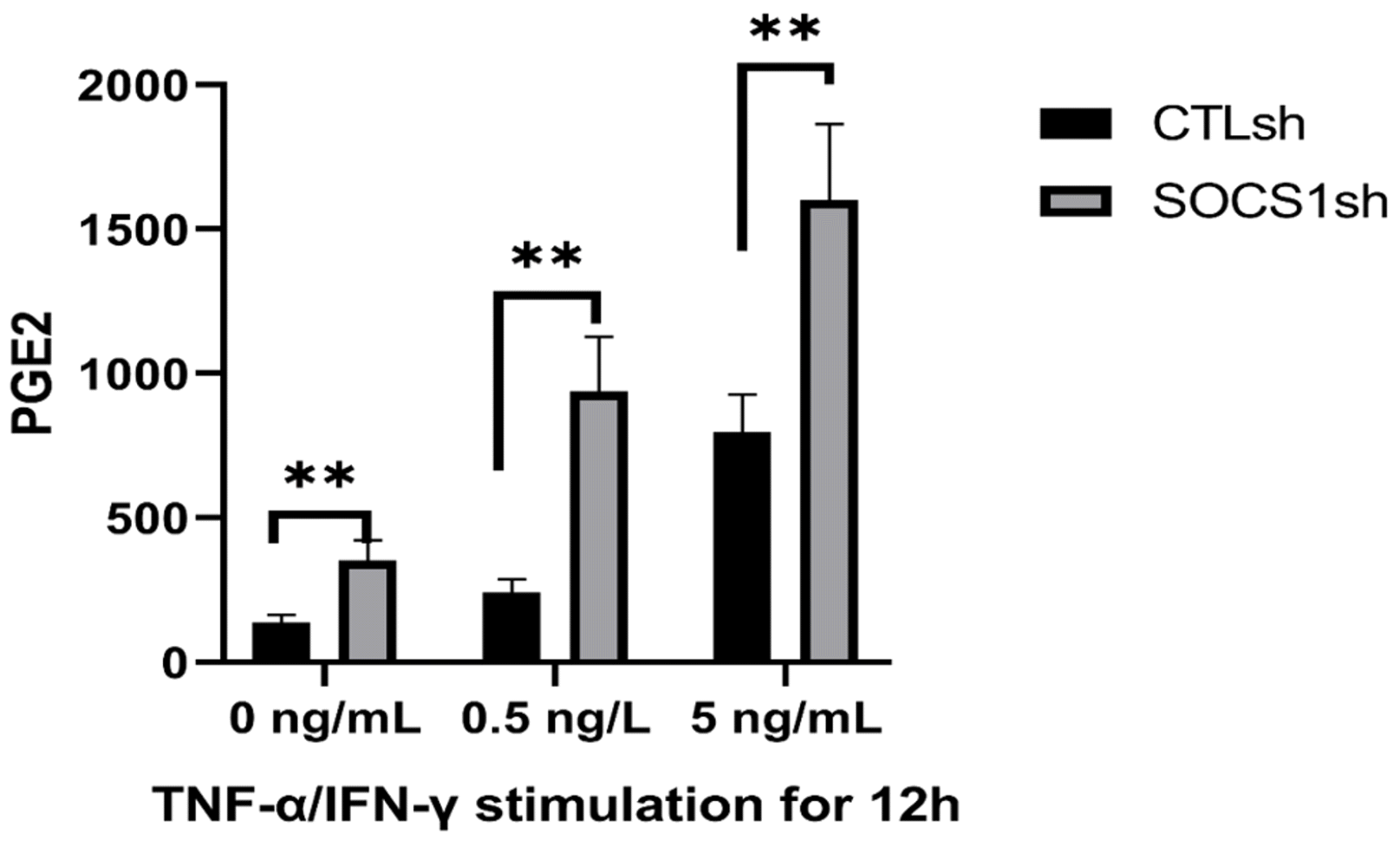 | Fig. 5SOCS1 knockdown promotes PGE2 secretion by MSCs. MSC/CTLsh or MSC/SOCS1sh were stimulated with different doses (0, 0.5 and 5 ng/ml) of TNF-α/IFN-γ for 12 h and then the level of PGE2 in the culture medium was measured by ELISA. Data were representative of three independent experiments. **p<0.01. Note: CTLsh, MSC/CTLsh; SOCS1sh, MSC/SOCS1sh.
|
Go to :
Discussion
MSCs have great potential for application in the treatment of certain immune-injured diseases due to their immunomodulatory properties and damage repair ability (1). However, the unstable immunomodulatory effects of MSCs on B cells and an inadequate understanding of the underlying mechanisms limit their use in the treatment of diseases associated with abnormal B cells. In the present work, by co-culture assays of MSCs and B cells, we found that MSCs inhibited B cell proliferation and differentiation and that this inhibitory effect could be regulated by SOCS1 from MSCs.
The few reported studies on B cells immune regulation by MSCs were not sufficient to define the effects of MSCs on B cell function, as they present conflicting data. Here, our results indicated that MSCs inhibited B cell function, which was consistent with some previous reports (10, 11, 15, 16, 18).
The immune regulation of T cells by MSCs is related to the concentration of inflammatory factors. Previous studies have shown that excessive inflammatory factors can induce MSCs to inhibit T cell responses (23, 30). However, the effects of these factors on B cell function are poorly understood. MSCs stimulated by the inflammatory factor IFN-γhave been reported to enhance the inhibition of B cell proliferation (12). Moreover, it is known that immune cells can secrete inflammatory factors including TNF-α, IFN-γand ILb in response to stimuli provided by LPS, TGF-β and ConA. Our experiments showed that differences in co-culture cultivation affects the immunomodulatory effects of MSCs. When MSCs were implanted before B cells addition or immediately after B cells implantation, MSCs promoted, but did not inhibit B cell proliferation (data not shown). On the other hand, when B cells were added 10 h after stimulation of MSCs by TNF-α/IFN-γ (data not shown) or when MSCs were co-cultured 20 h after B cells implantation, the proliferation of B cells was completely inhibited (Fig. 1). Therefore, the mode of planting MSCs or B cells probably affects the concentration of inflammatory factors in the co-culture. When B cells receive stimulating factors for a long period (for example, up to 20 hours), the concentration of inflammatory factors secreted by them in the medium may be high. If MSCs are added at this time, MSCs will be induced to exert an immunosuppressive effect. In contrast, whether MSCs are seeded before adding B cells or the implantation interval between B cells and MSCs is short, the inflammatory factors in the culture medium are insufficient to trigger the immunosuppressive effect of MSCs. However, even at lower concentrations, inflammatory factors are still able to induce the immune promoting effect of MSCs. Based on these observations, it can be concluded that the different immunomodulatory effects of MSCs on B cells may be highly correlated with the different degrees of inflammation. Moreover, these inflammation degrees are the result of different methods of planting MSCs or B cells. For the first time, it is reported that the sequence of implantation of MSCs and B cells in co-cultures affects the regulation of B cells by MSCs.
Inflammatory factors are able to induce SOCS1 expression and can be regulated by its negative feedback (20, 21). Zhang et al. (26) showed that inflammatory factors trigger SOCS1 expression in MSCs, while SOCS1 gene knockdown enhances MSCs to inhibit T cell proliferation. In the present study, our data indicated that SOCS1 gene knockdown in MSCs also increased the inhibitory effect of MSCs on B cell proliferation. In addition, we showed that MSC/CTLsh and MSC/SOCS1sh regulated of B cell proliferation in a dose-dependent manner. That is, as the dose of MSCs increases, the regulation of B cell proliferation by MSCs is gradually shifted from a promoting effect to an inhibitory one. The dose-dependent regulation of MSCs observed in our experiments is consistent with a previous study (11). This effect may be due to the fact that certain factors secreted by MSCs accumulate to a certain extent (threshold) to inhibit B cell proliferation, otherwise they promote B cell proliferation.
Some studies (1, 31, 32) have shown that MSCs can secrete PGE2 cytokines to inhibit T cell responses. Although PGE2 has been reported to be involved in B cell development (29), the role of PGE2 in the regulation of B cells by MSCs remains unknown. Our results indicated that SOCS1 knockdown in MSCs could increase PGE2 production. Downregulation of SOCS1 abolished the inhibitory effect of MSCs on B cell differentiation and promoted IgA+ B cell formation, which may be related to PGE2 and nitric oxide (NO) overproduction. PGE2 has been reported to promote IgA formation (33) and that NO production promotes the generation of IgA+ plasma cells (28, 34). Zhang et al. (26) have shown that MSC/SOCS1sh inhibited T cell proliferation by promoting NO production. In the present study, the increase in IgA+ B cells in the MSC/SOCS1sh group may be related to the contribution of NO in addition to PGE2. Although MSCs also produce NO and PGE2, co-culture with B cells does not promote the formation of IgA+ B cells. This can be related to other MSC-derived factors that suppress IgA+ B cell formation. Following SOCS1 gene knockdown in MSCs, the inhibitory effect of other factors on IgA production may be masked by the large amount of NO and PGE2 that promote IgA production. Remarkably, the specific mechanism by which SOCS1 regulates the interaction between MSCs and B cells remains to be further studied.
In conclusion, we found that the different immunoregulatory effects of MSCs on B cells are related to differences in co-culture cultivation. Furthermore, we revealed a novel role of SOCS1 in regulating B cell function by MSCs. These results provided new insights into B-cell immunomodulation by MSCs that may help to develop new treatments based on the use of MSCs for B cell mediated immune diseases.
Go to :
 XML Download
XML Download