

 PDF
PDF Citation
Citation Print
Print


Introduction
As a common metabolic disorder, diabetes mellitus (DM) is characterized by chronically raised blood glucose caused by insulin dysfunction (1). Methylglyoxal (MGO) plays a pivotal role in the occurrence and development of DM, which is a highly active dicarbonyl compound and a ubiquitous product of cell metabolism (2). Concretely, MGO is an inherent by-product of glycolysis with cell permeability, the aggregation of MGO is harmful, as it is the most active compound to induce glycosylation products in cells (3), and this will induce insulin resistance to further aggravate pathoglycemia and dyslipidemia, thus promoting the progress of the disease (4). In addition, MGO can easily react with proteins, fats and nucleic acids to form advanced glycation end products, leading to the occurrence of cataract, retinopathy, kidney disease, vascular disease and other diabetic complications (5-8). In recent years, although several MGO scavengers have been discovered, effective clinical strategy for treating insulin resistance induced by MGO is still not available (9). Therefore, finding new targets to reduce MGO-induced insulin resistance may have a positive significance in improving the vascular function of DM.
MicroRNAs (miRNAs) are a class of single-stranded non-coding RNAs with length of approximately 18∼23 nucleotides, and exert post-transcriptional modification through binding to the 3’untranslated region of target gene to affect the stability and translation of mRNA. It has shown that miRNAs account for about 1% of the genome of nuclear organisms, and their target genes regulate 30% of all genes (10). A recent study has found that miRNAs play an important role in the pathogenesis of DM by regulating the synthesis, secretion and signal transduction of insulin (11). Li (12) indicated that miR-375 regulates the expressions of insulin secretion-related genes to control the progression of DM. In addition, some scholars have found that miRNAs, such as miR-143, miR-145 and miR-29, also exert a vital function in insulin signaling pathway and insulin resistance. In related fields, miR-450a-5p can secrete various factors to regulate fat formation through paracrine signals (13). Correlatively, previous animal studies have shown that the formation of fat may be associated with the onset of DM (14). Nevertheless, the role and mechanism of miR-450a-5p in insulin resistance of endothelial cells remains unclear.
The relationship between miR-450a-5p and MGO was studied, and we further analyzed the effects and mechanisms of miR-450a-5p on MGO-induced human umbilical vein endothelial cells (HUVECs), including cell activity, migration, fat formation and insulin resistance. The aim of this study was to further analyze the action mechanism of miR-450a-5p in the insulin resistance induced by MGO in order to provide a potential target for related therapies, thus delaying the progression of DM and improving the prognosis of patients.
Go to : 
Materials and Methods
Cell culture and treatment
HUVECs were purchased by the American Type Culture Collection (ATCC, Rockville, MD, USA) and cultivated in EGM-2 Bullet Kit from Lonza (Walkersville, MD, USA). The cells were starved in medium containing with 0.25% albumin bovine serum (BSA) for 16 h and then respectively treated by low-glucose (5 mmol/L; Sigma, St Louis, USA), high-glucose (25 mmol/L; Sigma, St Louis, USA), and low-glucose combined with xylose (4.5 mmol/L; Sigma, St Louis, USA) for 24, 48, 72 h. In addition, different concentrations of MGO (0, 250, 500, 1000 μmol/L; Sigma, St Louis, USA) were added into HUVECs for 16 h, respectively. For comparison, cells were also pre-disposed with aminoguanidine bicarbonate salt (AG; 4 mmol/L; Sigma, St Louis, USA), and then treated by 500 μmol/L MGO for 16 h. Furthermore, insulin (100 nmol/L; Eli Lilly Florence, Italy) alone or in combined with MGO (500 μmol/L) were added into HUVECs for 10 min. For comparison, untreated cells (0 μmol/L) served as controls. Quantitative real-time polymerase chain reaction (qRT-PCR) assay was performed in treated HUVECs to determine the expression level of miR-450a-5p.
Cell transfection
Cell transfection was conducted in HUVECs in order to determine the effects of miR-450a-5p in HUVECs. Mimic control, miR-450a-5p mimic (#miR10001545-1-5, RiboBio, Guangzhou, China), inhibitor control, miR-450a-5p inhibitor (#miR20001545-1-5, RiboBio, Guangzhou, China), mimic plus negative control, and mimic combined with CREB (NM_009952.2, Han Bio, Shanghai, China) were separately transfected into MGO-treated cells. Moreover, miR-450a-5p mimic was also transfected into HUVECs after co-treatment with MGO and insulin. LipofectamineTM 2000 transfection reagent (Invitrogen, Carlsbad, California, USA) was used for cell transfection according to manufacturer’s instructions.
qRT-PCR assay
The expression levels of relevant mRNAs in HUVECs were measured by qRT-PCR assays. Concretely, total RNA was separated from HUVECs using TRIzol reagent (Invitrogen, Carlsbad, California, USA). NanoDrop-2000c spectrophotometer (Thermo Fisher Scientific, Massachusetts, USA) was used to determine the amount of RNA, and the integrity was determined by 1% agarose modified gel electrophoresis. Then, 1 μg of separated RNA was reverse transcribed into cDNA with the PrimeScript RT Master Mix Perfect Real Time (TaKaRa, Shiga, Japan) according to the manufacturer’s instructions. qRT-PCR assay was conducted using SYBR green qPCR assay (BioRad, USA) on the ABI Prism 7500 Fast Real-time PCR System (Applied Biosystems, Foster City, CA) following the reaction conditions: 30 cycles of hot start at 94℃ for 1 min, denaturation at 94℃ for 1 min, annealing at 50℃ for 45 s, and elongation at 72℃ for 2 min, then final elongation at 72℃ for 8 min. The sequences of primers were showed in Table 1 and synthesized by Gene Pharma (Shanghai, China). GAPDH or U6 served as internal reference, and the data were processed by the comparative 2−ΔΔCt method (15).
Cell viability
MTT assay was carried out to measure the viability of transfected HUVECs. Briefly, HUVECs were seeded in 96-well plates (1×104 cells/well). After co-treated with MGO and transfection, 20 μL MTT reagent (Sigma, USA) was added at 24, 48, and 72 h to determine the viability of cells, and the optical densities (OD) value was read at 570 nm using the ELX-800 Biotek plate reader (Winooski, USA).
Transwell assay
A 24-well Transwell chamber (Corning, MA, USA) was performed to detect the migration ability of HUVECs after the MGO treatment and transfection. In brief, the lower chamber was supplemented with media containing 10% fetal bovine serum (FBS; Gibco, USA). Resuspended HUVECs (5×104 cells) were moved into the upper chambers and cultured for 24 h. Then, migrated cells were fixed by methanol and stained with the 0.1% crystal violet solution for 15 min. Nikon Eclipse TS-100 inverted microscope (Tokyo, Japan) was used for observation.
Oil red O staining
Fat accumulation in HUVECs was detected by oil red O staining. Cells were washed by phosphate buffer saline (PBS) and fixed by 10% formaldehyde at room temperature for 10 min. After the MGO treatment and transfection, HUVECs were incubated with 0.5% Oil red O solution (Gibco, USA) for 15 min, and then washed by water for 5 min. Stained HUVECs were photographed and viewed under DP73 Olympus microscope (Tokyo, Japan).
Griess reaction
NO in HUVECs culture medium was measured by Griess reaction. After the treatment and transfection, cell culture medium was collected, and the supernatant fraction after centrifugation (1000 g, 15 min) was taken as a sample solution for the measurement of NO using Nitrate/nitric Assay Kit Colorimetric (Sigma, USA). The assay used contained a chromophoric azo-derivative that allows NO to be detected at 540∼570 nm by the microplate reader (Spectra MAX190; MD, USA).
Double-luciferase reporter analysis
Targetscan 7.2 (http://http://www.targetscan.org/.) predicted that CREB was the potential target for miR-450a-5p. For determination, double-luciferase report system detection kit (Promega, Madison, Wisconsin, USA) was used for double-luciferase reporter analysis. Briefly, HUVECs were seeded in 24-well plates (1×105 cell/well) for 24 h. Then, miR-450a-5p mimic or blank control was transfected into HUVECs combined with wild type CREB (CREB-WT) or its mutant type (CREB-MUT) using LipofectamineTM 2000. After 48 h, the cells were lysed and the luciferase activity was evaluated with the GloMax® Discover Multimode Microplate Reader (GM3000, Promega, USA).
Western blot (WB) analysis
The expressions of related proteins in HUVECs were measures by WB assays. Total proteins of cells were separated by the RIPA buffer (Solarbio, Beijing, China), and Bicinchoninic Protein Assay kit (BCA, Pierce, Rockford, IL, USA) was used for quantitation. 30 μg of total protein was isolated using the sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE, Beyotime, Shanghai, China), and moved onto the polyvinylidene fluoride (PVDF) membranes. Then, 5% non-fat dried milk was used to seal the membranes for 2 h. Primary antibodies including p-eNOS (1:2000, ab184154, Abcam, USA), p-eNOS (1:1000, ab138430, Abcam, USA), eNOS (1:1000, ab76198, Abcam, USA),p-Akt (1:1000, #9271, Cell Signalling Technology, USA) and Akt (1:1000, #9272, Cell Signalling Technology, USA) were subsequently used to incubated the membranes overnight at 4℃. GAPDH (1:1000, ab8245, Abcam, USA) served as the internal control. Subsequently, the homologous secondary antibodies goat anti-rabbit IgG H&L (HRP; 1:7000, ab97051, Abcam, USA) and goat anti-mouse IgG H&L (HRP; 1:1000, ab150113, Abcam, USA) were added for another 1 h at room temperature. The bands were developed by an enhanced chemiluminescence-detecting kit (Thermo Fisher, MA, USA).
Statistical analysis
Statistical Package of the Social Sciences 20.0 software (SPSS, Inc., Chicago, USA) was used for data analysis. The data were shown as mean±standard deviation (SD). The comparison between groups was performed by Student’s t-test or one-way analysis of variance (ANOVA) followed by Turkey’s post-hoc test. All experiments were repeated in triplicate. p<0.05 was considered as statistically significant.
Go to :
Results
Over-expressed miR-450a-5p inhibited viability, migration, and lipid formation of MGO-induced HUVECs
Based on qRT-PCR analysis, the expression level of miR-450a-5p in HUVECs was observably down-regulated after 72 h of high-glucose treatment (p<0.001), while low-glucose had no obvious effect on miR-450a-5p expression (Fig. 1A). Moreover, the expression of miR-450a-5p was obviously inhibited by adding MGO in HUVECs but reversed by AG (p<0.001; Fig. 1B and 1C). In order to further investigate the effects of miR-450a-5p and MGO on the cells, transfection was performed in the MGO-induced cells. The results revealed that miR-450a-5p was obviously up-regulated by mimic but was down-regulated by inhibitor (p<0.001; Fig. 2A), showing that the transfection was successful. Interestingly, MTT results showed that over-expressed miR-450a-5p prominently suppressed cell viability of MGO-induced HUVECs after transfection for 48 and 72 h (p<0.001; Fig. 2B), while down-regulated miR-450a-5p had no significant effect on cell viability. In transwell assay, microscopic and quantitative analyses showed that over-expressed miR-450a-5p observably suppressed the migration of MGO-induced HUVECs (p<0.001; Fig. 2C and 2D), whereas down-regulated miR-450a-5p had no significant effect on the migration. In the oil red O staining, lipid formation showed an orange color. From experimental images, we observed that over-expressed miR-450a-5p significantly reduced the lipid accumulation of MGO-induced HUVECs (Fig. 2D), while the lipid accumulation of cells was still high after down-regulating miR-450a-5p.
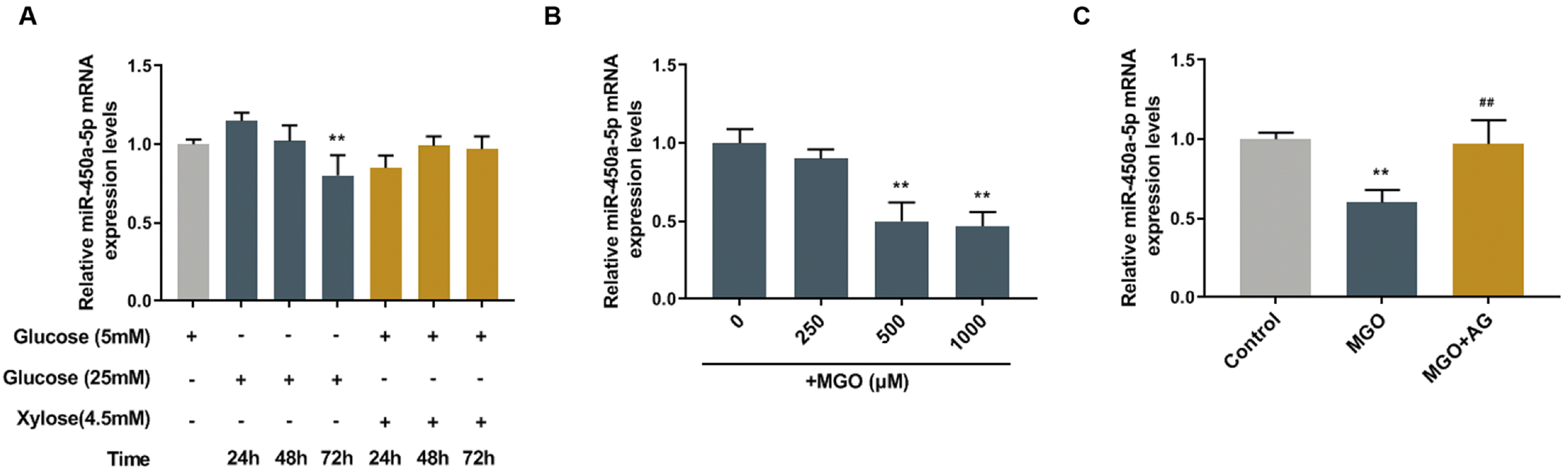 | Fig. 1Methylglyoxal (MGO) down-regulated the expression of miR-450a-5p in human umbilical vein endothelial cells (HUVECs). (A) Quantitative real-time polymerase chain reaction (qRT-PCR) assay measured the expression of miR-450a-5p in HUVECs after treatment with low-glucose (5 mM), high-glucose (25 mM), or low-glucose combined with xylose (4.5 mM) for 24, 48, 72 h. (B) qRT-PCR assay determined the expression of miR-450a-5p in HUVECs treated by different concentrations of MGO (0, 250, 500, 1000 μM). (C) qRT-PCR assay determined the expression of miR-450a-5p in HUVECs treated by control, MGO only or combined with aminoguanidine bicarbonate salt (AG). **p<0.001, vs. low-glucose, 0 μm, or Control; ##p<0.001, vs. MGO. n=3.
|
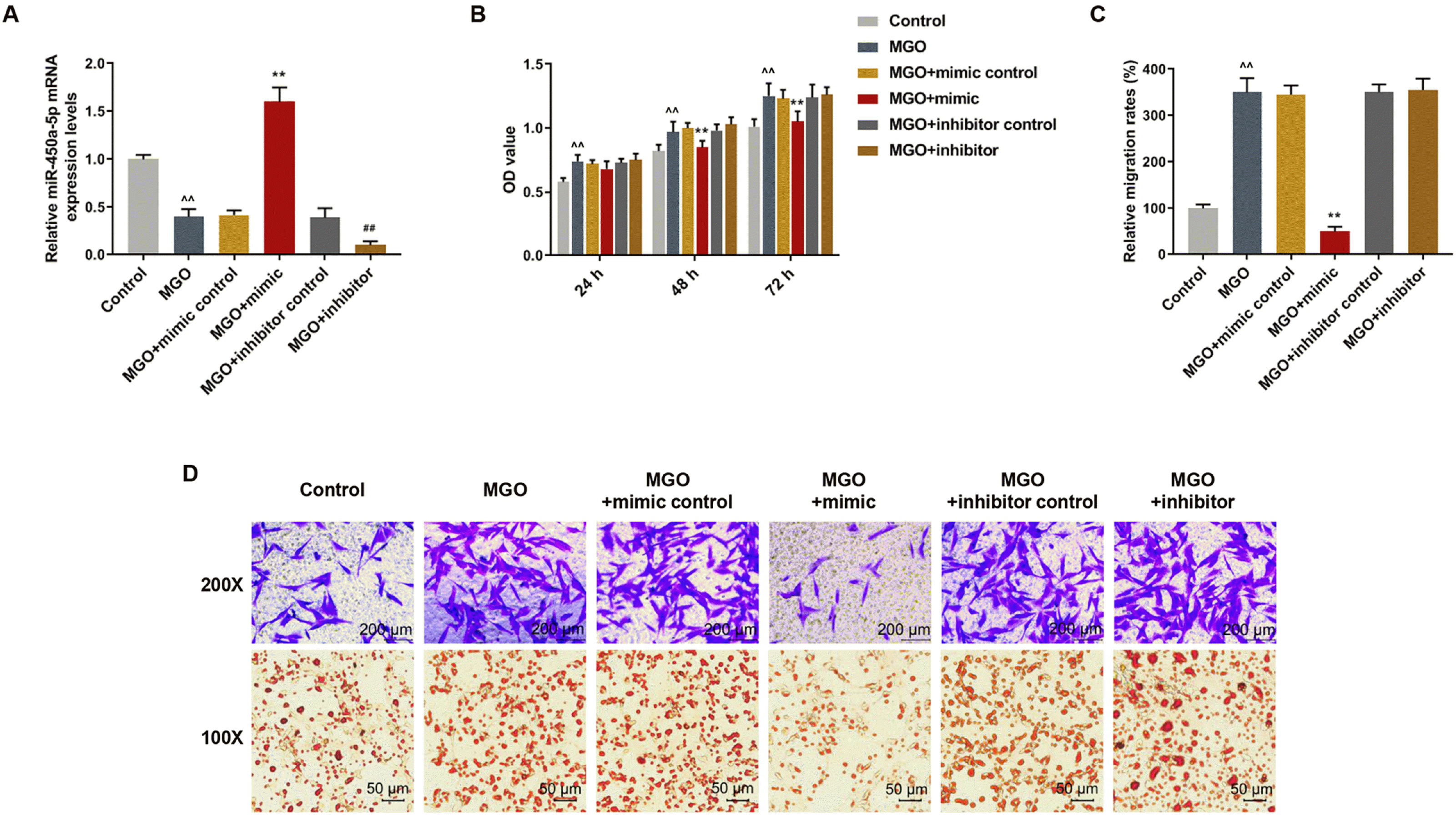 | Fig. 2Over-expressed miR-450a-5p inhibited viability, migration, and lipid formation of methylglyoxal (MGO)-induced human umbilical vein endothelial cells (HUVECs). Mimic control, miR-450a-5p mimic, inhibitor control, and miR-450a-5p inhibitor were respectively transfected into MGO-treated HUVECs. (A) Quantitative real-time polymerase chain reaction (qRT-PCR) assay measured the expression of miR-450a-5p in MGO-treated HUVECs after the transfection. (B) MTT assay detected the optical densities (OD) values of MGO-induced HUVECs after the transfection for 24, 48 and 72 h. (C) Quantitative analysis of Transwell assay in MGO-treated HUVECs after the transfection. (D) Microscopic images of Transwell and Oil red O staining experiments in MGO-induced HUVECs after transfection. ^^p<0.001, vs. Control; **p<0.001, vs. MGO+mimic control; ##p<0.001, vs. MGO+inhibitor control. n=3.
|
Over-expressed miR-450a-5p reduced the insulin resistance of MGO-induced HUVECs by targeting CREB
WB analysis suggested that the phosphorylation of eNOS and AKT was promoted by insulin but was reversed by MGO, and adding miR-450a-5p decreased the effects of MGO on eNOS/AKT pathway (p<0.001; Fig. 3A∼C). Analogously, Griess reaction displayed that MGO inhibited the insulin-induced NO release but the effect as such could be relieved by up-regulated miR-450a-5p (p<0.001; Fig. 3D).
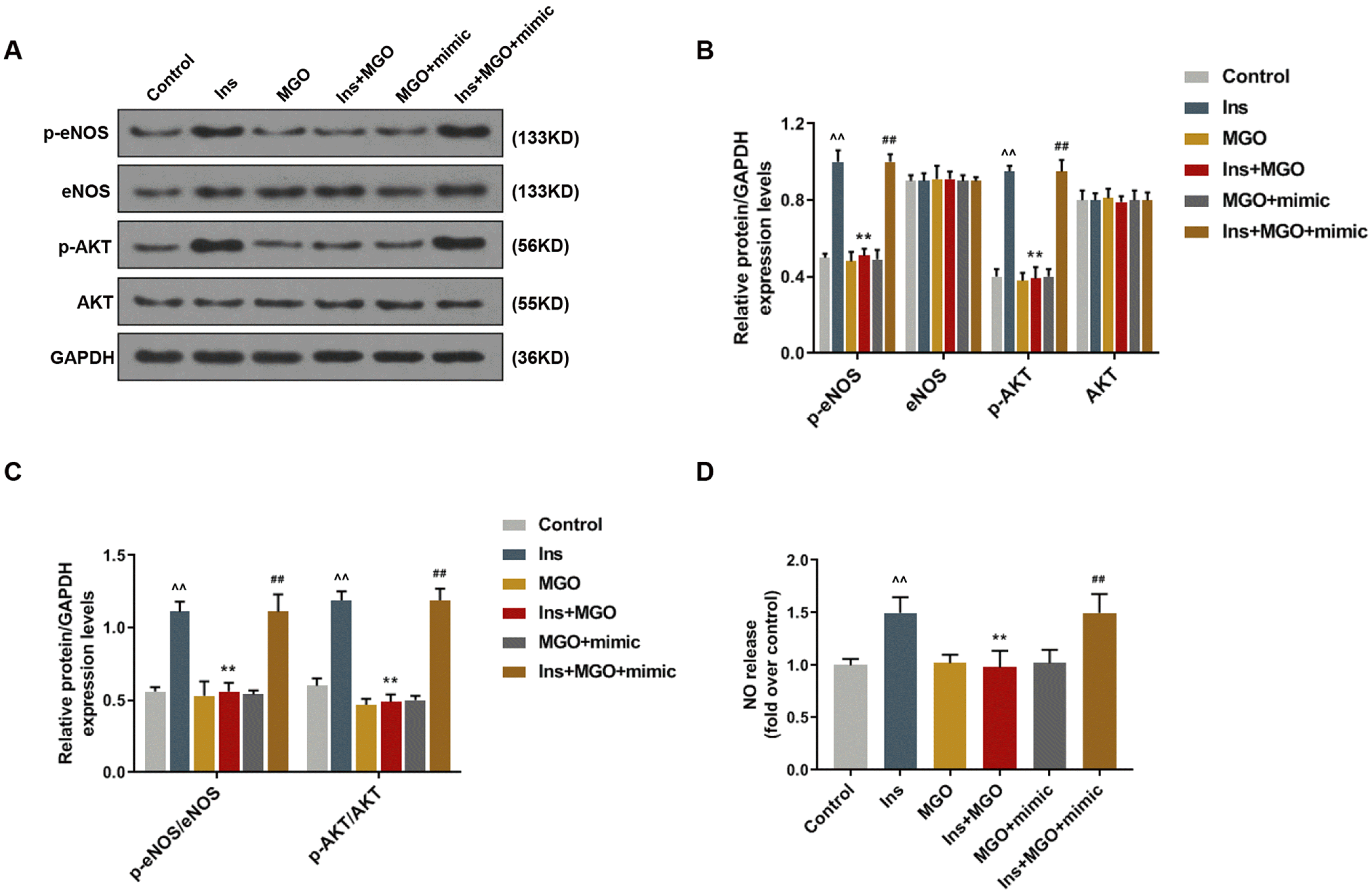 | Fig. 3Over-expressed miR-450a-5p inhibited the insulin resistance of methylglyoxal (MGO)-induced human umbilical vein endothelial cells (HUVECs). In this figure, HUVECs were respectively treated by control, insulin (Ins), MGO, insulin combined with MGO, miR-450a-5p mimic plus MGO, or mimic plus MGO and insulin. (A∼C) Western blot (WB) bands and quantitative analysis determined the expressions of eNOS/AKT pathway-related proteins in HUVECs. (D) Griess reaction assessed the release of NO in HUVECs. ^^p<0.001, vs. Control; **p<0.001, vs. Ins; ##p<0.001, vs. Ins+MGO. n=3.
|
Targetscan7.2 predicted that the position 108-114 of CREB1 3’UTR was the binding site of miR-450a-5p (Fig. 4A). Double-luciferase reporter analysis indicated that the luciferase activity of HUVECs was decreased observably after the co-transfection with miR-450a-5p and CREB-WT, suggesting that CREB was the target gene for miR-450a-5p (p<0.001; Fig. 4B). Moreover, MGO promoted the mRNA expression of CREB in HUVECs, whereas miR-450a-5p had the completely opposite effect (p<0.001; Fig. 4C). As for the effects of CREB on eNOS/AKT pathway, WB experiment revealed that the up-regulation of CREB reduced the phosphorylation of eNOS/AKT pathway in HUVECs induced by insulin (p<0.001; Fig. 5).
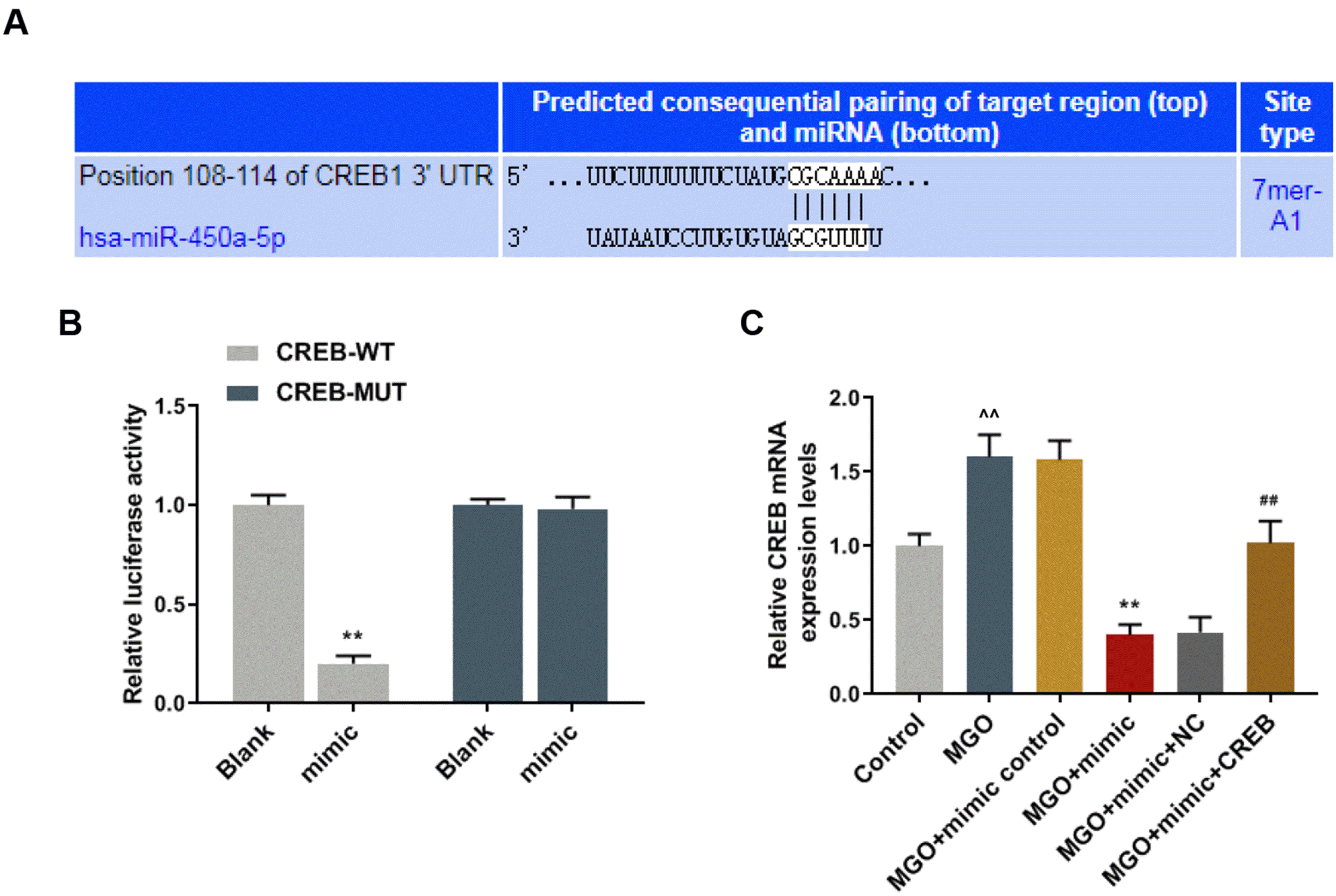 | Fig. 4CREB was the target gene for miR-450a-5p. (A) Targetscan7.2 predicted that the position 108-114 of CREB1 3’UTR was the binding site of miR-450a-5p. (B) Double-luciferase reporter analysis measured the luciferase activity of human umbilical vein endothelial cells (HUVECs) after transfection with miR-450a-5p mimic plus wild type CREB (CREB-WT) or its mutant type (CREB-MUT). (C) Quantitative real-time polymerase chain reaction (qRT-PCR) assay measured the expression of CREB in methylglyoxal (MGO)-induced HUVECs after transfection with mimic control, miR-450a-5p mimic, mimic plus negative control (NC) or CREB. ^^p<0.001, vs. Blank, or Control; **p<0.001, vs. MGO+mimic control; ##p<0.001, vs. MGO+mimic+NC. n=3.
|
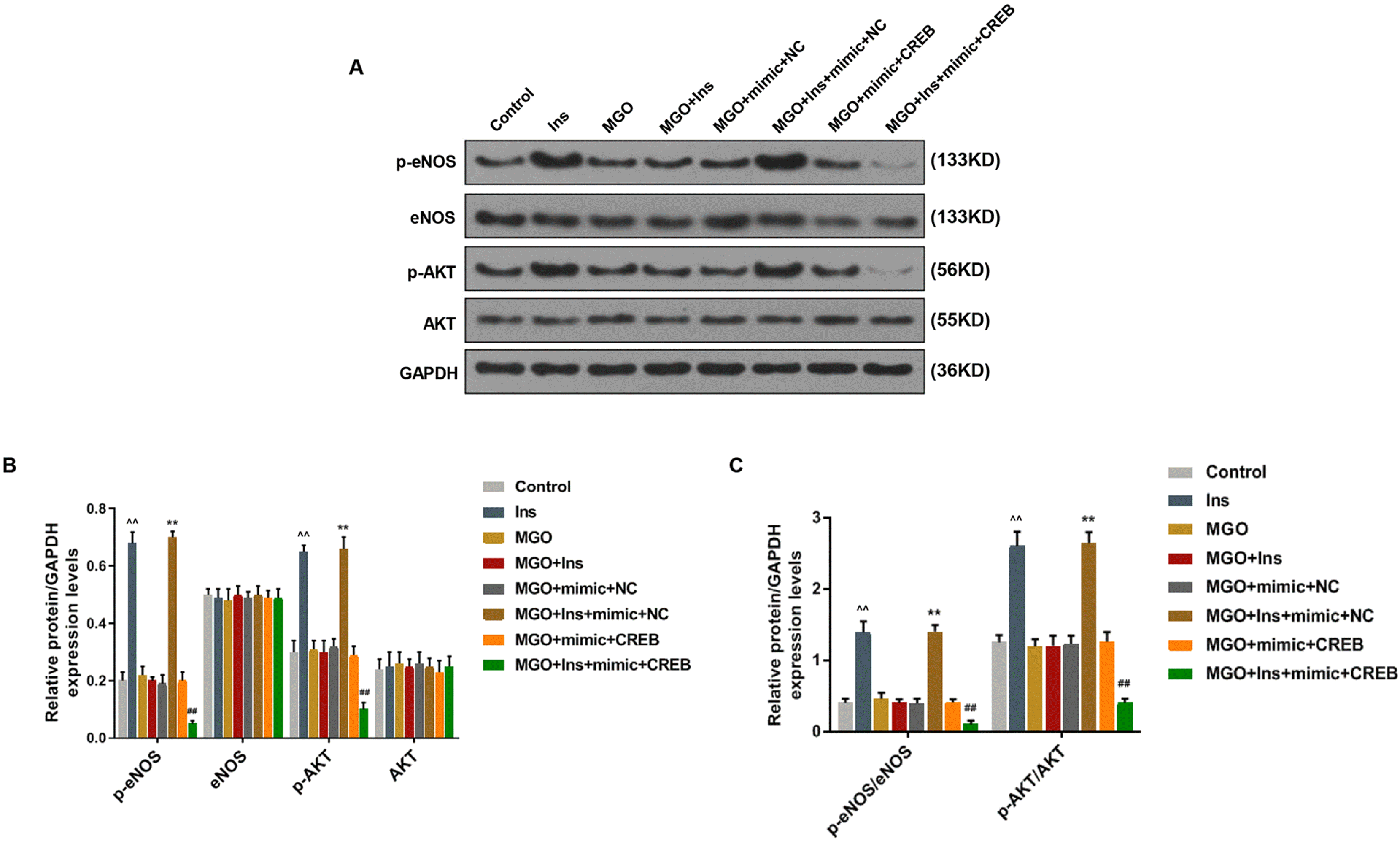 | Fig. 5Over-expressed miR-450a-5p reduced the insulin resistance of methylglyoxal (MGO)-induced human umbilical vein endothelial cells (HUVECs) by targeting CREB. Western blot (WB) bands and quantitative analysis determined the expressions of eNOS/AKT pathway-related proteins in MGO-induced HUVECs after treatment with control, insulin (Ins), MGO, MGO+Ins, MGO+miR-450a-5p mimic+negative control (NC), MGO+Ins+mimic+NC, MGO+mimic+CREB, or MGO+Ins+mimic+CREB. ^^p<0.001, vs. Control; **p<0.001, vs. MGO+mimic+NC; ##p<0.001, vs. MGO+mimic+CREB. n=3.
|
Go to :
Discussion
MGO can be obtained from food or produced in the body. In vivo, the main pathway of MGO production is glycolysis, and its content is abnormally increased by DM or hyperglycemia (16). MGO can be self-cleared by the body under normal physiological conditions, as MGO is characterized by hyper-responsiveness and short half-life, and the human body contains a variety of enzymes degrading MGO (17). In abnormal conditions, the dysfunction of MGO clearing pathway will lead to the gradual accumulation of MGO content, which can not only directly cause damage to cells, but also induce the massive generation of advanced glycation end products in the body (3). In previous studies, miRNAs such as miR-30b (18), miR-214 (19), miR-9a-3p (20) have been proved to be closely related to the content of MGO. In this study, the expression of miR-450a-5p, which showed a down-regulated trendency in HUVECs in high-glucose status, was also decreased with the increase of MGO concentration. Therefore, it can be speculated that miR-450a-5p might participate in the glycolysis process and play a role in the pathogenesis of DM.
From in vitro cell experiments, this paper discovered that MGO could not only enhance the activity of HUVECs, but also promote cell invasion and fat formation, which was consistent with a previous report (21). Interestingly, the over-expression of miR-450a-5p could reverse the actions of MGO in HUVECs, reduce cell activity, and inhibit cell migration and fat accumulation, suggesting that miR-450a-5p could improve MGO-induced cell damage. It has also been verified that the over-expression of miR-450 observably reduced the proliferation and invasion of lung cancer cells and controlled the growth of tumors (22). Moreover, miR-450a-5p also regulated cell apoptosis by blocking cell cycle and up-regulated miR-450a-5p has an inhibitory effect in the invasion of ovarian cancer cells (23). These findings further suggest that up-regulated miR-450a-5p could alleviate cell deterioration and could be used in treatment of related diseases.
As for the therapy of DM, insulin is the most frequent used drug in the treatment of the disease. Unsatisfactorily, the continuous accumulation of MGO can cause the occurrence and development of insulin resistance in the body (24). In related pathways, insulin has the function of protecting endothelial cells via modulating the activation of eNOS/AKT pathway (25). Concretely, eNOS is an endothelial nitric oxide synthase and can directly affect the ability of endothelial cells to synthesize NO (26). The phosphorylation of AKT activates eNOS, thereby promoting the production of NO and maintaining the normal function of endothelial cells (27). Insulin has the effect of activating eNOS/AKT pathway and promoting the release of NO, thus preventing endothelial cells from damage, which is the same as the results in the current study. However, we also found that MGO inhibited insulin-mediated eNOS/AKT pathway activation, whereas up-regulated miR-450a-5p eliminated the resistance of MGO to insulin, indicating that miR-450a-5p could reverse the effect on insulin resistance mediated by MGO.
Prediction tools and double-luciferase reporter analysis found that CREB had a binding site with miR-450a-5p and CREB was the target gene for miR-450a-5p. Furthermore, miR-450a-5p reversed the effect of MGO on promoting CREB. In the past, research mainly focused on the function of CREB in the nervous system, and it is known that CREB is widely involved in the biological functions including learning and memory regulation (28). In recent years, researchers demonstrated that activated CREB can promote the proliferation and migration of endothelial cells (29). In addition, activated CREB in the liver promotes hepatic insulin resistance and breaks glucose balance by promoting the CREB Coactivator 2 (CRTC2) activity (30). In this study, we found that CREB obviously suppressed the phosphorylation of eNOS and AKT, suggesting that CREB had an activation effect on the eNOS/AKT pathway. In related studies, Niwano et al. (31) also reported that CREB competitively bound to the cAMP/ATF reactive element to regulate eNOS gene expression in endothelial cells. These finding indicate that miR-450a-5p might relieve MGO-induced insulin resistance via targeting CREB.
In conclusion, the expression of miR-450a-5p is decreased obviously in endothelial cells under the conditions of high-glucose and 500, 1000 μmol/L MGO. In vitro cell experiments show that MGO could not only increase the activity of endothelial cells, but also accelerate cell migration and fat accumulation, which however, could be reversed by up-regulated miR-450a-5p. Moreover, MGO inhibits eNOS/AKT pathway activation and NO release mediated by insulin. Nevertheless, up-regulated miR-450a-5p eliminates the resistance of MGO to insulin via targeting CREB, thus, miR-450a-5p might be a potential therapeutic target to improve MGO-induced insulin resistance.
Go to :
 XML Download
XML Download