

 PDF
PDF Citation
Citation Print
Print


Introduction
Mesenchymal stem cells (MSCs) are spindle-shaped multipotent non-hematopoietic adult stem cells, first characterized by Friedenstein et al., in 1974 (1). MSCs originate from the mesoderm, possess extensive proliferative and self-renewal potential with the ability to attach to plastic and gather in colonies called colony forming units (CFUs) (2). MSCs are immune-privileged because of the lack or low expression of class-II major histocompatibility complex (MHC-II) and other costimulatory molecules (3, 4). Nowadays, it has been shown that MSCs can be established by differentiation of pluripotent stem cells like embryonic stem cells (ESCs) and induced pluripotent stem cells (iPSCs). The pluripotent stem cells-derived MSCs (PSCs-MSCs) have presented better self-renewal capability, engraftment efficacy and higher therapeutic potency than MSCs derived from adult tissues. The PSCs derived-MSCs replace damaged cells in injured tissues, exert immunomodulatory effects, release matrix proteins and growth factors that eventually adjusts the surroundings of damaged tissues. These PSCs-MSCs are evidently useful in regenerative medicine to treat a number of unmanageable immunological and neurological disorders (5, 6).
MSCs are found in multiple tissues, including bone marrow (BM-MSCs), umbilical cord blood (UCB-MSCs), umbilical cord (UC-MSCs), placenta (PL-MSCs), adipose tissue (AT-MSCs), fetal liver, muscle, lung, and heart which differs in their biological characteristics (7). MSCs have the ability to differentiate into cells of the mesodermal lineage (adipocytes, chondrocytes, osteoblasts, osteocytes, and myoblasts). However, few studies have also shown the ability of differentiation into ectodermic (neuron-like cells) and endodermic (hepatocytes) cell lineages (8, 9).
Although conflicting results have been reported, MSCs can be recognized according to the expression of a universal panel of cellular markers proposed by the International Society for Cell Therapy (ISCT). Using antibody-based classification methods, minimal criteria for defining human MSCs included positive expression of CD44, CD73 (SH3), CD90 and CD105, and negative expression of CD45, CD34, CD14, CD79a and HLA-DR (10). Additionally, using high-throughput techniques, transmembrane 4 L6 family member 1 (TM4SF1) have been proved effective as MSC-specific surface marker (11).
Since no available data have shown detrimental reactions to allogeneic or autologous MSCs transplants, these cells are great candidates for cell-therapy (12, 13). Furthermore, being more available, free from ethical issues, easy to isolate and expand, able to differentiate to multiple lineages and immunosuppressive, have made MSCs better candidates than embryonic stem cells (ESCs) and induced pluripotent stem cells (iPSCs) (14). Consequently, MSCs are considered to be safe as they are tested in many clinical trials of neurodegenerative, immunological and also cardiovascular diseases, which have had promising results (13, 15, 16).
Neurological diseases cause a huge burden on global health and their occurrence is growing in low and middle-income countries (17, 18). Numerous factors, including genetic and environmental components, may involve in the development and progression of this kind of diseases (19). The term “neurodegenerative diseases” consists of a lot of diseases like Alzheimer’s disease, Parkinson’s disease, and Huntington’s disease (19-21). Aggregation of insoluble forms of abnormal proteins between central nervous system (CNS) cells is a pathological characteristic seen in almost all of neurodegenerative diseases (22, 23). Following disease progression, CNS degrades gradually and neuronal functions and structures are lost (24). Our aim in this review is to summarize the most important manipulation methods that can be utilized to enhance MSCs survival and potency and their applications in treating the most prevalent neurodegenerative diseases.
Go to : 
Immunomodulatory, Neuroprotective and Regenerative Capacities of MSCs
Having the unique capacity to modulate immune cells (25, 26), MSCs are exclusive target for immunomodulation purposes (27, 28). The point that these cells are almost non-immunogenic rises from the fact that MSCs only express human leukocyte antigen-I (HLA-I) and lack HLA-II and other costimulatory molecules like CD80 and CD86, needed for T cell activation (27, 29). Although MSCs from different sources exert their immunomodulatory functions in different ways, the fundamental mechanism is shared among them. Both cell-cell contact and secreted molecules are involved in immunomodulatory properties mediated by MSCs (25, 30, 31).
Considering inflammatory responses as an inseparable part of neurological diseases, MSCs have shown themselves as a promising therapeutic approach for treatment of some of these unmanageable neurological disorders. Notably, neuroprotective and regenerative effects of MSCs results from their immunomodulatory and anti-inflammatory potential. Neuroprotection and therapeutic effects of MSCs in experimental CNS disease models have been shown in various studies. One of these researches unveiled the neuroprotective effects of AT-MSCs in models of Parkinson’s disease (32, 33). Similarly, regenerative capabilities of MSCs have been shown in experimental rats where administration of MSCs derived from dental pulps enhanced both motor nerve regeneration and behavioral activities (34). MSCs have the ability to secrete various growth factors, making the damaged tissue microenvironment ready for regeneration and tissue repair. For instance, insulin growth factor binding protein 6 (IGF BP-6) secreted by MSCs improves recovery from nerve injury in H2O2-injured cortical neuron cultures through inhibiting IGF-2 and interacting with insulin growth factor 1 receptor (IGF-1R) (35). In addition, hypoxic-ischemic brain damage (HIBD) is a prevalent cause of newborn death. Using PL-MSCs as a treatment in this situation prevented the production of pro-inflammatory cytokines and enhanced the production of IL-10 in the ischemic hemispheres of HIBD rats (36). On the other hand, in a rodent stroke model, it has been shown that intravenous injection of UC-MSCs improved functional losses and decreased neuronal damage (37). Suppressing inflammatory responses is the main effect of PL-MSCs and UC-MSCs for attenuating HIBD and relieving stroke effects, respectively (36, 37).
Since the therapeutic properties of MSCs are mainly exerted by affecting both innate and adaptive immune responses (38), hereby we discuss the most important immunomodulatory effects exerted by MSCs on immune cells.
Antigen-presenting cells
Antigen-presenting cells (APCs) include dendritic cells, macrophages and B cells that process and present antigens to T cells. Since DCs initiate immune responses via presenting antigens to naïve T cells, they appear to be more important (25, 39). MSCs inhibit monocytes from differentiation into DCs in the periphery which results into lesser induction of T cell response and cause M2 macrophage differentiation (27, 40, 41). The major soluble factors implicated in these events are prostaglandin E2 (PGE2), Interleukin 6 (IL-6), and macrophage colony-stimulating factor (M-CSF) (25). Different studies have been conducted to elucidate the mechanisms by which MSCs inhibit the maturation and function of the DCs. In a coculture system of BM-MSCS and DCs it has been shown that: (1) overexpression of microRNA-23b (miR-23b) in MSCs inhibited DCs maturation by blocking the NF-κB pathway (42); (2) concentrations of galectin-1 (Gal-1), IL-10, and IL-12 were increased in the supernatants and the protein expression of Gal-1 on and within MSCs was also enhanced which hampered the function of DCs (43).
Natural killer cells
Natural killer cells play an important role in cell innate immunity against tumor and virus-infected cells. MSCs can significantly reduce the proliferation and activation of NK cells through direct cell contact and also soluble factors like indoleamine 2,3-dioxygenase (IDO) and PGE2 production. The secretion of sHLAG5 by MSCs lessen NK cells cytolytic activity and IFN-γ production of these cells (25, 44, 45). In addition, MSCs have been demonstrated to reduce the secretion of IFN-γ and TNF-α by NK cells.
It is known that NK cells can kill cells by detecting the lack of HLA-I molecules on target cells. Prominently, expression of HLA-I molecules on the surface of MSCs makes them resistant to lysis by NK cells. The HLA-I expression on MSCs is even amplified when the cells are pretreated by IFN-γ, used for strengthening the anti-inflammatory effects of MSCs (27, 46).
T cells
T lymphocytes, predominantly produced in the thymus, are mainly involved in cellular immune response. It is well defined that MSCs inhibit T cells proliferation more than 50% (29, 47). BM-MSCs and UCB-MSCs cause the arrest of T cell cycle while AT-MSCs and UC-MSCs induce apoptosis to inhibit T cell proliferation. BM-MSCs reduce Thelper1 (Th1) and Th17 cells-derived cytokines and also cause upregulation of the anti-inflammatory cytokines production such as IL-4 from Th2 cells. The expression of inhibitory molecules like PDL1 and PDL2 or HLA-G, expressed by all PL-MSCs and also the lack of costimulatory molecules on MSCs led to T cells anergy. Further, MSCs cause Treg cells proliferation which can be enhanced by priming MSCs in hypoxia, as hypoxia increases the expression of IDO.
B cells
B lymphocytes are another component of the adaptive immune system which has important roles in the initiation and progression of many diseases, mostly by secreting antibodies (48). MSCs can be really effective in ameliorating these effects as they cause decreased B cell proliferation, differentiation, chemotactic properties and antibody-secreting ability (49-52). On the other hand, MSCs lead to augmentation of B lymphocytes viability. For instance, placental MSCs interfere with B cells apoptosis (Fig. 1) (40).
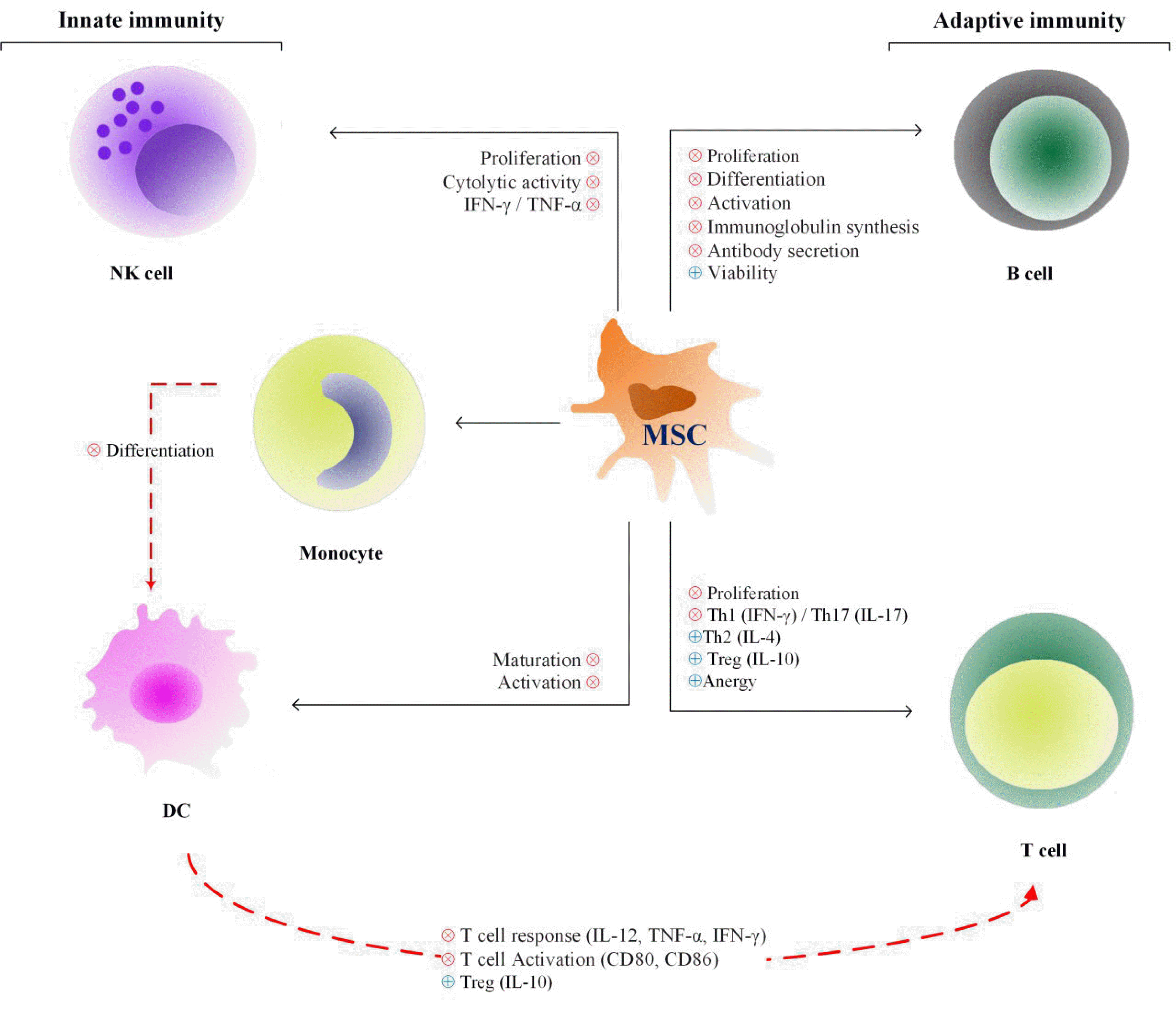 | Fig. 1MSCs immunomodulatory effects on immune cells. MSCs have inhibitory effects on both innate and adaptive immunity. Their immunomodulatory impacts which are conducted by secretion of soluble factors and cell to cell contact include: (1) inhibition of B and NK cells proliferation and functions; (2) suppression of T cells proliferation and Th1 differentiation, induction of TH2 and Treg cells; (3) inhibition of dendritic cell and monocyte maturation and activation.
|
Go to :
Manipulation of MSCs
To improve therapeutic characteristics, MSCs can be manipulated by different approaches. They can be pretreated with various chemical, physical and biological agents, and modified with varied genetic and/or epigenetic manipulations to enhance their functions, efficacy and safety of transplantation (53-55).
Pretreatment (preconditioning) of MSCs
MSC source: MSCs can be isolated from many adult and fetal tissues. However, MSCs from diverse origins have various characteristics. This offers that sources of MSCs may have an impact on their cellular destiny and behavior. Briefly, the overall orientation considering MSCs origin can be expressed as follows: (a) MSCs derived from a certain origin can differentiate better to cells of that tissue than other MSCs (BM-MSCs have better osteogenesis capacity than the others); (b) fetal MSCs, mainly, have a higher proliferation capacity than adult MSCs; (c) if the immunomodulatory effects are substantial, fetal MSCs or AT-MSCs are better options than BM-MSCs. (d) Overall, AT-MSCs reveal great in vivo capability for cell therapy (56, 57).
Hypoxia preconditioning: Typically, the oxygen concentration in MSC niches is rather low. A number of studies have shown that MSCs may be influenced by greater oxygen concentrations in ex vivo culture and their viability decreased (54, 58). Therefore, it seems that hypoxia preconditioning of MSCs, before using them in different therapeutical applications, is a suitable method to prepare them to face and endure difficult situations. Hypoxia preconditioning is applied by culturing MSCs at 2.5% O2 (for 15 minutes) and reoxygenation at 21% O2 (for 30 minutes) which is followed again by hypoxia preconditioning at 2.5% O2 (for 72 hours) (59, 60). The process of hypoxia and reoxygenation enhances the expression of pro-survival genes (55, 61).
In fact, when MSCs are cultured with 1∼7% O2, they are preconditioned by hypoxia and this manipulation enhances different aspects of MSCs characteristics (55). Hypoxia improves proliferation and migration of MSCs, and also causes lower expression of senescence-associated molecules like caspase-3 and β-galactosidase in these cells (54). Moreover, by improving the secretion of anti-inflammatory cytokines and other soluble factors, hypoxia pretreatment increases immunosuppressive characteristics of MSCs. For instance, human gingiva-derived MSCs produced more IL-10 and express more “Fas ligand (FasL)” when they were exposed to hypoxia. As well, human AT-MSCs pretreated with hypoxia showed upregulated IDO expression, causing T cell anergy, Treg generation, and increased IL-10 production (60, 62, 63). Hypoxic pretreatment makes hypoxia-inducible factor-1α (HIF-1α) more stable (54). HIF-1α reduces reactive oxygen species (ROS) production, promotes glycolysis (54) and induces expression of 78-kDa glucose-regulated protein (GRP78) which enhances cell proliferation through Akt pathway. After 12 h of hypoxia, GRP78-mediated phosphorylation of Akt, mTOR, and P70S6k (ribosomal protein S6 kinase β-1) was increased. GRP78 also increases cell cycle-associated proteins such as CDK2, cyclin E, CDK4, cyclin D1 through the regulation of Akt signaling pathway (64). HIF-1α activates nuclear factor kappa B (NF-κB), increase antioxidant and antiapoptotic proteins like Bcl-xl and Bcl-2 (54). Hypoxia preconditioning of MSCs also upregulates the expression of normal cellular prion protein (PrPc), which regulates superoxide dismutase (SOD) and catalase activity and hampers oxidative stress-induced apoptosis by inactivation of cleaved caspase3 (54, 65).
Nutrient deprivation: MSCs confront an ischemic microenvironment, recognized by reduced oxygen (O2) tension (hypoxia) and nutrient deprivation (serum or glucose starvation) concurrently, that may threaten survival of the MSCs, especially in tissue engineering. Human MSCs were exposed to (I) O2 deprivation (hypoxia), (II) serum starvation (for 48 hours), and (III) prolonged (up to 120 hours) hypoxia associated with serum deprivation (ischemia). As a result, the MSC apoptosis rates were not influenced by 48-hour hypoxia, but elevated through fetal bovine serum (FBS) starvation, indicating that nutrient deprivation is the stronger factor than hypoxia (66). Long-term hypoxia along with serum starvation bring almost complete apoptosis of MSCs, but this rate was decreased by half when MSCs were encountered hypoxia and 10% FBS. This finding shows that MSCs are susceptible to the concurrent serum and O2 deprivation which they face when transplanted in vivo (67). It evokes the development of new methods in cell therapy by MSCs.
Pretreatment of MSCs with cytokines: Cytokines affect interactions and communications between variouse cells. Likewise, MSCs are no exception to this fact and considerable work have been done to elucidate these effects. In comparison to others, proinflammatory cytokines like IFN-γ, TNF-α, and IL-1β exert more modulatory effects on MSCs (68).
Pretreating MSCs with IFN-γ enhances the production of PGE2 and IDO (responsible for the suppression of T and NK cells proliferation). These MSCs inhibit NK cells activation and cytotoxicity and block production of Th1-related cytokines (IFN-γ, TNF-α, and IL-1) (55, 60). TNF-α activates Erk1/2 and MAPK signaling pathway and causes increased proliferation and osteogenic differentiation of MSCs (55). Modulatory effects of TNF-α are seen more when MSCs are pretreated with both of IFN-γ and TNF-α. This combination increases the production of IDO, PGE2, PDL-1, and HLA-G. IDO and PGE2 are important in the secretion of factor H, a complement system inhibitor. CD206+ M2 macrophages are generated from CD14+ monocytes to suppress PBMCs proliferation. Alongside M2 macrophage generation, both cytokines result in enhanced PDL-2 expression by MSCs and CD4+ IL-10+ and CD8+ IL-10+ Treg cells differentiation. IFN-γ and TNF-α have the ability to induce the production of proinflammatory chemokines such as CCL5, CXCL9, CXCL10, CXCL11 through MSCs. These chemokines accumulate immune cells in close proximity to MSCs in order to lay them more exposed to MSCs immunosuppressive effects (55, 60, 69, 70).
IL-1β-treated MSCs show significant immunomodulatory properties such as enhanced migratory abilities through the increased expression of CXCR4 and other related chemokines. Production of several chemokines is another immunomodulatory influence of MSCs enhanced by IL-1β, resembling TNF-α effect. It can also cause MSCs to produce certain cytokines that can regulate the function of target cells (60, 71, 72).
Additionally, IL-17A-treated MSCs, cocultured with CD4+CD25− T cells, are able to increase CD4+CD25hiCD127loFoxP3+ Tregs (60). IFN-α-preconditioned MSCs also show increased migratory potential and they also inhibit Th1 inflammatory responses (55, 73). Stromal-derived factor-1 (SDF-1), known as CXCL12, is a chemokine used for MSCs preconditioning. This chemokine enhances the production of angiogenic cytokines through activation of Akt and Erk signaling pathways. SDF-1 rescues MSCs from H2O2-induced apoptosis which is followed by increased migration and survival rate of MSCs (55, 74).
Pharmacological and clinical drugs: Mesenchymal stem cell-based therapy is a great choice for treating CNS diseases, although loss of the cells after transplantations remains a serious setback. Exposure of MSCs to different pharmacological agents is another preconditioning method for strengthening their survival when they encounter the harsh microenvironment of the damaged tissue (75, 76).
Atorvastatin (C33H35FN2O5), a lipid-lowering statin, commonly used to prevent cardiovascular-related diseases. When used as a preconditioning agent, it can improve post-implantation survival of MSCs (77). This drug increases surface expression of CXCR4 in MSCs and stimulates migratory abilities of these cells by CXCR4/SDF-1 involvement (78).
Trimetazidine (C14H22N2O3) is an anti-ischemic drug used for treating angina in cardiac patients. In comparison to non-conditioned cells, Trimetazidine-preconditioned rat MSCs showed both notable protection against H2O2-induced membrane damage and increased viability. This drug increases HIF-1 α, survivin, BCl-2 gene expressions in MSCs (79).
Preconditioning of MSCs with deferoxamine (DFX); an iron-chelating drug, stabilizes HIF-1α and enhances the expression of genes involved in cell migration. Deferoxamine also affects the homing of MSCs by enhancing the expression of chemokine receptors (77). Adipose tissue-derived-MSCs preconditioned with 150∼400 μM of DFX showed upregulated mRNA levels of pro-angiogenic factors; like vascular endothelial growth factor (VEGF), neuroprotective factors; such as nerve growth factor, neurotrophin 3, and anti-inflammatory cytokines (i.e. IL-4 and IL-5) (80).
Following pretreatment with LPS, an endotoxin in the cell wall of gram-negative bacteria, it binds to TLR4 on MSCs, activates PI3K/Akt pathway and increases expression of neural epidermal growth factor (NEGF). The problem of using unpreconditioned MSCs for infarcted myocardium is the low survival of these cells. Pretreatment of MSCs with 1 μg/ml and then transplantation of these cells into ischemic myocardium yielded increased survival and cardiac protective capacity of MSCs. For that reason, MSC pretreatment with LPS ameliorates cardiac function after myocardial infarction (77, 81).
Curcumin, the active part of turmeric and diazoxide which is a mitochondrial ATP-sensitive potassium channel, protect MSCs from oxidative stress injury (82, 83). Lithium and valproic acid (VPA)-preconditioned MSCs indicate enhanced expression of different genes like antioxidant, anti-apoptosis, and migration-related genes. The increased expression of these genes causes augmented survival of MSCs following transplantation (75).
In addition, vitamins also could be used for pretreating of MSCs. Vitamin E, a representative of vitamins, decrease both oxidative stress caused by H2O2 and apoptosis-related in MSCs. This vitamin also enhances the expression of proliferative markers and TGF-β (55, 84).
Heat shock pretreatment (HSP) of MSCs: Heat shock pretreatment is an efficient method for elevating anti-apoptotic properties of MSCs. It has been reported that HSP pretreatment lessens apoptosis of MSCs and enhance their survival in the injured tissue, especially heart, liver and ovary, by induction of autophagy. However, no research studies have been conducted on HSP pretreatment for MSCs application in a wide range of diseases (85, 86).
MSCs pretreatment by freeze and thaw (cryopreserved MSCs): The efficacy of MSCs therapy will be determined by a great part according to the phenotype of MSC after preparations. Cells can sustain considerable changes in response to alterations in their microenvironment. Freeze and thaw, as one of these changes, has been demonstrated to affect the immunomodulatory properties of MSCs. However, MSC functions are entirely revived after a 24 h culture period. After thawing, there were alterations in the genes involved in innate immunity pathways and in cytoskeletal rearrangement. Furthermore, thawed MSCs are vulnerable to T cell mediated cytotoxicity but this phenomenon is partially saved by IFN-γ pre-licensing in MSCs (87, 88).
Cell-free therapy
MSCs cultured in different medium have been shown to secrete different factors referred to as MSCs conditioned medium, the secreted agents in the medium from MSCs as a whole, and microvesicles, exosomes and apoptotic bodies as well (89).
Conditioned medium of MSCs: Mesenchymal stem cells secrete different soluble factors into the culture medium. These secreted factors are referred to as conditioned media (CM), composed of growth factors like VEGF, epidermal growth factor (EGF), platelet-derived growth factor (PDGF), insulin-like growth factor (IGF-1), hepatocyte growth factor (HGF), pro/anti-inflammatory cytokines and other molecules like Leptin, angiogenin, granulocyte-macrophage colony-stimulating factor (GM-CSF), fractalkine, monocyte chemoattractant protein-1 (MCP-1), SDF-1/CXCL12. Mesenchymal stem cells from different sources produce variable amounts of conditioned medium depending on culture medium type, period of culture (16 hours to 5 days) and culture condition (monolayer or spheroid) (89, 90). MSC conditioned medium (MSC-CM) possess both differentiation capacity and therapeutic function. UC-MSCs-derived conditioned medium (UC-CM) is used for different clinical applications like wound healing, helping to induce apoptosis and differentiation. The conditioned medium has anti-fibrotic properties and regenerative abilities and can suppress the proteolytic system and ROS generation in muscle atrophied cells (91, 92).
Administration of MSCs-CM in a mouse model of neuropathic pain induced by PSL (partial sciatic nerve ligation) exhibited longer antinociceptive effect in comparison to gabapentin, which is conventional treatment of neuropathic pain. Conditioned media also decreased the levels of proinflammatory cytokines including IL-1β, TNF-α, and IL-6 while increasing IL-10 concentrations (93).
In autoimmune diseases like rheumatoid arthritis (RA), MSC-based therapies can also yield positive results. The use of conditioned medium in antigen-induced arthritis (AIA) significantly lessened knee swelling, cartilage loss, and some other pathological signs of AIA. It also inhibited TNF-α production and increased IL-10 production through enhancing Treg cell functions (94).
Administration of conditioned medium derived from MSCs showed significant therapeutic effects on the pulmonary ischemic situation and myocardial infarction (MI). Treatment of lung ischemia-reperfusion injury by CM reduced proinflammatory cytokines, infiltration of inflammatory cells and increased Tregs and M2-like macrophages in the site of injury (95). Three weeks after treating by MSCs-CM, MI pigs showed higher myocardial capillary density, decreased myocardial infarct size and better systolic and diastolic performance (96).
Oxidative stress affects cell survival and it is important to prevent its effects on the cells. Conditioned medium taken from MSCs alleviated injury of H2O2-induced oxidative stress in hepatocyte cells, and decreased levels of miR-143, which is increased after oxidative stress. The conditioned medium also restored Bcl level and decreased Bax and BMF levels in hepatocytes (97). The use of conditioned medium as a cryoprotectant results in reduction of apoptosis level in CD34+ umbilical cord blood cells and protection of these cells from oxidative stress due to ROS generation (98).
MSCs-derived exosomes and microvesicles: Exosomes and microvesicles are two secreted vesicles released from MSCs along with apoptotic bodies (53). These extracellular vesicles are distinguished from each other by their size, flotation density on sucrose, lipid composition, protein cargo and biogenesis pathway (99).
Exosomes are molecules with bilayer membrane with 40∼100 nm diameter which is secreted by most of the cells. They contain lipids including cholesterol (mostly), sphingomyelin, ceramide, proteins, and nucleic acids. Certain cluster of differentiation markers including CD9, CD63 and CD81 and tumor susceptibility gene 101 (Tsg101) as well as MHC-I and II are expressed on exosomes (53, 100). They are generated from early endosomes or multivesicular bodies and are released through the fusion of the endosome with the cell surface in a P53-regulated exocytosis process. This process is dependent on cytoskeletal activation but independent of cellular calcium. These molecules can be released in almost all kinds of fluids like blood, CSF, urine, amniotic fluid, ascites, saliva, seminal fluid, and etc. There are several methods to obtain exosomes from MSC cultures. Ultracentrifugation and polymer-based precipitation kits are the most common methods (53, 101). Exosomes are enriched with numerous bioactive molecules like proteins, lipids, mRNAs, micro RNAs, genomic DNA, cDNA, and mitochondrial DNA. They also contain cytokines and growth factors with immunomodulatory effects, including TGF-β, IL-6, IL-10 and HGF (53). Exosomes contents are protected from the RNases and trypsin by its bilayer lipid membrane (100). These secreted exosomes are involved mostly in intercellular communication and transportation of genetic material unleashed by MSCs. They have the ability to interact with various cells in close proximity and make them show appropriate response. One of the very important roles of MSCs-derived exosomes is their ability to support maintenance of homeostasis within the tissue especially in the event of diseases (53, 99).
In almost various types of diseases, the usage of MSCs produce good results. However, considering some of the risks that could happen by the administration of MSCs, researchers are encouraged to regard different aspects of these cells. Since most of the effects of MSCs are exerted through their paracrine actions, exosomes have been a good candidate for cell free therapy. Some of the risks that could arise by using MSCs but won’t happen by the use of exosomes are as follows: (1) Persistency of MSCs proliferation when the need is over, so it can cause ectopic tissues formation (100,102-104) in liver diseases. Although administration of MSCs helps to regenerate hepatocytes and reduce liver inflammation, the risk of iatrogenic tumor formation and cellular rejection remain unsolved (105). (2) Because of the large cell size of MSCs, the intravenous injection could cause occlusion, and ossification and calcification in tissues which caused long-term safety concerns (100). Overall, MSCs-derived exosomes have shown promising therapeutic effects when used in different kinds of diseases like cardiovascular diseases, immune-mediated diseases, neurological diseases and tumor (53).
In neurological diseases, exosomes can help in the removal of unwanted stress proteins and amyloid fibril formation. Exosomes also participate in the release of β-amyloid in Alzheimer’s disease (100, 106, 107). The use of exosomes in a model of spinal cord injury (SCI) has attenuated lesions and enhanced functional recovery after SCI. Exosomes also have been shown to decrease gene expression of Bax (apoptotic agent) and proinflammatory cytokines (IL-1β, IL-6, TNF-α) and increase BCL-2 and IL-10 (108).
Exosomes and microvesicles obtained from MSCs protected mice against joint damage in the osteoarthritis model (OA) by increasing the expression of chondrocyte markers (Type II collagen and aggrecan), while decreasing inflammatory markers such as induced nitric oxide synthase (iNOS) (109). Furthermore, in an atopic dermatitis mouse model, the use of adipose tissue-derived MSC exosomes decreased pathological symptoms and clinical score of the disease (109).
Microvesicles like exosomes are nanosized (100∼1000 nm), bi-lipid layered vesicles (53, 110). These vesicles are secreted out of MSCs through budding of plasma membrane in a Ca2+-dependent pathway. Microvesicles express ARF6, VCAMP3, phosphatidylserine, and lipid raft-associated molecules (tissue factor and flotillin) and they are also enriched with CD40. These secreted microvesicles transfer different components and information such as mRNAs, micro RNAs, and proteins among stem cells and tissues (111). Microvesicles can be regarded as a good alternative therapeutic vehicle to MSCs. One good property that make them good candidate for stem cell-based therapy is that repeated administration of these factors do not incite immune responses. However, large-scale production of microvesicles and their long-term safety are issues that should be taken into consideration before using them as regenerative agents (100, 111).
Microvesicles can cause deviations in T cells differentiation, increase Th2 and regulatory T cell subsets. They can also augment apoptosis level in PBMCs and CD3+ T cells (112). Microvesicles can have a tolerogenic activity which can bring peripheral tolerance among autoreactive cells. Expressing PD-L1, galectin-1 and membrane-bound TGF-β enable exosomes to inhibit proliferation of autoreactive cells and promote Treg cells generation more (113).
Microvesicles have also shown promising results when utilized in treatment of different diseases. It was shown that HGF released from MSCs-microvesicles was the agent to restore endothelial barrier function and decrease paracellular and transcellular permeability (114). Human embryonic MSCs-derived microvesicles presented promising therapeutic activities in anti-tumor defense; inhibiting leukemia cells proliferation, decreasing the BCL/Bax ratio and upregulating autophagy which can induce apoptosis (115). The use of human MSCS-derived microvesicles on pneumonia-infected mice exhibited increased survival and decreased invasion of inflammatory cells, cytokines, and bacteria to the site of injury. It has also been shown that pretreatment of MSCs with TLR3 agonists could intensify the therapeutic effects of the microvesicles (116). A good therapeutic effect of MSCs-derived microvesicles through the transfer of mRNA has been revealed in an acute kidney injury (AKI) model. Microvesicles made epithelial cells more resistant to apoptosis and accelerated their functional recovery from glycerol-induced AKI in severe combined immunodeficiency (SCID) mice (117).
MSCs-derived apoptotic bodies: Another member of the extracellular vesicles are the apoptotic bodies. Apoptotic bodies’ size is about 500∼4000 nm.These vesicles can be derived from any dying cell. Excessive production of them, due to increased apoptosis, can lead to several neurological disorders like Parkinson’s and Alzheimer’s disease. Visa versa, reduced production of apoptotic bodies, as a result of decreased apoptosis, can lead to autoimmune disease development (118). They are enriched in histones, DNAs and noncoding RNAs and cell organelles are found inside of these extracellular vesicles. Apoptotic bodies express certain molecules like TSP, C3b and macrophage-related markers (53, 119, 120).
These vesicles are absorbed by macrophages, dendritic cells, endothelial cells and fibroblasts and then they are degraded inside lysosomes of these cells. The cells use αvβ3 integrin to absorb apoptotic bodies. Phosphatidylserine on the surface of apoptotic bodies, as an “eat me” signal, play an important role in their recognition by phagocytes. Research studies on the therapeutic effects of apoptotic bodies are not as abundant as researches on exosomes and microvesicles. It has been shown that formation of apoptotic bodies has a significant role in osteogenic and adipogenic differentiation of bone marrow MSCs and they are involved in bone homeostasis and can be used to treat osteoporosis (118, 121).
Epigenetic manipulation of MSCs
Epigenetics is the set of inheritable alterations involving the expression of genes without mutations in the DNA sequence. For gene expression and other chromatin modifications, genes interact with transcriptional factors and nucleus modulators. These interactions are regulated by DNA methylation, histone modifications, microRNAs and chromatin remodeling which determine the fate of MSCs, their commitment and differentiation (122, 123). For instance, one method of cell therapy, is the use of microRNAs transfection, such as microRNA-378, under hypoxic-ischemic condition, which increase survival rate of MSCs (124). Also, Exosomal microRNAs (miRNAs) released by MSCs are hopeful therapeutic target for MI, Stroke, Alzheimer’s disease (AD), Parkinson’s disease (PD), and osteoarthritis (OA). This could be dependent on the role of miRNA in desired tissue biology, including tissue regeneration, cell differentiation, apoptosis, neovascularization, and remodeling (125).
Genetic manipulation of MSCs
Genetic manipulation is another way to improve MSC. Cumulating evidence have shown hopeful results of this method in the treatment of neurodegenerative, inflammatory and autoimmune diseases (126). Genetic modification of MSCs can drive different processes of cell differentiation like osteogenesis, adipogenesis, angiogenesis, and chondrogenesis (127). MSCs secrete several proangiogenic factors which promote formation of new vessels when used without any modifications (127). One of the most important proangiogenic factors is VEGF (128). It has been shown that VEGF-modified MSCs promoted angiogenesis and limb retention when injected into mouse ischemic hind limbs (129). Overexpression of adrenomedullin which produces a peptide promoting angiogenesis in MSCs improved capillary density and cardiac function in infarcted rat heart (130). Bone morphogenetic proteins (BMPs) and TGF-β are two important factors that are used generally to promote osteogenesis and chondrogenesis (127).
There are various methods to deliver a therapeutic gene into MSCs. Transcription activator-like effector nuclease (TALEN), zinc finger nuclease (ZFN) and CRISPR/Cas system are some of the significant methods for genetic manipulation in MSCs. These enzymes start the break of double-stranded DNA; following this change, endogenous DNA repair starts and the desired sequence can be inserted into the genome (131). Viral and non-viral methods are used to deliver genes of interest into MSCs. Each method of transduction has its pros and cons. Viral methods yield high transduction efficiency and gene expression for a long period of time. On the other hand, non-viral methods show low transfection efficiency and transgene expression for a shorter period of time. However, they are less toxic and safer and also larger size of genes can be transferred into MSCs by using these methods (127).
Manipulation of MSCs can be a very good method for cell therapy if this modification bring improved survival of MSCs and the target tissues as well (127). Overexpression of BCL-2 in rat BM-MSCs increased survival and enhanced heart tissue repair and organ regeneration after MI. Monkeys and porcine MSCs expressing hTERT gene showed longer lifespans and lessened apoptotic rate with increased proliferation (129). Using viral methods to overexpress Akt in MSCs causes increased survival of these cells. Secreted frizzled-related protein-2 (Sfrp2) is a factor released by Akt-MSCs, and it has a vital role in the survival of ischemic cardiac myocytes (132). Heat shock protein-20 (HSP-20) overexpression and hypoxic preconditioning increased survival of MSCs under ischemic situations. Also, use of the secreted agents as a conditioned medium of HSP-20-MSCs enhanced cardiomyocytes survival under oxidative situations (133).
Migration abilities of MSCs is another parameter that holds great significance in MSC-based therapy. Weak targeting abilities of MSCs is an important barrier which inhibits most of the therapeutic effects of these cells to be exerted on target cells. Genetic manipulation has been used to strengthen the migratory abilities of MSCs. CXCR4-mRNA-transfected MSCs showed significantly enhanced migration toward SDF-1 gradient and improved infarcted myocardium (127). CCR1-expressing MSCs migrated to the site of injury especially in myocardium infarction (134). Since IL-10 and IL-18 from damaged myocardium cause MSCs death, overexpression of IL-18-binding protein in MSCs can lead to protection of them from cell death (135).
Mesenchymal stem cell-secreted factors can have anti-tumor effects on cancerous cells and inspiring results have been obtained from using MSCs soluble factors on glioma, hepatoma, melanoma, breast cancer and lung cancer (136, 137). IFN-α-expressing MSCs increased apoptosis in a melanoma lung metastasis model and thus they induced decrease of cancerous cells proliferation. MSCs expressing CX3CL1, a T cell attractant chemokine, show inhibition of cancerous cells metastasis to lung tissue after injection of cancerous cells. TRAIL-expressing MSCs induced apoptosis in various cancer cell lines (129).
An example of application of ex vivo genetic manipulation is the use of this method in acute lung injury (ALI). Angiotensin II (Ang II) is upregulated in the injured part of lung tissue. Systemic injection of MSC-AT2R (angiotensin II receptor) into ALI mice reduced pulmonary vascular permeability and amended the lung histopathology and had further anti-inflammatory effects (138).
There are several trophic factors released by MSCs that can have good impacts on some neurodegenerative diseases. Glial cell-derived neurotrophic factor (GDNF), brain-derived neurotrophic factor (BDNF), and nerve growth factor (NGF) are therapeutic factors that can improve Parkinson’s disease, Huntington’s disease and Alzheimer’s disease. Genetically modifying MSCs to overexpress these proteins can be useful for better results in therapies of neurodegenerative diseases (Fig. 2) (131).
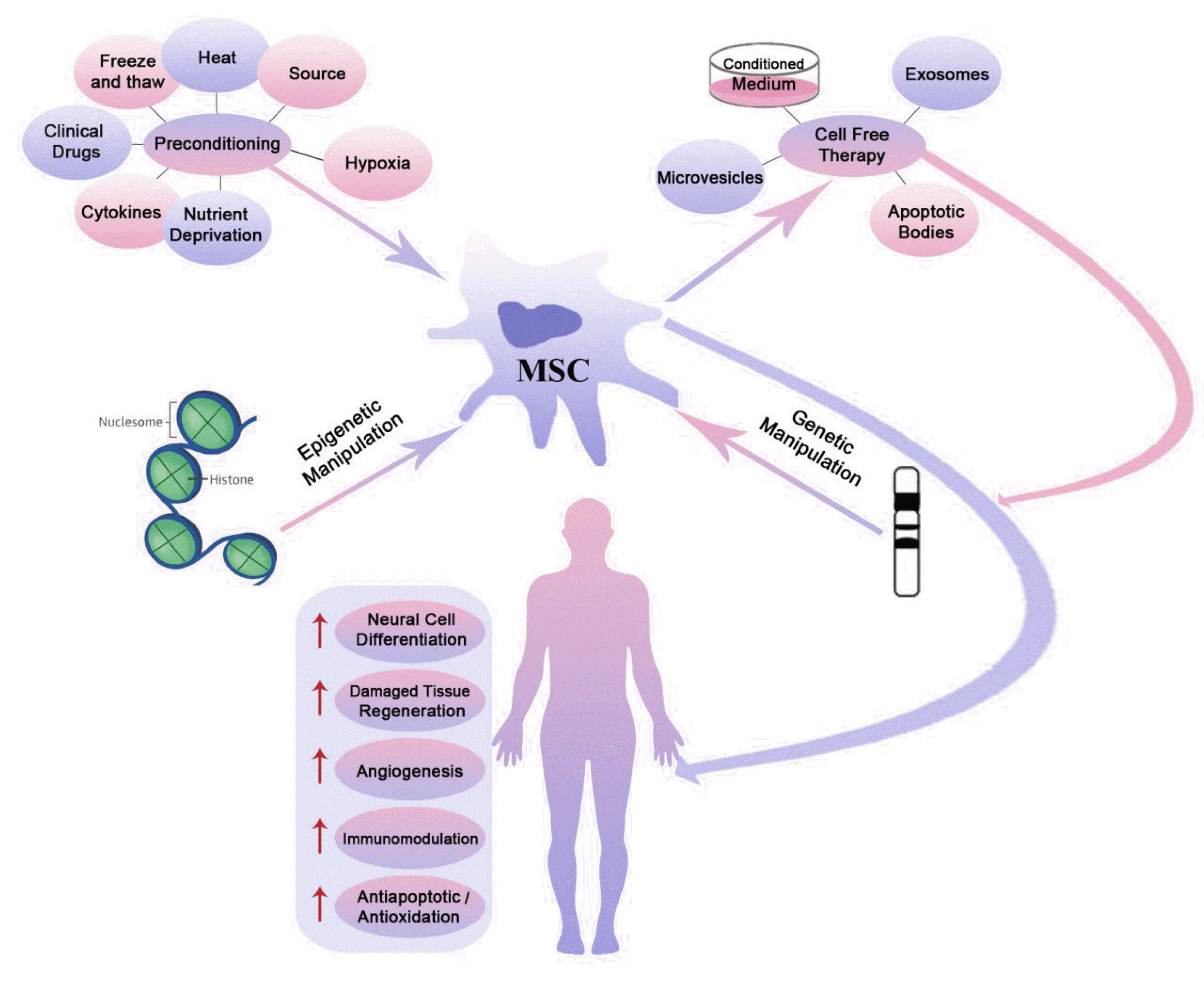 | Fig. 2Modifications of mesenchymal stem cells (MSCs). Preconditioning, genetic and epigenetic manipulation of MSCs and also cell free therapy are key strategies to get better survival, proliferation and function of MSCs. Manipulation of MSCs increase neural cell differentiation, regeneration of damaged tissue, angiogenesis, immunomodulation, and also improve antiapoptotic/antioxidation effects of MSCs which are used in treatment of neurodegenerative diseases.
|
Go to :
Applications of MSCs in the Treatment of Neurodegenerative Diseases
Alzheimer’s disease
Alzheimer’s disease (AD) is the most prevalent neurodegenerative disease which affects approximately 47 million people worldwide. Aging is presumed to be the most important risk factor for AD development. Continuous synaptic damage and neuronal loss, resulting in slow cognitive deterioration and dementia, are the characteristics of the disease. The main microscopic and pathological marks of AD are the amyloid beta and Tau protein agglomerated within the brain (139-141). It is not known for certain whether these plaques and tangles are protective or causative. Decreased levels of acetylcholine, acetylcholinesterase (AChE) and acetyltransferase in the hippocampus and neocortex and reduced levels of neurons in the basal forebrain cholinergic nuclei (BFChN) are the reasons for cholinergic formation hypothesis of the disease (131). Approved drugs for Alzheimer’s disease only impede the progression of the disease and they have tranquilizing effects in almost all of the cases (131, 142). Although preclinical studies of stem cell-based therapies in AD exhibited variable effects, they are mostly associated with decrease in the Aβ load and increase in the Aβ-degrading enzymes. The results gained from stem cell therapy in AD is typically dependent on the types and sources of MSCs. Mesenchymal stem cells exert their therapeutical effects through four main mechanisms which are: 1) restoring the degenerated neurons; 2) neuroprotection through secreted factors; 3) exerting immunomodulatory effects on the cells responsible for the disease development; and 4) proliferation of the endogenous cells (143).
By enhancing the PI3k/Akt pathway, glycogen synthase kinase 3 beta enzyme (GSK3β) plays an important role in AD pathogenesis. Transcriptomic analysis of MSCs especially gingiva-derived MSCs pretreated with cannabidiol (CBD) showed inhibition of GSK3β via binding to TRPV1 receptor. CBD also decreased the expression of genes coding for the kinases and secretases involved in tau protein phosphorylation and Aβ formation, respectively (144). On the other hand, in Alzheimer’s disease, the balance between inflammatory and immune-regulatory responses is dysregulated and a chronic inflammation occurs. It is well indicated that systematic administration of hypoxia-preconditioned MSCs to APP/PS1 mice (AD disease mouse model) induces the secretion of miR-21-enrtiched exosomes, which are important in the control of this balance. This upregulation of miR-21 enhanced memory abilities of mice and Aβ plaque depositions were lessened (145).
Moreover, soluble factors derived from MSCs have shown great therapeutical effects in AD. Activin A, released from MSCs in the coculture of human MSCs with subventricular zone-derived neural cells of 5XFAD mice, caused neural cells to develop and growth (146). Growth differentiation factor-15 (GDF-15) released by neural stem cells (NSCs) in the media of human UCB-MSCs (hUCB-MSCs) enhanced proliferation of NSCs in hUCB-MSCs-NSCs coculture (147). Transplantation of MSCs to the brains of AD mice led to the release of CCL5 from BM-MSCs. In fact, the amyloid beta-enriched regions stimulate the secretion of CCL5, the attracting chemokine released by microglia cells to help the phagocytosis and removal of Aβ plaques. Memory function was enhanced by IL-4 and neprilysin (NEP) production of alternative microglia. NEP is a zinc-dependent metalloprotease that degrades Aβ plaques (148). Administration of microglial cells coupled with recombinant ICAM-1 into the mice brain causes the expression of NEP in dose and time-dependent manner as well (149). Proteasome complex activity is downregulated in Alzheimer’s disease and it results in the accumulation of ubiquitin-conjugated proteins. Lee et al. (150) showed that agouti-related peptide (AgRP) released from human MSCs enhance the proteasome complex activity and administration of human MSCs or the AgRP itself to the hippocampus of 5XFAD mice can decrease the accumulation of unwanted proteins in the brain. Thrombospondin (TSP-1) is another secreted molecule from hUCB-MSCs to relieve synaptic dysfunction, caused by Aβ peptides (151).
Genetic manipulation of MSCs has yielded promising results in enhancing MSCs effects. MSCs increase neurogenesis and differentiation of neural progenitor cells into mature neurons by enhancing the Wnt pathway (152). The exosomes and microvesicles released from modified MSCs are enriched in factors of interest to enhance the therapeutic effects. Overexpression of Aβ degrading enzymes or downregulation of secretase enzymes related to Aβ formation with transfecting different RNAs can be useful in ameliorating disease symptoms (153). Brn-4 protein in MSCs can inhibit the accumulation of Aβ in the brain but miR-937 presence impedes the translation of Brn-4 mRNA to protein. Administration of (Antisense)-miR-937-MSCs helped in Aβ deposition reduction and clearance in the brain (154, 155). Overexpression of BDNF in MSCs extended survival and increased synaptic markers in neurons of 5XFAD mouse, and APP/PS1 transgenic mouse model (156, 157).
Parkinson’s disease
After AD, Parkinson’s disease (PD) is identified as the second most common neurodegenerative disease, which causes severe movement disorder, tremor, rigidity, and postural instability (158, 159). Presence of abnormal alpha-synuclein protein aggregates inside the brain is the most important neuropathological characteristic of the disease (160). Gait difficulties in PD are because of gradual degeneration of neurons in the substantia nigra. The main responsibility of these neurons is to produce dopamine to keep balance in motor activities (161). Most of the treatments for Parkinson’s disease are about the restoration of dopamine inside the brain but too many side effects of these treatments have prompted researchers to look for alternative therapeutical agents. One promising target is the use of regenerative medicine especially stem cells in the amelioration and treatment of the disease (158, 161).
Mesenchymal stem cells play an important role in blocking of α-synuclein transmission. MSCs do this through interaction with N-methyl-D-aspartate receptors and inhibition of α-synuclein endocytosis. Surveys have shown that secreted factor “galectin-1” has an important involvement in the control of transmission of α-synuclein (162, 163).
As discussed previously, pretreatment of MSCs with different conditions and agents can enhance the therapeutic actions of MSCs. This method has shown great results in the therapy of PD as well. Hypoxic preconditioning of MSCs (3% O2 treatment) in the process of neural differentiation has led to an increase in HIF-1α expression, phosphorylation of P38 MAPK and Erk. These changes in the genetic profile of MSCs resulted in enhanced neurosphere generation, dopaminergic neuronal differentiation alongside increased differentiation of neuronal cells. Neurons generated by hypoxic preconditioning exhibit elongated survival and can be more therapeutic than ordinary neurons (164). PC12 cells (a cell model of PD) affected by the supernatant of curcumin pretreated hUC-MSCs showed enhanced proliferation and elevated expression of microtubule associated protein-2 (MAP-2) and tyrosin hydroxylase, a rate-limiting enzyme in dopamine synthesis. These effects were not seen to this extent when using the supernatants of hUC-MSCs alone. This shows that preconditioning of MSCs by curcumin can enhance their therapeutic applications in PD (165).
Using the secreted agents from MSCs as cell-free therapy can have beneficial effects on Parkinson’s disease as well. ROS generation is one of the pathological hallmarks of PD. Neuroblastoma SH-SY5Y cells were pretreated with the conditioned medium extracted from rat adipose tissue-derived MSCs (AT-MSCs) to check MSCs conditioned media’s effect on pro-oxidizing agents. Neuroblastoma cells cocultured with conditioned media showed more decreased mortality and ROS generation when treated with H2O2 or 6-OHDA (6-hydroxydopamine), a dopaminergic selective toxin; than Neuroblastoma cells not preconditioned with conditioned media (166). MSCs conditioned medium is a good therapeutic candidate as neural stem cells, pretreated with conditioned medium (CM-NSCs) showed increased survival and enhanced migratory activity; but NSCs alone lacked these extra characteristics (167).
Genetically engineered MSCs exhibit increased functional and therapeutical activities. Conserved dopamine neurotrophic factor (CDNF)-expressing BM-MSCs can reduce 6-OHDA induced neurotoxicity on dopaminergic neurons and enhance behavioral recovery of these cells (168). Another gene responsible for CNS tissues protection is neurotrophin-3 (nt-3gene) which enhances neuronal-like differentiation. Overexpression of this gene in MSCs promotes the expression of tyrosine hydroxylase (TH), an indicator of the intracellular signaling in dopaminergic neuron differentiation (169).
Adenoviral and lentiviral delivery of specific genes into MSCs could have good results on PD therapy. Hepatocyte growth factor expressing MSCs enhance recreation of damaged neurons. They also express LMX1a, an essential participant in dopaminergic differentiation, which increase tyrosine hydroxylase and dopaminergic genes expression (170, 171).
Huntington’s disease
Huntington’s disease (HD) is the most prevalent monogenic neurodegenerative disease which is inherited in an autosomal dominant pattern and occurs when polyglutamine (CAG) repeats exist in the first exon of Huntingtin gene, located on the short arm of the fourth chromosome. The mutation inside this gene leads to bad protein production which ultimately causes the symptoms of the disease such as choreic movement and troubles in cognitive and emotional feelings. HD leads to death in most of the patients (131, 172, 173). Lack of a suitable conventional therapy is a characteristic shared between most of the neurodegenerative diseases such as HD. Mesenchymal stem cells have been tested on HD because of high differentiation capacity to neuronal cells and ameliorating effects of them (174).
MSCs secrete a wide range of neurotrophic factors, show anti-inflammatory effects and are less vulnerable to getting rejected. Therefore, they have shown enough capability to slow the beginning of neurodegenerative deficiencies and behavioral incompetence. Neuropathological symptoms of HD in rodent models relieved after intrastriatal transplantation of MSCs (175, 176). The decrease in neurotrophic factors (NTFs) can have an important involvement in HD development. Specific procedures have been developed to induce BM-MSCs to secret NTFs. Transplantation of these NTFs-secreting MSCs to the brains of rats after quinolinic acid (QA) injection, as a model for HD, decreased striatal mass changes and had beneficial therapeutic effects (177). Human BM-MSCs can integrate with the host cells to increase the level of some secreted factors like von willebrand factor (VWF), SDF-1 and its receptor CXCR4. Transplanted MSCs can decrease the expression of apoptotic factors, including Bax and Caspase-3, and increase P-Erk1/2 expression (178).
Preconditioning of MSCs with lithium and valproic acid, commonly used as mood stabilizers, can improve the therapeutic efficacy of these cells. Preconditioned MSCs lessened HD aggregates in HD mice and hampered striatal neuronal loss. These MSCs exhibited intragenic effects such as an enhanced expression of antioxidants, migration-related genes, and anti-apoptosis molecules. This level of gene expression profile was not seen when using unpreconditioned cells (75).
Because genetic profile has an important role in the formation of HD, genetic manipulation of MSCs can be really useful in HD therapy. MSCs can be used to transfer specific interfering RNAs (RNAi) which can target mutant and defective huntingtin in neuronal cells (179). Genetically manipulating MSCs to overexpress neural growth factors (ex: BDNF and GDNF) and transplanting these cells through intrathecal or intracerebral way can exert neuroprotective effects. They can reduce both free radicals levels and apoptosis in neuronal cells. BDNF can create a situation inside the striatum that slows neurodegeneration in the YAC128 HD mouse model (Table 1) (180, 181).
Table 1
The list of therapeutic applications of MSCs in Alzheimer disease (AD), Parkinson’s disease (PD), and Huntington’s disease (HD)
| Disease | Applications of MSCs | References |
|---|---|---|
| AD | ∙ Modulation of the expression of AD-related genes in gingiva-derived MSCs by CBD. | (144) |
| ∙ Amelioration of cognitive decline by rescuing synaptic dysfunction and regulating inflammatory responses in APP/PS1 mice through miR-21 enriched exosomes derived from hypoxia-preconditioned MSCs. | (145) | |
| ∙ Neural development and neurite outgrowth induced by Activin A, secreted by human MSCs in an in vitro model of AD. | (146) | |
| ∙ Amelioration of AD mice through soluble CCL5 produced from bone marrow-derived MSCs by recruiting bone marrow-induced microglia immune responses, including phagocytosis, and IL-4 and NEP production. | (148) | |
| ∙ AgRP released from human MSCs upregulates proteasome activity in 5XFAD mice, an AD model. | (150) | |
| ∙ The rescue of neurons from synaptic dysfunction through thrombospondin-1 secreted by human umbilical cord blood-derived MSCs in AD model. | (151) | |
| ∙ Administration of antisense-miR-937-MSCs to help to decrease and clearance of Aβ deposition in the brain through inhibiting the translation of Brn-4 mRNA. | (154, 155) | |
| ∙ Extended survival and increased synaptic markers in neurons of 5XFAD mouse, and APP/PS1 transgenic mouse model by overexpression of BDNF in MSCs. | (156, 157) | |
| PD | ∙ Blocking of α-synuclein transmission by MSCs through inhibition of α-synuclein endocytosis and galectin-1 secretion in PD model. | (162, 163) |
| ∙ Hypoxia preconditioning of MSCs for promoting dopaminergic differentiation of MSCs and elongated survival of neurons in a rat model of PD. | (164) | |
| ∙ Preconditioning of MSCs by curcumin on 1-Methyl-4-phenylpyridine-Induced PD Cell Model lead to enhanced proliferation and elevated expression of MAP-2 and tyrosin hydroxylase. | (165) | |
| ∙ MSC-Secreted Factors and Neural Stem Cell coculture and Transplantation increase Recovery of PD Rat model. | (167) | |
| ∙ Genetically engineered CDNF-expressing MSCs reduced 6-OHDA induced neurotoxicity on dopaminergic neurons. | (168) | |
| ∙ Brain tissue engineering through combination of NT3-overexpressing MSCs and PLGA microcarriers as a promising tool for treatment of PD. | (169) | |
| ∙ Lentiviral delivery of LMX1a and HGF into human bone marrow MSCs for PD therapy. | (170, 171) | |
| HD | ∙ A potential therapy for HD rat model through transplantation of NTF-secreting MSCs to the brains of Rats after QA injection. | (177) |
| ∙ Transplanted human BM-MSCs prolong survival and ameliorate motor deficit through decrease of Bax and caspase3 and increase of Erk1/2 in HD mouse models. | (178) | |
| ∙ Lessening of HD aggregates in HD mice by preconditioning MSCs with lithium and VPA exhibiting intragenic effects like an enhanced expression of antioxidants, migration-related genes, and anti-apoptosis molecules. | (75) | |
| ∙ Genetically manipulating of MSCs to transfer specific RNAi targeting mutant and defective huntingtin in neuronal cells or overexpressing BDNF and GDNF exerting neuroprotective effects. | (179∼181) |
AD: Alzheimer’s disease, PD: Parkinson’s disease; HD: Huntington’s disease, MSCs: Mesenchymal stem cells, CBD: Cannabidiol, AgRP: Agouti-related peptide, BDNF: Brain derived neurotrophic factor, CDNF: Cerebral dopamine neurotrophic factor, 6-OHDA: 6-hydroxydopamine, NT3: Neurotrophin-3, HGF: Hepatocyte growth factor, NTF: Neurotrophic factor, QA: Quinolinic acid, VPA: Valproic acid.
![]()
Go to :
Conclusions
Neurodegenerative diseases impose a huge burden on every aspect of life. The fact that there is no proper treatment for most of these diseases, has become a great incentive for researchers to focus on regenerative medicine. In this field, the use of mesenchymal stem cells has yielded good results. MSCs are multipotent cells that have self-renewal, neuroprotective and regenerative abilities and can differentiate into a number of cell types which makes them perfect candidates for regenerative medicine. MSCs have a lot of therapeutic applications as they can have modulatory effects on almost all of immune cells. Therapeutic applications of MSCs can be strengthened by manipulating different aspects of them. MSCs can be pretreated with different subjects, manipulated genetically or cultured to take their secretome.
Go to :
 XML Download
XML Download