

 PDF
PDF Citation
Citation Print
Print


Introduction
Hematopoietic stem cells (HSCs) are multipotent primitive cells that can develop into all types of blood cells, including myeloid-lineage and lymphoid-lineage cells (1). HSCs can be found in several organs, such as peripheral blood (PB), bone marrow (BM), and umbilical cord blood (UCB). All blood cell lineages are produced via functional maturation of a rare population of multipotent HSCs that can proliferate by self-renewal and differentiation. Thus, understanding the molecular mechanisms regulating the self-renewal and cell fate determination of HSCs/progenitor cells is important for the development of clinical applications based on disease type and severity. Only a small population of HSCs is required to initiate the entire hematopoietic process. Since Cheng et al. (2) first reported that HSC quiescence is maintained by p21cip1/waf1 using p21−/− mice, numerous papers have found that long term-hematopoietic stem cells (LT-HSC) quiescence and activation are regulated by the genetic and epigenetic regulation of key molecules as well as by microenvironmental factors (3, 4). In addition to the attribute of self-renewal, a characteristic that distinguishes HSCs from other mature cells is the ability to undergo specific and large-scale differentiation into cells of various lineages. These findings from a colony-forming unit (CFU) assay also revealed HSC activity, and further measured the capacity of HSCs to produce mature blood cells in humans following myeloablation. Similar with LT-HSC, numerous papers has been studied the properties of short term HSC (ST-HSC) and progenitors and addressed that HSC/progenitors can directly or indirectly contributes in regenerating tissues and blood disorders. Based on these literatures, we described the available sources of HSCs, aging HSCs, and plasticity of HSCs in scientific information and clinical application of HSCs in blood disorders and liver deficiency.
Go to : 
Definition of Hematopoietic Stem Cells (HSCs)
All lineage blood cells are produced by a rare population of multipotent HSC, which can proliferate by self-renewal and differentiate to accomplish the functional maturation. From HSC to mature cells, there are several intermediate progenitor cells (5). These cells can display both functional multipotent and lineage-committed properties simultaneously or separately prior to complete maturation. Blood is a highly regenerative tissue due to its short life span, and BM supports the dynamic movement of diverse cells to maintain homeostasis of blood cells. Approximately, one trillion cells are generated daily to compensate apoptotic cells in human BM, suggesting rapid circulation of blood cells. HSC can be divided into LT-HSC, ST-HSC, and multipotent progenitor (MPP) in terms of duration of repopulation (6). In normal physiological conditions, rare HSC populations such as LT-HSC can develop into all lineage blood cells in the BM. However, HSC populations in the PB tend to be higher in myelosuppressive conditions caused by drug and granulocyte colony-stimulating factor (G-CSF), and rapidly migrate from the BM, suggesting different properties. HSC/progenitor cells in the PB are ST-HSC, which might directly contribute to recovering damaged tissues, and these are regarded as optimal curative cell sources in regenerative medicine. Blood cells from HSCs are divided into two lineages: lymphoid cells and myeloid cells (7). The lymphoid branch consists of T, B, and natural killer (NK) cells, which are relevant to innate and adaptive immune cells. This process is known lymphopoiesis. Myeloid lineage cells include all blood cells except lymphoid cells. There are several types of cells including monocytes from monoblasts, erythrocytes from erythroblasts, platelets from megakaryocytes, and granulocytes, which consist of neutrophils, eosinophils, and basophils, and are from myleoblasts. Lineage committed progenitor cells are specified for cell fate and develop into mature cells through myelopoiesis. In myeloid lineage cells, committed myeloid progenitors can be converted into mature types of myeloid cells. Lymphoid cells also have a similar process as the myeloid cells for generating progenitor cells and lymphoblast are from committed lymphoid progenitor cells. These blood cells are affected by dynamic niche composition including hormone and pericytes in BM (8). Similar to stem cells, progenitor cells in each step can also function as a strong cell source in regenerative medicine (Fig. 1). Thus, understanding HSC/progenitor cells is important to apply these cells in regenerative medicine depending on the disease type and severity. Thomas et al. (9, 10) reported that human BM cells infused into irradiation and chemotherapy receiving patients, resulted in the recovery of white blood cells and increased hemoglobin level. Two decades ago, existence HSC/progenitor cells in vivo is proved by advanced technology such as retroviral skill and clinic application using HSC has been continuously developed with exclusive demand. Remarkably, Till and Mcculloch (1) showed a colony forming unit (CFU) assay that can quantify HSC/progenitor cells; because HSC/primitive progenitor cells can repopulate through self-renewal, regenerative HSCs can be assayed with clonal expansion to generate multipotent HSCs. Moreover, this method further motivated scientist to develop ex vivo expansion for HSCs. Acquiring high number of HSCs/progenitors is a critical issue in regenerative medicine in order to overcome the limitations of cell sources. Besides attributes of self-renewal, a distinguishing feature of HSC aggregation from other mature cells is its specific differentiation with huge numbers into various lineage cells. These findings from the CFU assay also revealed LT-HSC activity, which further quantified HSC in producing mature blood cells in patients with myeloablation. Ultimately, we know that self-renewal proliferation of HSCs and differentiation into committed cell lineages through intermediate stages including progenitors are pivotal properties of HSCs. Because rare populations of HSC can lead the entire hematopoiesis, knowing the properties of HSC is very important to enhance therapeutic effects in clinical application. Recently, aging is emerging as an issue in HSC biology. Since Morrison et al. (11) mentioned homing and engraftment of HSCs based on aging, many papers have demonstrated the relevance of aging by DNA damage in HSC and molecules such as ATR gene and Cdc42 are pivotal to maintain phenotypes without HSC loss and back to rejuvenation (12). The immune system is orchestrated in terms of coordination of adaptive immune cells by lymphoid lineage cells including T and B cells and innate immune cells by myeloid cells. During aging, there is notable reduction in lymphocytes, especially naïve T cells, which are apparent, as well as the accumulation and clonal expansion of memory T cells (13). Meanwhile, myeloid lineage cells in BM seem to be increased or persist at the same level in an age-dependent manner, which leads to a proinflammatory environment (14). Thus, identification and isolation of bona fide HSCs with specific indicators should be demanded prior to experiments to address HSCs by aging. Until now, identification of HSCs using CD markers has advanced and the technique for isolation with high purity has been improved.
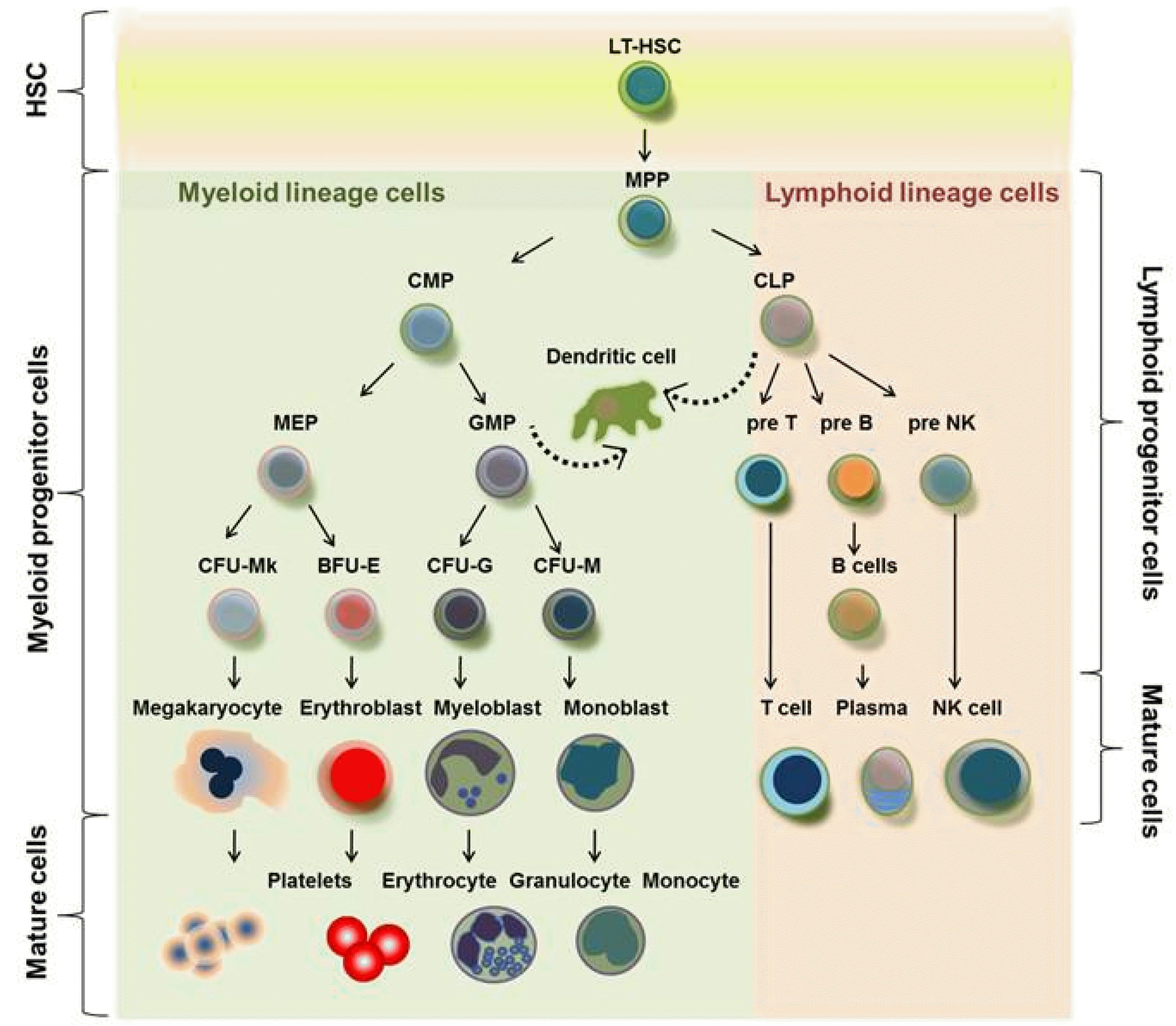 | Fig. 1Schematic illustration showed hematopoiesis in adult BM. HSCs have self-renewal activity and differentiation capability into blood cell lineages. LT-HSCs are quiescent and sustained by arrested cell division in endosteal niche, whereas ST-HSCs are promptly differentiated into myeloid and lymphoid lineage cells in vascular niche, and then lineage cells exit into periphery.
|
Go to :
Aging of Hematopoietic Stem Cells
Research reports have consistently showed that understanding the properties of HSCs is very important for enhancing their therapeutic effects in clinical applications. Aging is relevant to the functional decline of normal HSCs as well as increased risk of hematologic malignancies (15). Abnormal clonal hematopoiesis which frequently occurs in the HSCs of the elderly is caused by DNA mutation in specific loci and shortening telomerase (16). Besides, there are several emerging intrinsic factors such as increased polarity, reactive oxygen species, and injured autophagy and mitochondria metabolism as ample evidence of aging of HSCs (17), which show the main features of aged HSCs compared to that of young HSCs. Especially, Mohrin et al. (17) showed the reversible contributing factor, deregulation of a UPR(mt)-mediated metabolic checkpoint, in HSC aging, which suggested that mitochondria metabolism may play a role to govern juvenescence of HSCs (12). Although these evidence remain unclear, pharmacological and metabolic interventions in these pathways may be continuously exploited to restore function in aged HSCs. Epigenetic reprogramming as a part of intrinsic regulation is one of the main factors in aging of HSCs. Although epigenetic abnormality is easily detected, correlation between aged HSC and epigenetic aberrancy is unreadable due to the slow loss of normal stem cell potential. Epigenetic regulation by TET2, DNMT3, and EZH2 led to hematologic malignant transformation, implying the faithful role of epigenetic control in the aging of HSCs (18). Epigenetic fidelity in a normal stem cell niche is required to maintain normal HSCs, because aging of HSCs is driven by a strong contribution of aged niche (19). Aged BM has two- to ten-fold HSCs when compared to that of the young BM (11). Multiple myeloma (MM) is a clonal plasma cell malignancy from BM failure by epigenetic defects (20). Thus, an epigenetic modulating agent such as decitabine is used to enhance therapeutic efficacy in MM. Regardless of aberrant epigenetic defects in the niche as well as autonomous HSCs, gene mutations may ultimately alter the epigenetic memory of HSCs, and then clonally expand with mutant epigenetic memory leading to abnormal hematopoiesis. It shows that modulation of epigenetic agents synergistically enhances clinical drug responses and normalization of abnormal mutation from aging status. The aging of HSCs depends on cellular changes, such as epigenetic factors, telomere and genomic damage, and molecular damage, including DNA damage, and finally resulting in dysfunctional HSCs (15). Notably, the immune system is regulated via the coordination of adaptive immune cells by lymphoid lineage cells, including T cells and B cells, and of innate immune cells by myeloid cells. Aging leads to a reduction in lymphocytes, especially naïve T cells, and the accumulation and clonal expansion of memory T cells (13). Meanwhile, myeloid lineage cells in BM appear to increase or persist at the same level depending on age, leading to a proinflammatory environment (14, 21). There are two models of HSC aging; one is a clonal alteration model, in which all lymphoid cells can temporarily be converted into myeloid cells due to aging via a differentiation stage. The other is a population shift model, in which myeloid-biased stem cells exclusively expand in old BM but not young BM, thus lymphoid-biased stem cell populations are overwhelmingly decreased, resulting in an imbalance (17, 21). As previously mentioned, there is some debate regarding the properties of aged HSCs in humans, and analysis of HSC characteristics during aging remains inconclusive. Pang et al. (22) insisted that aged HSCs do not engraft with increased homing abilities or differentiate into blood lineage cells with decreased efficiency compared to young HSCs. In contrast, increased proliferative activity has been reported by Kuranda et al. (23) in aged HSCs compared to young HSCs. Because both studies used different HSC markers to investigate the characteristics of aged and young HSCs, there have been inconsistencies after transplantation. Thus, the identification and isolation of bona fide HSCs are required prior to investigating aging HSCs, which can be achieved by applying recently developed advanced isolation techniques that ensure high purity, and using improved HSC-specific markers.
Go to :
Hematopoietic Stem Cell Transplantation (HSCT) for Blood Disorders
HSCT involves the intravenous infusion of autologous or allogeneic stem cells collected from BM, PB, and USB to reconstruct a functional hematopoietic system in patients with blood disorders. This process is used to allow a person to receive high-dose chemotherapy to treat a disease. Globally, 53% and 47% of HSCT is autologous and allogeneic, respectively, per year, and almost 68.8% of total 854 patients (Acute myeloid leukemia (AML) 158, acute lymphoblastic leukemia (ALL) 59, non-Hodgkin lymphoma (NHL) 392, Hodgkin disease (HD) 245) are alive 10 years after autologous HSCT (24). Cell therapy using HSCs was first performed more than 60 years ago (25). In humans, HSCs were identified and cultured in the 1980s (26). HSCT is one representative of many revolutionary solutions using HSCs, including cell and gene therapy. Among many other diseases, leukemia especially requires HSC therapy. AML is an aggressive malignancy with high mortality; it has been considered the worst blood cancer. For all AML types except M3, which is known as acute promyelocytic leukemia, HSCT is commonly conducted post-chemotherapy; the ultimate goal is the restoration of functional immune cells. HSCT prolongs life and can be divided into two main methods: autologous and allogeneic transplantation. Both methods of HSCT are regarded as supportive care and are an established therapy in many cases; in fact, HSCT has been integrated into the therapeutic strategy of most large, multicenter, cooperative study group trials. Recently, studies on the effectiveness of autologous HSCT in older adults have included patients harboring multiple types of hematologic malignancies; for example, autologous HSCT has been used as a therapeutic option among patients with both multiple myeloma (MM) and lymphoma (27). Among hematologic malignancies, MM is the most common indication for autologous HSCT, and overall survival is prolonged among patients treated with autologous HSCT compared to those treated with only conventional chemotherapy. Of note, autologous HSCT during the first complete remission is not superior to the continuation of chemotherapy in any of the reviewed trials. Although the debate is ongoing for AML, autologous transplantation in AML patients is still challenging due to the severe heterogeneity of AML and graft versus host disease (GVHD). Meanwhile, allogeneic HSCT is more popular for many types of diseases, including malignant and non-malignant disorders. For allogeneic HSCT, it is important to achieve a high degree of human leukocyte antigen (HLA) matching between the donor and recipient. Graft immune rejection and GVHD are remarkably decreased when well-matched HSCs are transplanted. To achieve this, the HSC donor should be investigated in diverse clinical aspects, especially hematologic condition and HLA typing. Allogenic HSCT is used to treat the following disorders: leukemia, including AML, ALL, chronic myeloid leukemia (CML), and chronic lymphocytic leukemia (CLL); myeloproliferative disorders; MM, NHL; and anemia. Young CLL patients are able to undergo both autologous and allogeneic HSCT. Both NHL and HD patients, who failed to achieve complete remission by conventional therapy, are used to high dose chemotherapy with HSCT. This treatment can successfully induce complete remission in relapsed patients (28). Anemia is a group of diseases involving red blood cells. Among them, thalassemia major patients need blood transfusions; however, blood transfusion is an insufficient treatment due to the presence of the genetic problems associated with the disease (29). HSCT treatment has undergone advancements over the last two decades, and 80% disease-free survival has been reported with allogeneic HSCT (30). Representative therapeutic approaches, such as HSCT, have been improved; furthermore, improved techniques, such as gene transfer with HSCT, could be used to treat a variety of diseases (31).
Go to :
Contribution of HSCs/Progenitor Cells to Regeneration of Non-Hematopoietic Cells
HSCT is feasible in myeloablated and non-myeloablated hosts, leading to a chimeric condition when allogeneic HSCT is carried out. In addition to using HSCT to induce hematopoietic cell reconstruction, cell therapy using BM-derived stem/progenitor cells has been continuously applied in regenerative medicine. Additionally, tissue-specific stem/progenitor cells in hematopoietic and non-hematopoietic tissues are also regarded as important therapeutic cell sources. BM-derived stem/progenitor cells are able to differentiate into cells of other lineages in a process called transdifferentiation, as well as differentiation (32). Many reports have shown that HSCs can transdifferentiate into cells of other lineages, such as endothelial cells, cardiomyocytes, neural cells, and hepatocytes (33, 34). These results seem to contradict the common assumptions of cell origins from the three germ layers and question whether HSCs are multipotent cells that can transdifferentiate or whether they are more primitive functional cells. However, since the mid-2000s, refutations of the plasticity of HSCs are emerging in regenerative medicine fields, suggesting that there is only a limited population of HSCs under physiological conditions (35). Stem/progenitor cells are needed to regenerate organs, and interest in stem/progenitor cells has been fueled by a serious need for lineage-specific progenitor cells for the treatment of diseases. Additionally, despite the rarity of stem/progenitor cells in regenerated organs, it is now believed that diverse effects from stem/progenitor cells other than direct cell contribution, such as paracrine effects, cell recruitment, and microenvironmental modulation, can repair injured organs. In particular, liver cells, which are from an endodermal lineage, rapidly acquire noticeable cellular functions and regenerative capacity. Because all changes in the liver are elicited by highly conserved intrinsic signals and genetic factors, the activity of both HSCs and oval progenitor cells in the liver is extremely relevant to developmental biology. Studies using animal disease models are leading to the development of methods to derive hepatocyte and hepatic oval cells from diverse stem/progenitor cell types (36). Among progenitor cells, endothelial progenitor cells (EPCs) represent progenitor cells that are derived from blood lineage cells; EPCs were first reported by Asahara et al. (32). Later, EPCs were studied by many investigators in regenerative medicine (37). Similar to HSCs, the ability to transdifferentiate EPCs into cells of other lineages has also been continuously debated (32). To enhance the use of stem/progenitor cells in therapeutic approaches, understanding how intrinsic regenerative capacities can be stimulated will help facilitate the control of cellular plasticity in stem/progenitor cells. Although the direct transdifferentiation of HSCs or BM-derived progenitor cells to regenerative organs has been continuously debated and does not seem to happen in vivo, HSCs/progenitor cells are still considered important elements in injury repair in terms of disease severity and organ types (38-49). We have briefly summarized the contribution by the plasticity of HSCs/progenitors to non-hematopoietic tissue regeneration in Table 1.
Table 1
Contribution of HSCs/progenitor cells in regeneration of non-hematopoietic cells
| Source of stem cells | Differentiation of HSCs/progenitors | Year | Reference | |
|---|---|---|---|---|
| 1 | BM-hematopoietic cells | Microglia/Macroglia | 1997 | (38) |
| 2 | BM-myogenic progenitors | Muscle cells | 1998 | (39) |
| 3 | BM-Stem cells | Endothelial precursors | 1999 | (40) |
| 4 | BM-Stem cells | Hepatic oval cells | 1999 | (41) |
| 5 | BM-Stem cells | Neuronal cells | 2000 | (42) |
| 6 | BM-Stem cells | Hepatocytes | 2000 | (43) |
| 7 | Hematopoietic stem cells | hepatocytes | 2000 | (34) |
| 8 | BM-Stem cells | Cardiac muscle | 2001 | (44) |
| 9 | BM-side population cells | Cardiac muscle, endothelium | 2001 | (45) |
| 10 | PB-stem cells | Hepatocytes/epithelial cells | 2002 | (46) |
| 11 | BM-Stem cells | Cardiac muscle | 2005 | (47) |
| 12 | BM-Stem cells | Lymphatic endothelial progenitors | 2010 | (48) |
| 13 | Corticalbone-derived stemcell | Myocytes/vascular cells | 2013 | (49) |
![]()
Go to :
HSCs in Liver Regeneration
Liver regeneration is a fundamental response of the liver to injury. Similar to other organs, it is well known that local preexisting hepatic progenitor cells and circulating BM-derived progenitor cells are major contributors during liver regeneration (50). All elements are orchestrated, and then complete liver regeneration is accomplished. It has been shown that hepatocyte proliferation mainly contributes to liver regeneration and is regulated by various factors and cytokines, such as hepatocyte growth factor, epidermal growth factor, and interleukin-6 (51). When hepatocytes cannot function due to factors such as toxins and carcinogens, hepatic progenitor cells can be found in the injured liver. Liver progenitors/oval cells are very small and contain little cytoplasm and oval nuclei. Although preexisting mature hepatocytes are the quickest and most efficient cell sources for liver repair, their renewal capacity is exhausted and impaired under some pathological conditions. Thus, hepatic stem/progenitor cells, rather than mature hepatocytes, actively proliferate and differentiate into hepatocytes. This phenomenon occurs in both mice and humans (52). Clinically, liver transplantation is the only available therapy for terminal liver failure. However, the greatest impediment is finding a suitable HLA-matched donor. Therefore, there has been a major effort to develop alternative methods or sources to improve liver regeneration. Increasing evidence suggests that BM-derived stem/progenitor cells can differentiate into hepatocytes, thus resulting in the accumulation of hepatocytes (53). Starzl (54) performed the first liver transplantation and attempted five more transplantations in 1963. After that, the mixed chimeric status of blood cells in the liver after transplantation was reported by Taniguchi et al. in 1996 (53). Hepatocytes can be found in the transplanted livers of animals and humans, and transplantation of reduced-size liver grafts accelerates the recruitment and regeneration of these cells (50). This paper showed that BM-derived CD34+ cells can be directly incorporated into newly regenerated liver following liver transplantation in humans, suggesting the transdifferentiation of BM-derived stem/progenitor cells into hepatocytes. Although liver cells are derived from the endoderm and blood cells are derived from the mesoderm, the concept of transdifferentiation is emerging in liver regeneration fields. In mice, the adult liver consists of c-kit+Sca-1+Linlow/− cells, which are defined as HSCs, and these cells can form hematopoietic colonies in vivo and in vitro (53), which shows the intimate relationship between hematopoietic cells and hepatocytes. Although hepatic lineage cells are totally different from hematopoietic lineage cells, Petersen et al. also showed that hepatic oval cells can express the Thy-1 HSC marker when hepatic injury is induced by 2-acetylaminofluorene. This finding indicates that the Thy-1+ oval cell population could be used to study hepatic oval cells. Theise et al. (55) reported that BM cells can transdifferentiate into hepatocytes in irradiated mice and humans. Based on this concept, Lagasse et al. (34) tried to isolate purified HSCs and differentiate them into hepatocytes, resulting in a high proportion of bone marrow stem cell (BMSC)-derived hepatocytes following strong positive HSC selection. Several studies have suggested that hepatic stem/progenitor cells possess the phenotype of a side population of blood cells that has the ability to efflux Hoechst 33342. This side population of cells among liver cells is similar to hematopoietic lineage cells and expresses the CD34 marker. When hepatic stem cells are activated, BCRP1 (ABCG2) mRNA usually increases in liver tissue. Because BCRP1 is one of the determinants of the side population, its detection indicates an increase in hepatic stem cells with the side population phenotype (56). Most experiments that address the contribution of HSC-derived cells in liver regeneration use a mouse injury model and have shown that liver regeneration is caused by the clonal expansion of multipotent HSCs (33, 34, 50). Krause et al. (33) reported that non-hepatic adult stem cells can differentiate into hepatocytes in humans. In addition, the differentiation capacity of human HSCs into cells of other lineages, including skin, gut, and liver cells, has been reported (57). Human biopsy liver samples have shown that circulating blood stem cells differentiate into mature hepatocytes and epithelial cells via sex-mismatched transplantation. As mentioned previously, all extrahepatic stem/progenitor cells contribute to liver regeneration. However, the low efficiency (less than 0.01%) of extrahepatic stem/progenitor cell contribution post HSCT has been a matter of debate. Most scientists think that extrahepatic cells, such as BM-derived HSCs/progenitor cells, fuse with host hepatocytes rather than transdifferentiate, which requires confirmation by cytogenetic analysis (58). However, while it is relatively easy to obtain samples from mouse models, acquiring a transplanted liver sample from humans is very difficult. Nevertheless, all evidence suggests that cell fusion occurs rather than BM-derived HSCs/progenitor cell transdifferentiation. However, Lee et al. (50) clearly showed transdifferentiation of BM-derived CD34+ stem/progenitor cells in human liver transplants. Despite the different levels of contribution, BM-derived HSCs/progenitor cells might be a suitable tool for addressing some liver diseases, such as metabolic defects, via fusion-mediated additive gene transfer. Additionally, because de novo regenerated liver is very important for transplant survival in humans, the contribution of BM-derived HSCs is also considered important in clinical practice. Many clinical studies have suggested the implementation of HSC and hepatocyte transplantation (59). Clinical approaches using BM-derived cells, including mononuclear cells (MNCs), CD34+ stem cells, and mesenchymal stem/stromal cells (MSCs), have been attempted for the treatment of liver diseases (60-83). We expect that multipotent BM-stem/progenitor cells have beneficial effects and can rescue liver cells via cell transdifferentiation, paracrine effects, anti-fibrotic effects, and proangiogenic events. So far, there have been no unexpected severe side effects. However, the potentially increased risk of hepatic disease associated with HSC therapy requires long-term monitoring in clinical trials. In Table 2, we have summarized the reports on the use of HSC therapy.
Table 2
Clinical trials on stem cell therapy in liver disease
| Source of stem cells | No. of patients | Year | Reference | |
|---|---|---|---|---|
| 1 | BM-MNCs | 9 | 2006 | (59) |
| 2 | PB-MNCs, G-CSF | 2 | 2006 | (60) |
| 3 | BM-MSCs, G-CSF | Active Treatment: 8 (5 male) | 2006 | (61) |
| 4 | CD34+ cells | 5 | 2006 | (62) |
| 5 | CD34+ cells | 4 | 2007 | (63) |
| 6 | BM-MSCs | 10 | 2007 | (64) |
| 7 | PB-monocytes, G-CSF | 2 | 2007 | (65) |
| 8 | CD34+ cells | 4 | 2008 | (66) |
| 9 | CD34+ cells | 5 | 2008 | (67) |
| 10 | PB-MSCs, G-CSF | 40 | 2008 | (68) |
| 11 | CD34+ cells | 9 | 2008 | (69) |
| 12 | CD34+, CD133+ HSCs | Active Treatment: 90 (78 male) | 2010 | (70) |
| 13 | BM-MNCs | 10 | 2010 | (71) |
| 14 | BM-MNCs | 15 | 2010 | (72) |
| 15 | hHPCs | Active Treatment: 4 | 2010 | (73) |
| 16 | BM-MNCs, HSCs | 6 | 2011 | (74) |
| 17 | BM-MNCs | 5 | 2011 | (75) |
| 18 | BM-MNCs (BM-derived hepatocytes) | 20 | 2011 | (76) |
| 19 | CD34+ cells, G-CSF | 23 | 2012 | (77) |
| 20 | BM-MSCs, G-CSF | 28 | 2013 | (78) |
| 21 | BM CD133+ cells | 16 | 2015 | (79) |
| 22 | BM CD134+ cells, G-CSF | 81 | 2015 | (80) |
| 23 | PB CD34+ cells | 22 | 2015 | (81) |
| 24 | BM CD133+ cells, MNCs | 12 | 2016 | (82) |
| 25 | PB monocytes | 9 | 2017 | (83) |
|
|
||||
| Source of stem cells | Clinicaltrials.gov identifier | Year | Phase | |
|
|
||||
| 26 | Autologous expanded CD34+ HSCs | NCT00655707 | 2008∼2015 | Phase2 |
| 27 | Autologous BM cells | NCT02943707 | 2016∼2020 | Phase2 |
| 28 | Autologous BM-derived CD133 stem cells | NCT01120925 | 2010∼2014 | Phase2 |
| 29 | BM cells | NCT01412593 | 2011∼2013 | Phase2 |
| 30 | Autologous BM-derived CD133 stem cells | NCT00713934 | 2008∼2010 | Phase1 |
| 31 | Adult stem cells | NCT00147034 | 2005∼2016 | Phase1 |
| 32 | Autologous BM-derived CD133 stem cells | NCT01025622 | 2009∼2010 | Phase1 |
| 33 | Allogenic BMSCs | NCT01221454 | 2010 | Phase2 |
| 34 | Allogenic BMSCs | NCT01223664 | 2010 | Phase2 |
| 35 | Human BMSCs | NCT01724697 | 2012 | Phase1 |
| 36 | Autologous BMSCs | NCT02943707 | 2016 | Phase2 |
![]()
Go to :
Concluding Remarks
Several assumptions regarding HSCs underlie the translation of stem cell biology to regenerative medicine. As mentioned, HSC/progenitor cells isolated using CD markers can potentially replace hematopoietic cells. Among all adult derived stem cells, HSCs are the most powerful cell sources to treat diverse diseases and regenerative medicine. Since HSCs can be differentiated into progenitors including endothelial progenitors and mesenchymal stem/progenitor cells, it may be useful to construct strategies for effective therapy in pathologic issues. Especially, the treatment of liver diseases using stem cell therapy has been improved, and there has been evidence of a positive effect from stem cells. Although the mechanism by which stem cells ameliorate diseases such as liver and blood disorders remain unclear, positive results are continuously reported for their use in regenerative medicine. These results strongly suggest that further research using stem cells will be required to fully elucidate potential adverse effects, such as immune rejection, and understanding HSCs will be helpful for further expansion with feasibility of using stem cells in regenerative medicine.
Go to :
 XML Download
XML Download