

 PDF
PDF Citation
Citation Print
Print


Introduction
Embryonic stem (ES) cells undergo self-renewal and can be differentiated into three germ layers: ectoderm, mesoderm, and endoderm. Thus, ES cells have been considered as useful tools for both research and regenerative medicine (1, 2). The regulators of ES cell self-renewal have been identified. Leukemia inhibitory factor (LIF), the factor secreted by feeder cells, is necessary for self-renewal of mouse ES cells in feeder cell-free condition (3), but not in human ES cells (1, 2, 4). Knock-out experiments showed that Oct4 (5), Sox2 (6), and Nanog (7, 8) transcription factors are required for self-renewal of ES cells. While increasing Oct4 expression induced differentiation of endoderm (9), the overexpression of Nanog was sufficient to sustain ES cell self-renewal in the absence of LIF (7, 8).
Wnt/β-catenin signaling has been shown to play an important role in various stem and progenitor cells (10, 11). In ES cells, the aberrant activation of Wnt/β-catenin signaling maintains stemness of ES cells by blocking cell differentiation. While ectopic expression of secreted frizzled related protein 2 (sFRP2), an antagonist of Wnt signaling, induces enhanced neural differentiation, Wnt1 overexpression prevents neural differentiation (12). Stabilization of β-catenin by adenomatous polyposis coli (APC) mutation or ectopic expression of a dominant active form of β-catenin resulted in inhibition of differentiation (13, 14). Furthermore, the activation of Wnt signaling by a pharmacological glycogen synthase kinase-3 (GSK3)-specific inhibitor leads to maintenance of self-renewal in human and mouse ES cells (15, 16). However, evidence supporting Wnt signaling in ES cells is disputed. The β-catenin-mediated TOP activity in undifferentiated human ES (hES) cells is lower than in differentiated cells and treatment of Wnt3a leads to enhanced differentiation of hES cells in the absence of an anti-differentiation factor (17). In addition, canonical Wnt signaling regulates differentiation into mesodermal lineages or neural precursors (18-22). This contradictory response of ES cells upon Wnt signaling suggests the need for further studies to understand the regulation of ES cells by Wnt signaling.
Stemness and differentiation of ES cells are regulated by a number of transcription factors. Because (T-cell factors) Tcfs and (lymphoid enhancer factor 1) Lef1 are transcription factors transmitting Wnt signal, it is possible that Tcfs and Lef1 differentially regulate self-renewal or differentiation of ES cells upon Wnt signaling. The Tcfs/Lef1 family is composed of four genes in vertebrates (Tcf1, Lef1, Tcf3, and Tcf4) (23). Tcfs/Lef1 consists of four domains in common: a β-catenin binding domain, a central domain, a HMG DNA-binding domain, and a C-terminal tail. A variety of Tcfs/Lef1isoforms are generated by alternative splicing and promoter usage (24, 25). At low levels of stabilized β-catenin, Tcfs/Lef1 proteins act as transcriptional repressors along with corepressor proteins, such as Groucho, C-terminal binding protein (CtBP), hypermethylated in cancer 1 (HIC1), and hypermethylated in cancer 5 (HIC5). In the presence of Wnt stimulation, however, Tcfs and Lef1 act as transcriptional activators when β-catenin displaces Groucho (24, 26). Knockout studies of Tcfs and Lef1 genes showed that Tcfs and Lef1 exhibit redundant and non-redundant functions in embryonic development and adult stem cell regulation (25). These diverse functions of Tcfs and Lef1 suggest important roles in Wnt signaling in ES cells. Although it has been known that Tcf3 acts as a limiting factor in self-renewal of ES cells (11,27-31), the role of other Tcfs and Lef1 in ES cells remains unknown.
In this study, we show that the expression of Tcfs and Lef1 proteins in ES cells changes dynamically and the differential pattern under the three conditions of differentiation suggests that each Tcfs and Lef1 may act differently in ES cells. Ectopic expression of Tcf1 contributes to their self-renewal and inhibits their differentiation even in the absence of LIF by sustaining the level of Nanog. In contrast, the overexpression of Lef1 does not result in self-renewal and knockdown of Lef1 inhibits differentiation. Overall, our data suggest that each Tcfs and Lef1 has a specific role in the maintenance of stemness and differentiation of ES cells.
Materials and Methods
Culture and differentiation of mouse ES cells
A6P10 mES cells (a gift from Dr. Chyuan-Sheng Lin, Columbia University, USA) and 46C mES cells (ES cell line in which EGFP was replaced into the open reading frame of Sox1 gene, provided by Dr. Qilong Ying, University of Southern California, USA) were cultured in ES medium (DMEM (Gibco) with 15% FBS, 2 mM GlutaMAX (Gibco), MEM nonessential amino acids, β-mercaptoethanol (Gibco), tylosin, 1% Pen/Strep (Gibco)) supplemented with LIF (Chemicon) on 0.2% gelatin-coated dishes. To induce neuronal differentiation, 46C cells were cultured in N2B27 medium (DMEM/F12 (Gibco), Neurobasal medium (Gibco), N2 supplement (Invitrogen), B27 supplement (Invitrogen), 1 mM GlutaMAX (Gibco), 0.1 μM β-mercaptoethanol (Gibco), 1% Pen/Strep (Gibco)) on 0.2% gelatin-coated tissue culture dish (Falcon). N2B27 medium was changed every 2 days. Embryoid body (EB) formation was induced by hanging drop method. Briefly, 20 μl drops (including 600 cells) of dissociated ES cells with ES medium plus 20% FBS were placed on inverted lids of petri-dish (Falcon), which was filled with 3 ml PBS. After incubation for 3 days, EB was plated on a 0.2% gelatin-coated dish in ES medium supplemented with 20% FBS. The medium was changed every 2 days.
Plasmids and transfection
RNA obtained from a mixture of undifferentiated and differentiated mES cells was used to clone Tcf1, Lef1, Tcf3, and Tcf4. Wild-type and dominant negative forms of Tcfs/Lef1 were inserted into the pCS2-HA3 vector. HA-tagged Tcfs and Lef1 were transferred into the pCAG-1 vector (modified from pPCAGIZ vector). The shRNA targeting sequences against mouse Lef1 were designed using the web tool from Promega.
Sense (5’-GATCCCCGACTTAGCCGACATCAAGTTTCA AGAGAACTTGATGTCGGCTAAGTCTTTTTGGAAA-3’) and antisense (5’-AGCTTTTCCAAAAAGACTTAGCCGA CATCAAGTTCTCTTGAAACTTGATGTCGGCTAAGTCGGG-3’) oligonucleotides were annealed and ligated into the BglII and HindIII sites of the pSUPER.retro.puro vector (Oligoengine). HA-tagged Tcfs and Lef1 plasmids were electroporated by Amaxa Nucleofector according to the manufacturer’s protocol and then selected with 50 μg/ml Zeocin (Invitrogen). The shLef1 plasmid was electroporated by Amaxa Nucleofector technologyTM and selected with 1 μg/ml puromycin (Sigma).
Western blotting and antibodies
ES cells were lysed in lysis buffer (20 mM Tris-HCl, pH 7.5, 150 mM NaCl, 0.5% Triton X-100, 50 mM NaF, 2 mM EDTA, 100 μM Na-orthovanadate, 1 mM PMSF, 5 μg/ml leupeptin, and 1 μM pepstatin A). The lysates were centrifuged at 13,000 rpm for 15 min at 4℃ and the supernatant was collected and used for Western blotting. Bradford (Bio-Rad) reagent was used to measure the quantity of protein. Equal amounts of protein were boiled in Laemmli sample buffer and resolved via SDS-PAGE followed by transfer to a PVDF membrane (Pall). Anti-β-actin (Sigma), anti-TCF1 (Cell Signaling), anti-LEF1 (Cell Signaling or Santa Cruz Biotechnology), anti-TCF3 (Santa Cruz Biotechnology), anti-TCF4 (Santa Cruz Biotechnology) antibodies were used as primary antibodies.
Alkaline phosphatase (AP) staining
ES cells were plated layered on a 12-well plate and cultured with or without LIF. After washing twice with PBS, cells were fixed with 4% paraformaldehyde for 10 min at room temperature followed by PBS washing several times. AP staining was performed with NBT/BCIP (4-nitro blue tetrazolium chloride, Roche; 5-Bromo4-chloro-3-indolyl-phosphate, Roche) staining buffer (0.1 M Tris, pH 9.5, 100 mM NaCl, 5 mM MgCl2) for 15 min in the dark.
Chromatin immunoprecipitation (ChIP) assay
Cells were cross-linked with 1% formaldehyde (Sigma) at room temperature for 10 min with gentle shaking and then incubated in 0.125 M glycine for 5 min with gentle shaking. Cells were washed twice with ice-cold PBS before harvest. Re-suspended cells with hypotonic buffer (10 mM Hepes-KOH, pH 7.8, 10 mM KCl, 1.5 mM MgCl2) were swollen on ice for 10 min and passed through a 26.5-gauge needle 6 times. After centrifugation at 1000 g for 5 min at 4℃, the pellets were incubated in nuclei lysis buffer (1% SDS, 50 mM Tris-HCl, pH 8.0, 10 mM EDTA) for 10 min on ice with occasional vortexing. Chromatin was sheared to an average length of 0.2∼1 kb by sonication on ice. Cellular debris was removed by centrifugation (13,000 rpm, 15 min, 4℃), and the concentration of supernatant was determined with a spectrophotometer. The appropriate volume of chromatin was diluted 1/10 in ChIP dilution buffer (0.01% SDS, 20 mM Tris-HCl, pH 8.0, 167 mM NaCl, 1.2 mM EDTA, 1.1% Triton X-100) followed by preclearance at 4℃ for 2 h with 10 μl protein A/G plus-agarose beads (Santa Cruz Biotechnology). For immunoprecipitation, goat, rabbit-IgG (Bethyl), anti-Lef1 (Santa Cruz Biotechnology or Cell Signaling), anti-Tcf3 (Santa Cruz Biotechnology), anti-Tcf1 (Cell Signaling) antibodies were used at 4℃ overnight. Immunoprecipitated chromatins were eluted, and then reverse cross-linked by the addition of 0.3 M NaCl at 65℃ overnight. Following phenol-chloroform extraction and ethanol precipitation, the DNA was dissolved in 50 μl TE buffer (10 mM Tris-HCl, pH 8.0, 1 mM EDTA). For PCR, 2 μl DNA was used. A Nanog primer (forward: 5’-TCTGCTTATACACA GAAGCC-3’ and reverse: 5’-AAGTGCCTCAGCCGTCTA AG-3’) was used for PCR.
Reverse transcriptase-PCR (RT-PCR)
Total mRNA from ES cells were isolated by TRIzol (Invitrogen) according to manufacturer protocol. cDNA was made from total RNA using reverse transcriptase (Promega) with random primers. For PCR, taq polymerase (Cosmo genetech) was used and visualized by 1% of agarose gel. GAPDH or 18s rRNA were used for normalization of mRNA expressions. Primers used for RT-PCR can be found in Supplementary Table S1.
Results
Differential expression of Tcfs and Lef1
Wnt signal stimulates the self-renewal and differentiation of ES cells into various lineages. We investigated the varying response of ES cells in the presence of Wnt signal. As Tcfs and Lef1 act as the downstream effectors in Wnt signal transmission, we speculated that the differential response may result from differential function of each Tcfs and Lef1.
Prior to investigation of the function of Tcfs and Lef1, the change in Tcfs and Lef1 mRNA levels during differentiation was detected by RT PCR (Fig. 1A). Under all conditions of differentiation, the Tcf3 transcript was found to be increased during the early days of differentiation. This data is consistent with previous reports that TCF3 is involved in the early stage of differentiation by suppressing pluripotent genes (32). In addition, Tcf4 transcript was also gradually increased during differentiation. Interestingly, Lef1 transcript was transiently induced after the removal of LIF although this induction was maintained in N2B27 media. There was no significant change in Tcf1 expression following LIF depletion and N2B27. However, Tcf1 transcript decreased from day 6 post-EB formation. These results suggest that Lef1 might play a role in the initial stages of differentiation and Tcf4 might be associated with late differentiation.
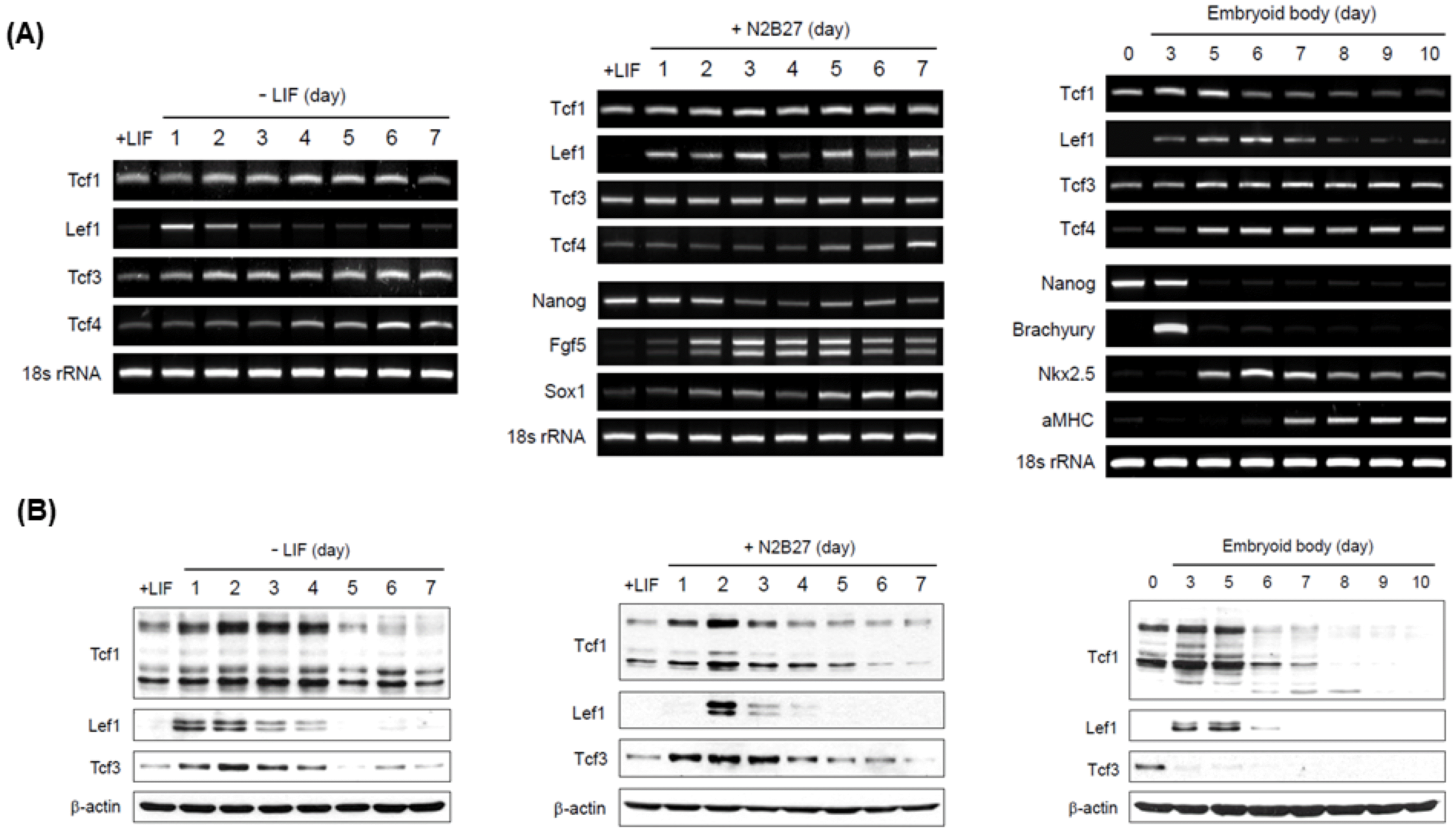
Fig. 1
The pattern of Tcfs and Lef1 expression in ES cells. (A) A6P10 ES cells were differentiated in the absence of LIF (left), in N2B27 medium without LIF (middle), or via EB formation without LIF (right). The differentiated ES cells were harvested on the days indicated. The amount of Tcfs and Lef1 transcripts was measured by RT-PCR analysis. The bands for Tcfs and Lef1 represent wild-type transcripts including β-catenin-binding domain. Self-renewal marker (Nanog), neural precursor markers (Sox1 and Fgf5) for N2B27 differentiation and mesodermal markers (Brachyury, Nkx2.5, αMHC) for EB differentiation were used to confirm proper differentiation; 18s rRNA was used as a loading control. (B) The endogenous protein levels of Tcfs/Lef1 were detected by western blotting. β-actin was used as a loading control.
![]()
It has been known that the function of Tcfs/Lef1 is affected by post-translational modifications such as phosphorylation and sumoylation (26). Thus, we performed western blotting analysis to investigate the protein levels of Tcfs and Lef1 (Fig. 1B). The protein levels of Tcfs and Lef1 were dynamically changed compared to the changes in Tcfs and Lef1 transcripts. Similar to RT PCR data, Lef1 protein was expressed transiently at an early stage of differentiation. In contrast, the expression of Tcf1 and Tcf3 proteins revealed different patterns of mRNA levels. Under the conditions of LIF depletion or N2B27 medium, the levels of Tcf1 and Tcf3 protein were elevated until the day Lef1 protein was expressed followed by a sharp decline during subsequent differentiation. The expression pattern of Tcf1 protein during EB formation was similar to that under other conditions, whereas the pattern of Tcf3 expression differed completely. After EB formation for 3 days, the loss of Tcf3 protein was inconsistent with RT PCR data, suggesting that the Tcf3 protein might be degraded by unknown mechanisms during EB formation. These results suggest that measurement of protein levels might be more appropriate for Tcfs/Lef1 studies in ES cells. The variable expression of Tcfs and Lef1 proteins under differentiated and undifferentiated conditions supports our hypothesis that the role of each Tcfs and Lef1 may differ in self-renewal and differentiation of ES cells.
Ectopic expression of Tcf1 maintains self-renewal of ES cells in the absence of LIF
To investigate the functions of Tcfs and Lef1 proteins, Tcfs and Lef1 constructs cloned from RNA expressed in ES cells were stably introduced into ES cells (Fig. 2A). First, we tested which Tcfs and Lef1 proteins regulated the stemness of ES cells using alkaline phosphatase (AP), a marker of self-renewal, staining (Fig. 2B and 2C). When LIF was removed from media, AP positive cells (AP+) were remarkably reduced and the morphology was lost in ES cells expressing empty vector. Likewise, AP+ decreased steadily in ES cells expressing Lef1, Tcf3, or Tcf4 (Fig. 2C). In contrast, AP+ was retained and the morphology remained intact in ES cells expressing Tcf1 but not dominant-negative Tcf1 (TCF1-DN) even after the removal of LIF for 12 days (Fig. 2B). Moreover, ES cells expressing Tcf1 were still stained with the antibody against SSEA1, a marker for proliferation (data not shown). These results suggested that Tcf1 may regulate the self-renewal of ES cells.
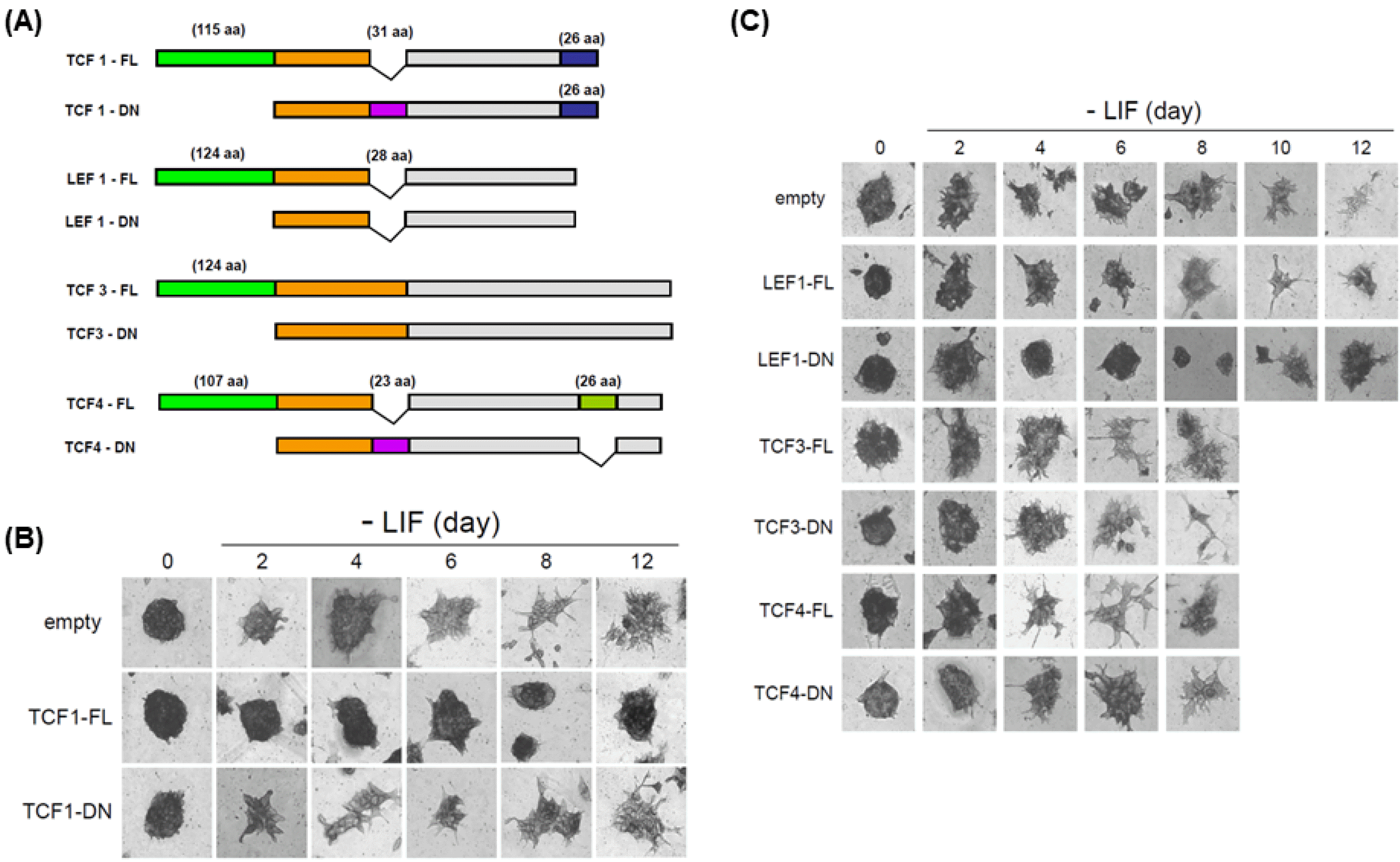
Fig. 2
The effect of ectopic expression of Tcfs and Lef1 after LIF depletion. (A) A schematic diagram of Tcfs and Lef1 constructs (FL, full length; and DN, dominant negative). (B, C) A6P10 ES cells were transfected with control vector, Tcf1 (FL or DN), Lef1 (FL or DN), Tcf3 (FL or DN), or Tcf4 (FL or DN) and then selected using Zeocin. Selected ES cells were stained with alkaline phosphatase (AP), a marker of self-renewal, on the days indicated after LIF removal.
![]()
To further explore how Tcf1 regulates the self-renewal of ES cells, we checked the levels of core stem cell markers; Oct4, Sox2 and Nanog upon Tcf1 overexpression at ES cells stage and Day 4 of differentiation. We found that during ES cells differentiation, the levels of Oct4, Sox2 and Nanog were reduced. However, upon overexpression of Tcf1, the reduction of stem cell markers was hindered. These effects were most predominantly seen in Nanog levels compared to Sox2 and Oct4 (Fig. 3A). We speculated that Tcf1 is the major regulator for Nanog, therefore we focused on Nanog in subsequent experiments.
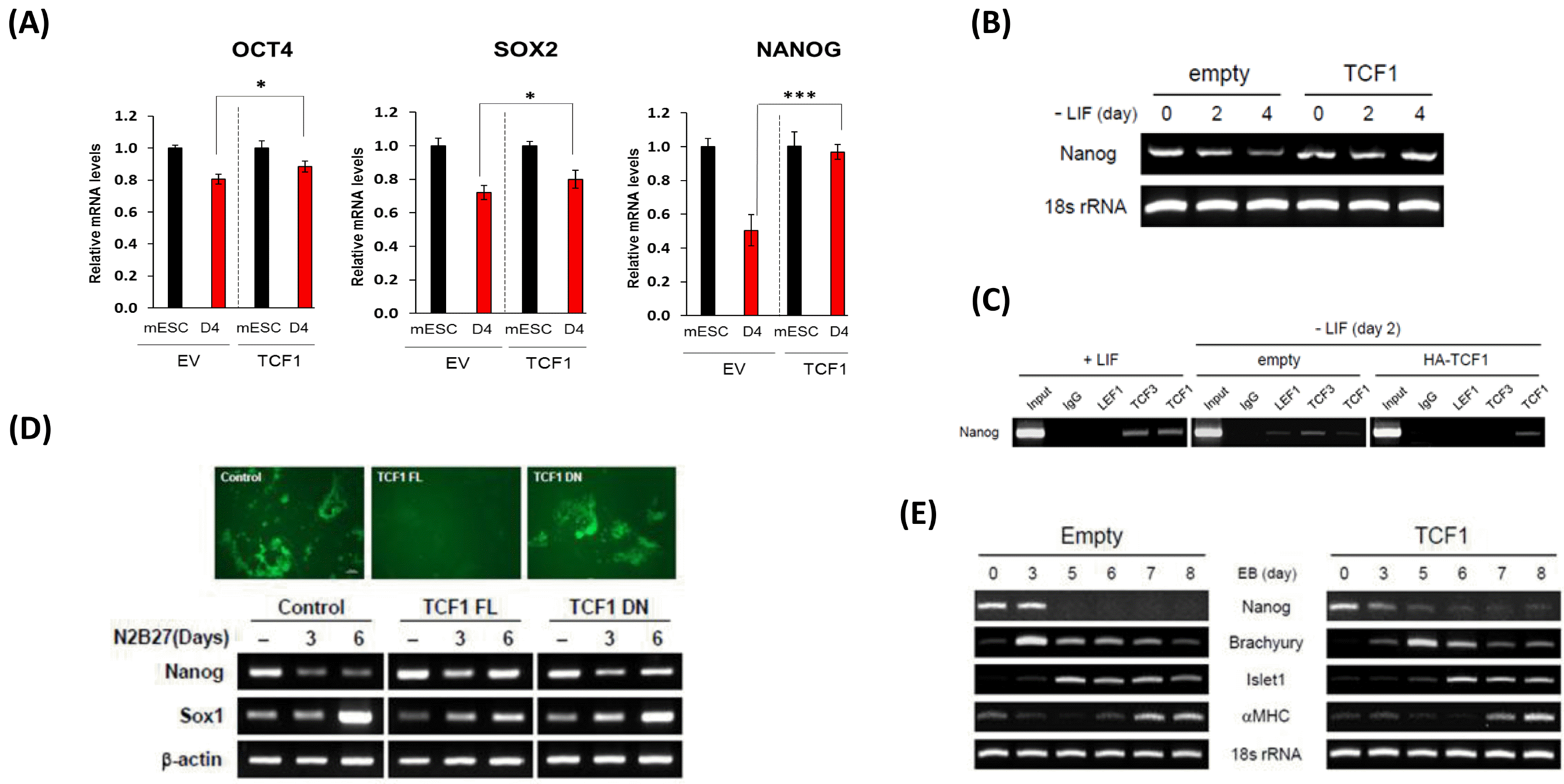
Fig. 3
Ectopic expression of Tcf1 maintains self-renewal and delays differentiation of ES cells. (A) mRNA expressions for Oct4, Sox2 and Nanog at ES cells stage and Day 4 of differentiation upon TCF1 overexpression were determined by using qPCR. RNA was isolated from cells expressing empty vector (EV) or Tcf1-FL (TCF1) on specified stage. GAPDH was used for normalization. (B) The amount of Nanog transcript was measured by RT-PCR analysis. RNA was isolated from ES cells expressing control vector or Tcf1-FL on the days specified. 18s rRNA was used as a loading control. (C) ChIP assay was performed to examine the binding of Tcf1, Lef1, and Tcf3 on Nanog promoter. Samples for ChIP were isolated from ES cells grown with LIF and from ES cells expressing control vector or Tcf1-FL cultured for 2 days without LIF. For immunoprecipitiation, endogenous antibody against Tcf1, Lef1, or Tcf3 was used. (D) To differentiate into neural precursors, 46C ES cells expressing control vector, Tcf1-FL, or Tcf1-DN were plated for 6 days in N2B27 medium. GFP fluorescence showed that 46C ES cells expressing control vector or Tcf1-DN, but not Tcf1-FL effectively differentiated into neural precursor (upper panel). The amount of Nanog or Sox1 transcript on the days indicated was measured via RT-PCR analysis (lower panel). β-actin was used as a loading control.
(E) RNA samples were isolated from A6P10 ES cells expressing control vector or Tcf1-FL on the days indicated after EB formation. Self-renewal (Nanog), mesoderm (Brachyury), and cardiomyocyte (Islet1, αMHC) markers were used for RT-PCR analysis.
![]()
Nanog is vital for the self-renewal of ES cells (7, 8) and its promoter is regulated by Tcf3 (27, 28). Since all Tcfs and Lef1 proteins bind to the same consensus sequence, we tested whether Tcf1 regulates the expression of Nanog in days specific manner. We found that Nanog expression was reduced in control ES cells after day 4 of LIF depletion, whereas the expression of Tcf1 inhibited the reduction in Nanog expression (Fig. 3B). These data indicate that ectopic Tcf1 induced the self-renewal of ES cells by promoting Nanog expression.
To investigate whether Tcf1 binds to Nanog promoter in practice, we performed a chromatin immunoprecipitation (ChIP) assay using an endogenous antibody for Tcf1, Tcf3, and Lef1 (Fig. 3C). In the presence of LIF, Tcf1 bound to the promoter of Nanog. Consistent with previous reports (27, 28), Tcf3 also interacted with Nanog promoter, suggesting that Tcf3 acts as an limiter of ES cell self-renewal. Since Lef1 protein is not expressed in the presence of LIF, no interaction between Lef1 and Nanog promoter was observed. Based on these data, we hypothesized that the occupancy of Tcf1 on Nanog gene may be reduced during differentiation, which was demonstrated using a ChIP assay for Nanog gene after LIF removal for 2 days. As expected, the occupancy of Tcf1 was reduced relative to that of Tcf3 and Lef1 (Fig. 3B). The protein levels of Tcf1, Tcf3, and Lef1 were the highest 2 days after LIF removal or after incubation with N2B27 media (Fig. 1B). Thus, the reduced binding of Tcf1 with Nanog promoter might be attributed to competition with Tcf3 or Lef1 protein. To support this possibility, ES cells stably expressing Tcf1 were used in the ChIP assay. Interestingly, most of the Nanog promoter was occupied by Tcf1 (Fig. 3B). Taken together, these results indicate that Tcf1 promotes the transcription of Nanog by binding with its promoter. The binding of Tcf1 protein with Nanog promoter is prevented by the increasing levels of Tcf3 or Lef1 protein during differentiation.
ES cell differentiation is prevented by Tcf1 expression
Nanog has been shown to be necessary for self-renewal and to prevent differentiation of ES cells in the absence of LIF (7, 8). Since ectopic expression of Tcf1 induced Nanog expression even after LIF depletion, we investigated whether overexpression of Tcf1 inhibited normal differentiation. For differentiation into neural precursors, we used Sox1-GFP knock-in (46C) ES cells in which the differentiated cells are easily detected based on the GFP signal when cultured with N2B27 medium (33). In control 46C ES cells, GFP-positive cells appeared after 6 days of incubation in N2B27 medium. However, the ectopic expression of Tcf1, but not TCF1-DN, inhibited the emergence of GFP-positive cells (Fig. 3D, upper panel). Consistent with the GFP signal, the expression of Sox1, a marker of neural precursors, was not induced in 46C ES cells expressing Tcf1 (Fig. 3D, lower panel). In addition, Nanog expression was not declined compared to the control cells after 6 days of incubation in N2B27 medium. These data indicate that overexpression of Tcf1 results in the inhibition of differentiation into neural precursors via maintaining the levels of Nanog.
To determine whether the inhibition of differentiation by Tcf1 overexpression occurred only in the case of differentiation into neural precursors, we further analyzed the cardiomyocyte differentiation via EB formation (Fig. 3D). While the expression of Nanog was diminished starting from day 5 in control EB, it persisted even after 8 days in EB expressing Tcf1. Moreover, the expression of marker genes related to cardiomyocyte differentiation was delayed in EB-expressing Tcf1. These data show that the ectopic expression of Tcf1 leads to delayed differentiation into cardiomyocytes. Overall, the overexpression of Tcf1 prevented normal differentiation via maintaining the level of Nanog although it is possible that Tcf1 may still regulate the genes related to differentiation.
Knockdown of Lef1 inhibits differentiation of ES cells
AP staining revealed that the ectopic expression of Lef1 did not induce self-renewal of ES cells. Interestingly, we also found that the expression of dominant-negative Lef1 (Lef1-DN) maintained ES cell self-renewal even after 12 days of LIF removal (Fig. 2C). In addition, endogenous Lef1 protein was transiently expressed following differentiation (Fig. 1B). These results suggested that Lef1 may be related to differentiation. To test this possibility, the level of Lef1 was reduced by shRNA against β-catenin-binding domain of Lef1 to knock down full-length Lef1 but not dominant-negative Lef1. Two days after LIF removal, the expression of Lef1 protein was remarkably reduced by shRNA for Lef1 (Fig. 4A). In order to identify the effect of Lef1 reduction, 46C ES cells expressing shControl or shLef1 were differentiated into neural precursors in N2B27 medium. Notably, 46C ES cells expressing shLef1 did not express Sox1-GFP compared with control after differentiation (Fig. 4B). Nevertheless, the level of Nanog in 46C ES cells expressing shLef1 was reduced in the control after 7 days of differentiation (Fig. 4B, lower panel). These results indicate that knockdown of Lef1 inhibits differentiation of ES cells into neural precursors without upregulation of Nanog expression. Similar to shLef1, the overexpression of dominant negative Lef1 blocked the differentiation into neural precursors. We further tested whether ectopic expression of Lef1 enhanced the differentiation of ES cells. Surprisingly, overexpression of full-length Lef1 did not enhance differentiation, which was blocked, instead (Fig. 4C). This result may be explained in part by the transient expression of endogenous Lef1 protein at the onset of differentiation (Fig. 1B). The effect of Lef1 reduction was also evaluated in the differentiation into cardiomyocytes. The expression of α-MHC, a marker for cardiomyocytes, was attenuated in ES cells expressing shLef1. Since EB can also be partially differentiated into endoderm, we investigated the effect of shLef1 on endoderm differentiation. Knockdown of Lef1 reduced the expression of GATA4, an endoderm marker (Fig. 4D). These data show that the reduced Lef1 expression attenuated the differentiation into cardiac mesoderm and endoderm. Taken together, these results suggest that the transient expression of Lef1 protein at the onset of differentiation is required for differentiation to three germ layers.
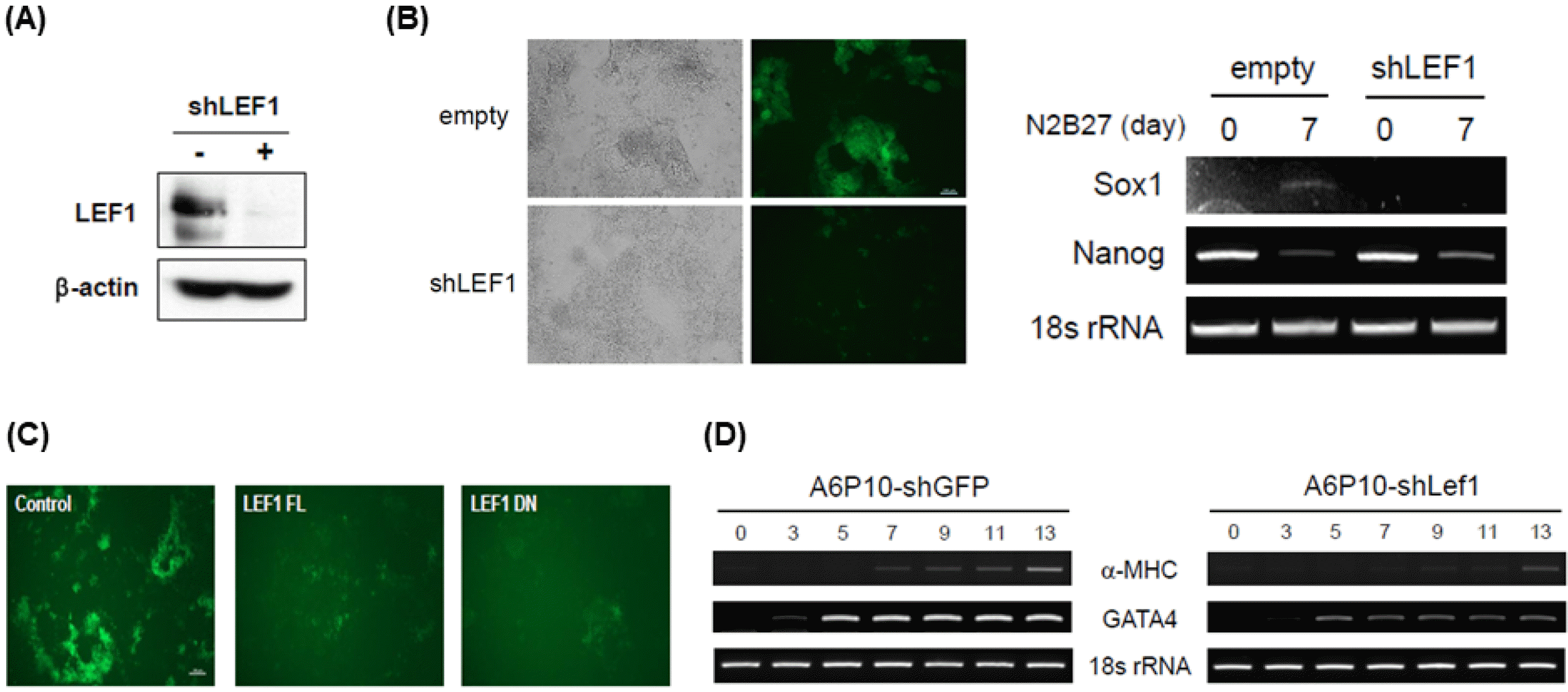
Fig. 4
Transient expression of Lef1 is required for proper differentiation. (A) A6P10 ES cells were infected with retroviruses expressing control or Lef1 shRNA, followed by selection using puromycin. After 2 days without LIF, the selected ES cells were harvested and analyzed by western blotting with anti-Lef1 antibody. β-actin was used as a loading control. (B) 46C ES cells expressing control shRNA or Lef1 shRNA were plated for 7 days in N2B27 medium. GFP fluorescence indicating a neural precursor was not induced in shLef1-46C cells (left panel). The amount of Nanog or Sox1 transcript was measured by RT-PCR analysis (right panel). (C) 46C ES cells were transfected with control vector, Lef1-FL, or Lef1-DN and then selected using Zeocin. 46C ES cells expressing each vector were plated for 7 days in N2B27 medium and GFP fluorescence was captured. (D) RNA samples were isolated from A6P10 ES cells expressing shGFP or shLef1 on the days indicated post-EB formation. Cardiomyocyte (α-MHC) and endoderm (GATA4) markers and loading control (18s rRNA) were used for RT-PCR analysis.
![]()
Discussion
Given that the effects of Wnt signaling vary, we speculated that the capacity of downstream effector molecules in the Wnt signaling pathway may differ depending on the status of ES cells. In this study, we found that the protein levels of Tcfs and Lef1 changed dynamically during the three types of differentiation and the expression pattern of each Tcfs and Lef1 varied (Fig. 1B). These results suggested that the individual Tcfs and Lef1 proteins may have a specific role in the regulation of self-renewal or differentiation of ES cells into specific lineages. Another interesting finding was that the expression patterns of Tcfs and Lef1 proteins varied, in part, from those of Tcfs and Lef1 transcripts. Especially, the level of Tcf3 protein declined sharply via unknown mechanism during EB formation while the level of transcript remained constant throughout differentiation (Fig. 1). The study of Tcfs and Lef1 1 function in ES cells has been limited to investigations at transcriptional level (27-30). Because Tcfs and Lef1 are regulated via unexpected degradation similar to post-transcriptional modification (26), the protein levels of Tcfs/Lef1 must be investigated to define their expression patterns.
Tcf1, Lef1, and Tcf4 have been generally used without distinction due to their structural and functional similarity in vitro, while Tcf3 has been used separately. However, knockout studies showed that Tcfs and Lef1 have non-redundant functions in part (25). In addition, our data showed varied levels of Tcfs and Lef1 protein (Fig. 1B). Based on these data, we hypothesized that each Tcfs and Lef1 protein may have a differential role in self-renewal or differentiation of ES cells. We found that the overexpression of wild type Tcf1contributed to self-renewal (Fig. 2B), whereas knockdown of Tcf1 had no effect on the undifferentiated state induced by LIF (data not shown). These results indicate that Tcf1 is sufficient, but not necessary, to maintain self-renewal. As Tcf1 protein level increased during differentiation (Fig. 1B), Tcf1 is possibly related to differentiation. To confirm this concern, further experiments are needed. Based on knockdown and ectopic expression of Lef1, we demonstrated that transient expression of Lef1 at the onset of differentiation was required for normal differentiation (Fig. 4). Interestingly, canonical Wnt transcripts are expressed similar to Lef1 expression pattern (19, 20) and TOP reporter activity is increased at the early stage of differentiation (unpublished data). These data suggest that Lef1 may induce cell-autonomous stimulation of differentiation by canonical Wnts.
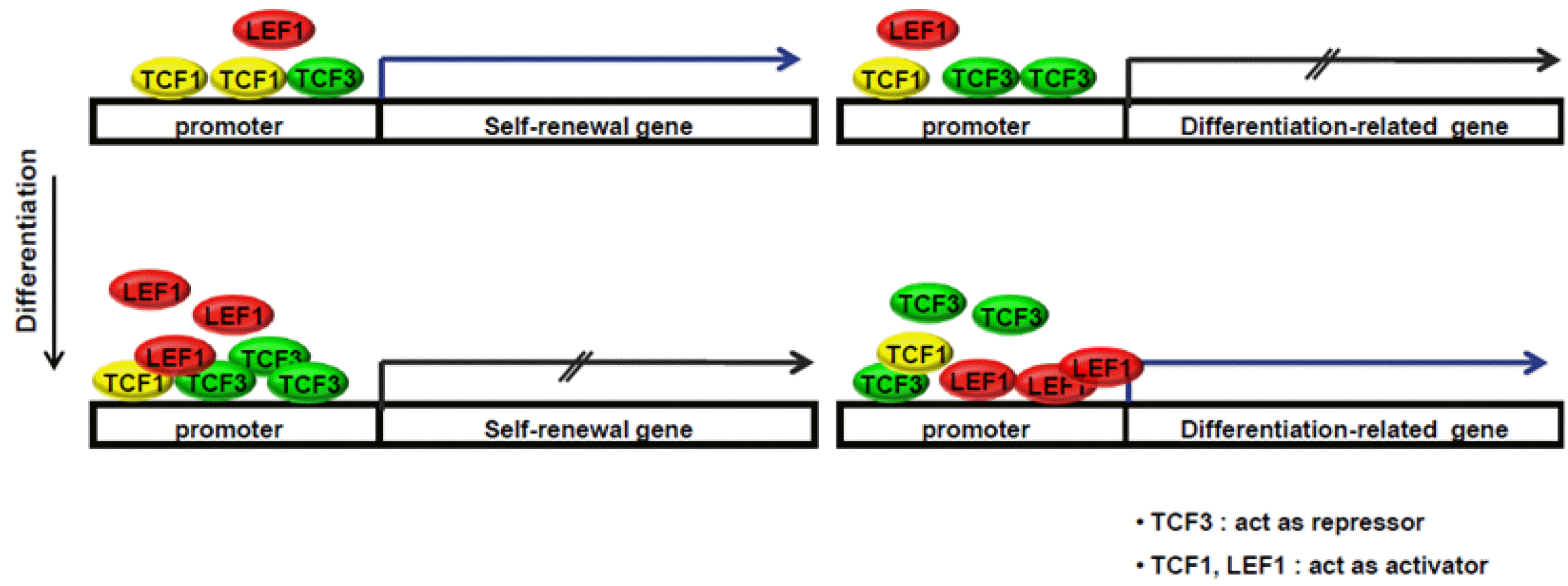
In summary, we identified distinct roles of Tcf1 and Lef1 in the regulation of self-renewal and differentiation of ES cells. Specific models illustrating the roles of Tcfs and Lef1 in the maintenance of stemness and regulation of differentiation are presented (Fig. 5). In undifferentiated ES cells, Tcf3 represses both self-renewal and differentiation-related genes, and Tcf1 mainly activates genes related to self-renewal to maintain stemness. During differentiation, increased Lef1 and Tcf3 may competitively bind to promoters of genes related to self-renewal and reduce the binding of Tcf1 on those promoters. In addition, increased levels of Lef1 may replace Tcf3 on gene promoters related to ES cell differentiation. In this study, we suggest that a balance between Tcfs and Lef1 is important for the modulation of self-renewal and differentiation in mouse embryonic stem cells.
Supplementary Materials
Supplementary data including one table can be found with this article online at http://pdf.medrang.co.kr/paper/pdf/IJSC/IJSC-13-s20044.pdf.
 XML Download
XML Download